
Prolonged Sleep Deprivation Induces a Reprogramming of Circadian Rhythmicity with the Hepatic Metabolic Transcriptomic Profile

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Sleep Deprivation
2.3. Blood Tests
2.4. Intraperitoneal Glucose Tolerance Test (IPGTT)
2.5. Locomotor Rhythm Recording
2.6. Total RNA Extraction
2.7. Real-Time PCR
2.8. RNA-Seq
2.9. Statistical Analysis
2.10. RNA-Seq Data Analysis
3. Results
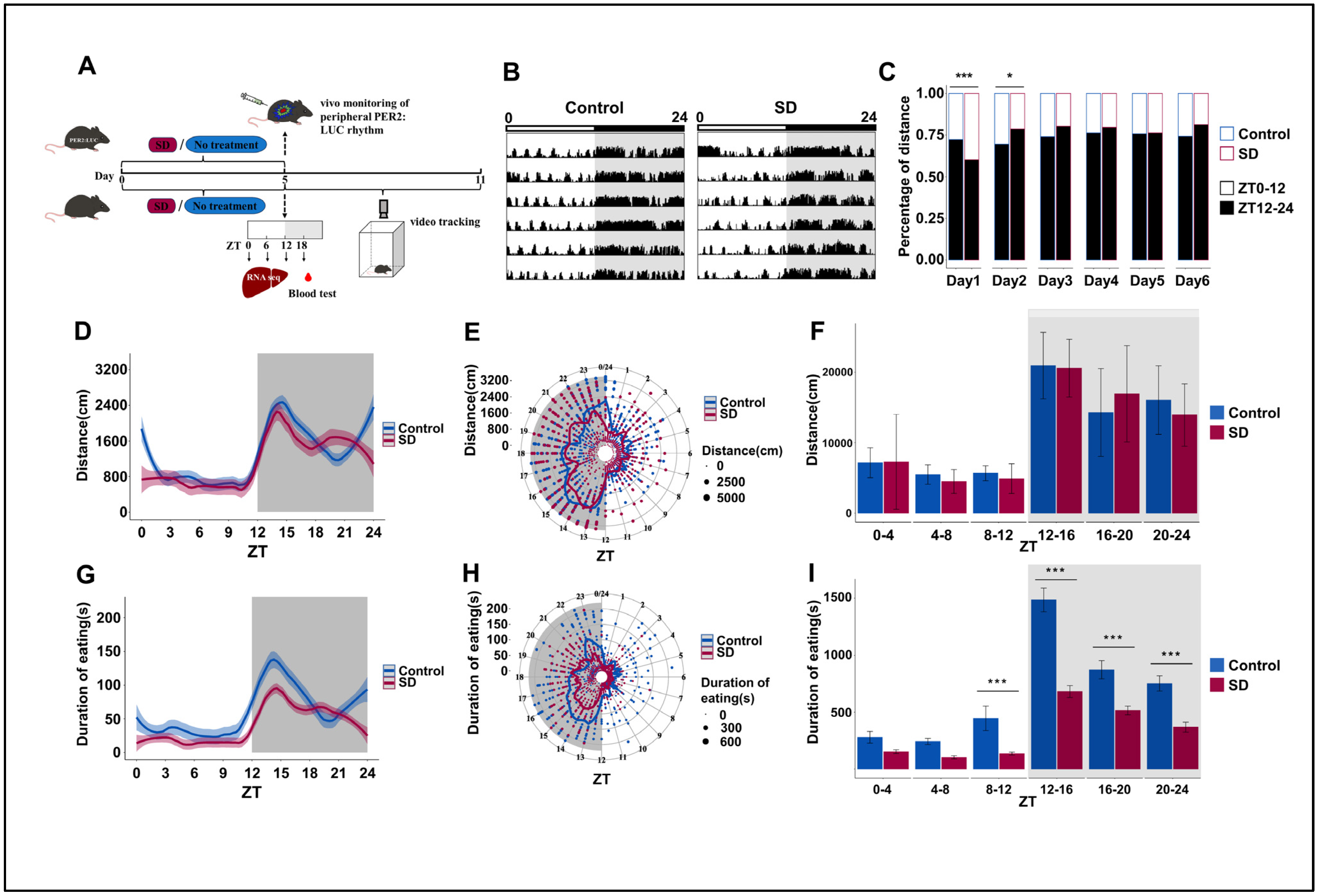
3.1. Prolonged Sleep Deprivation Alters the Locomotor-Rest and Feeding Rhythms in Mice
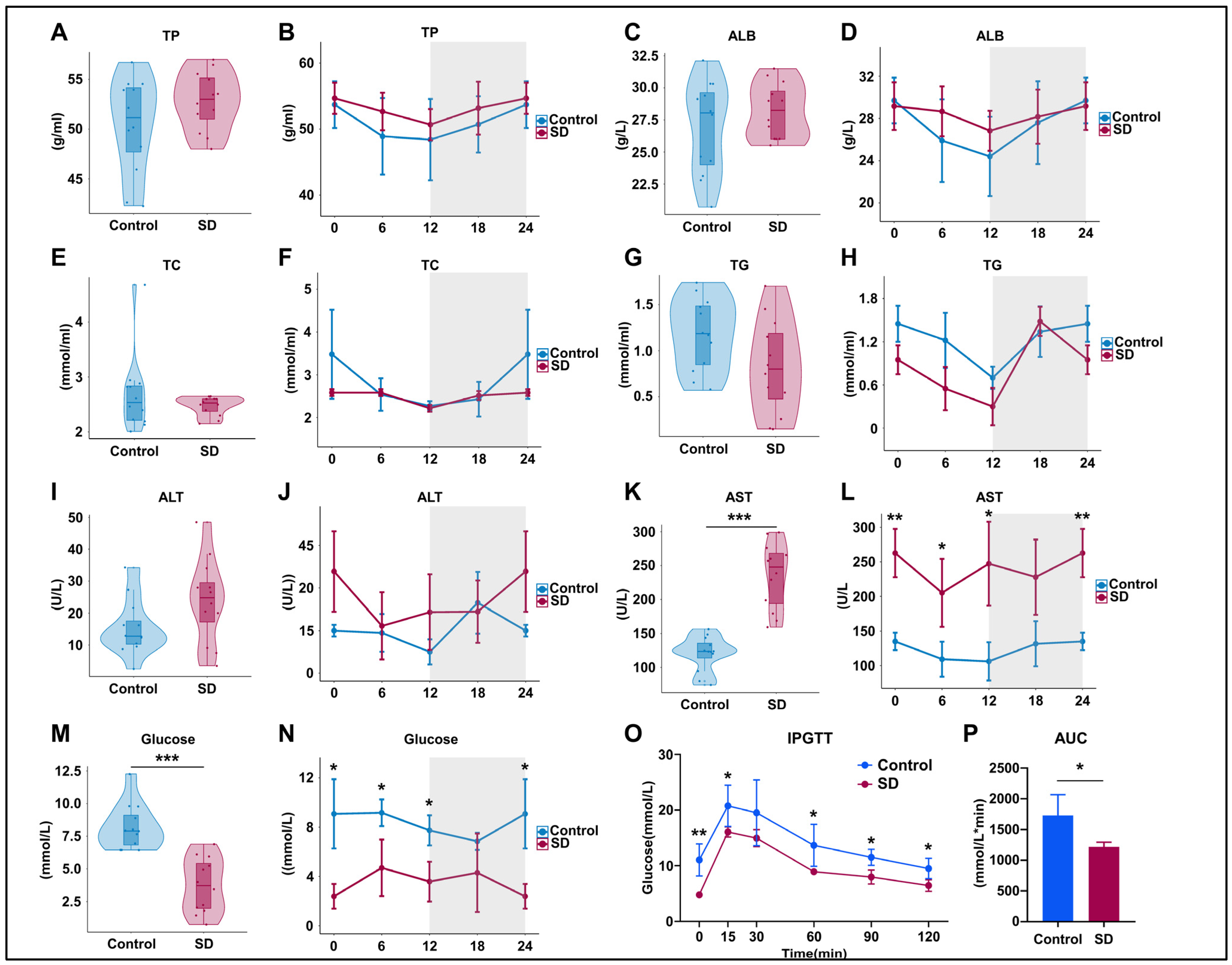
3.2. Prolonged Sleep Deprivation Alters Metabolism and Liver Function in Mice
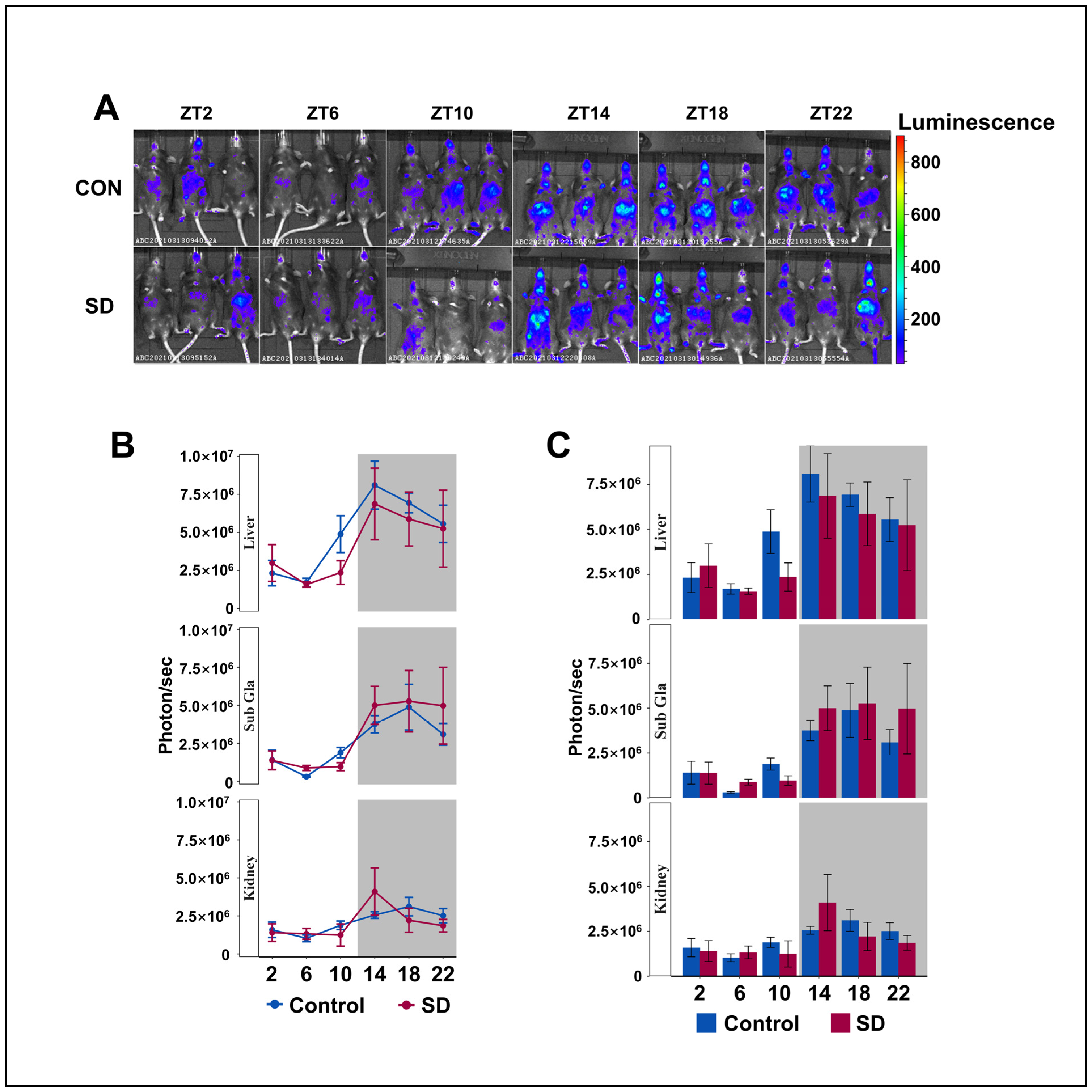
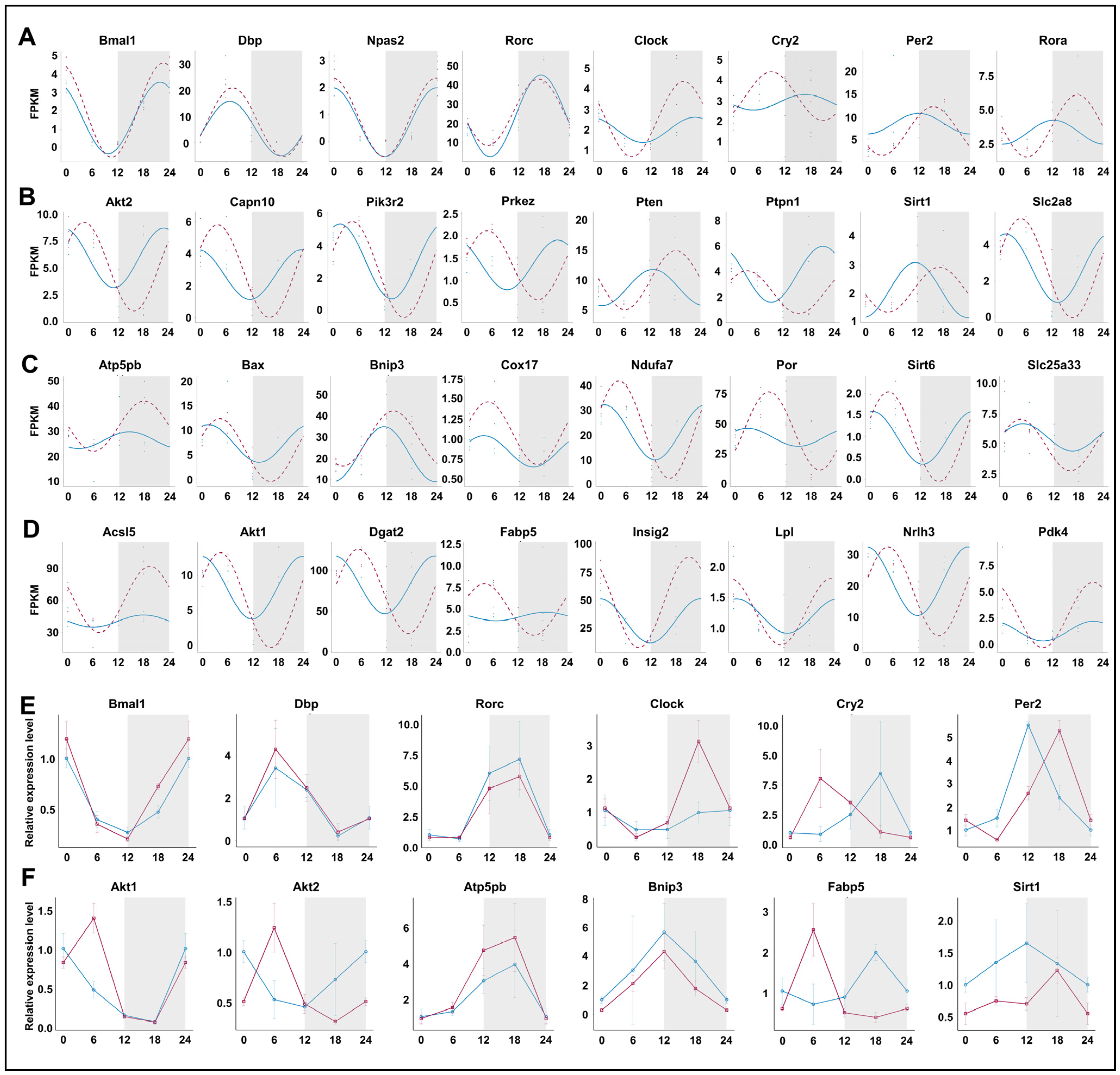
3.3. Peripheral Per2 Rhythms Exhibited Subtle Changes Following Prolonged Sleep Deprivation
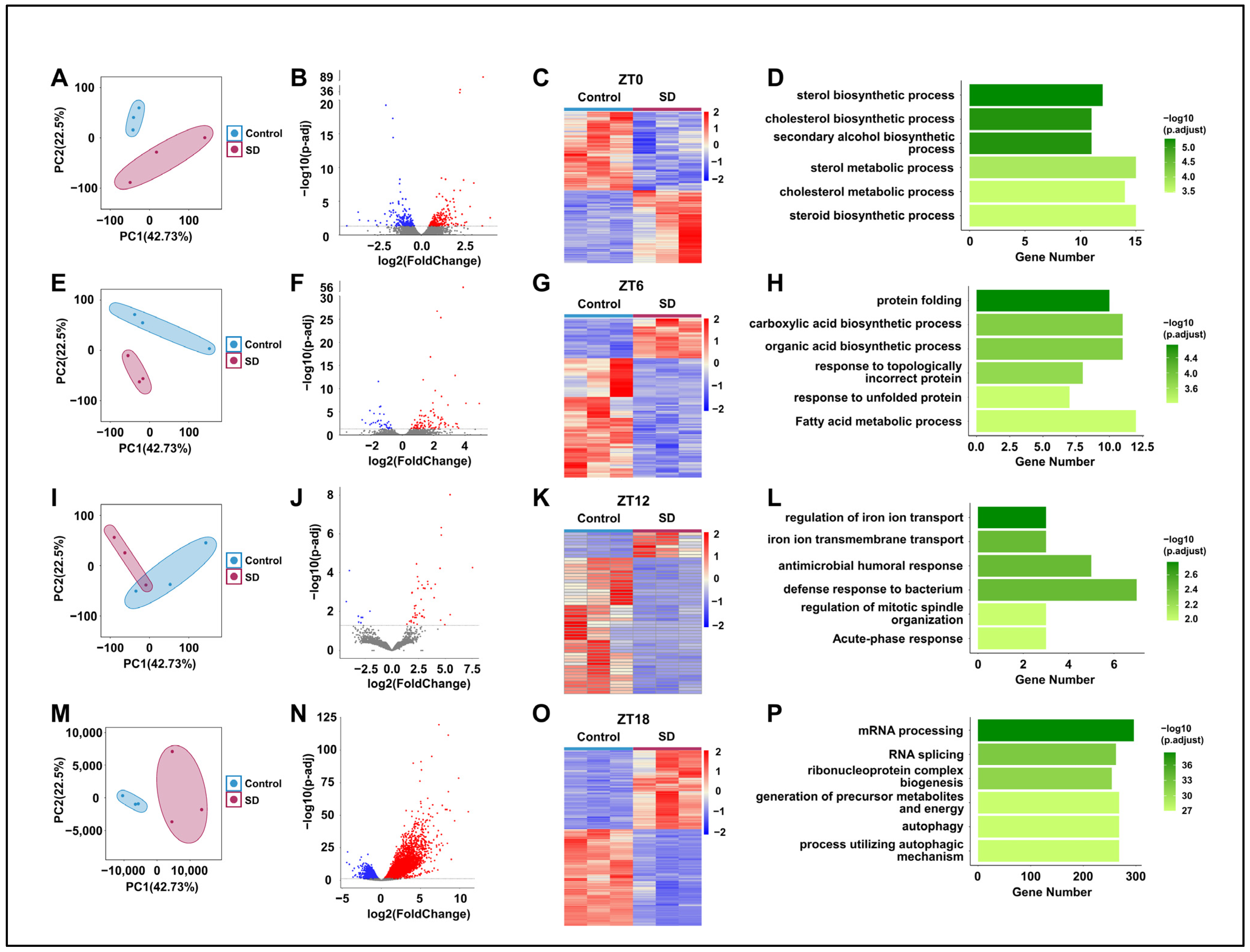
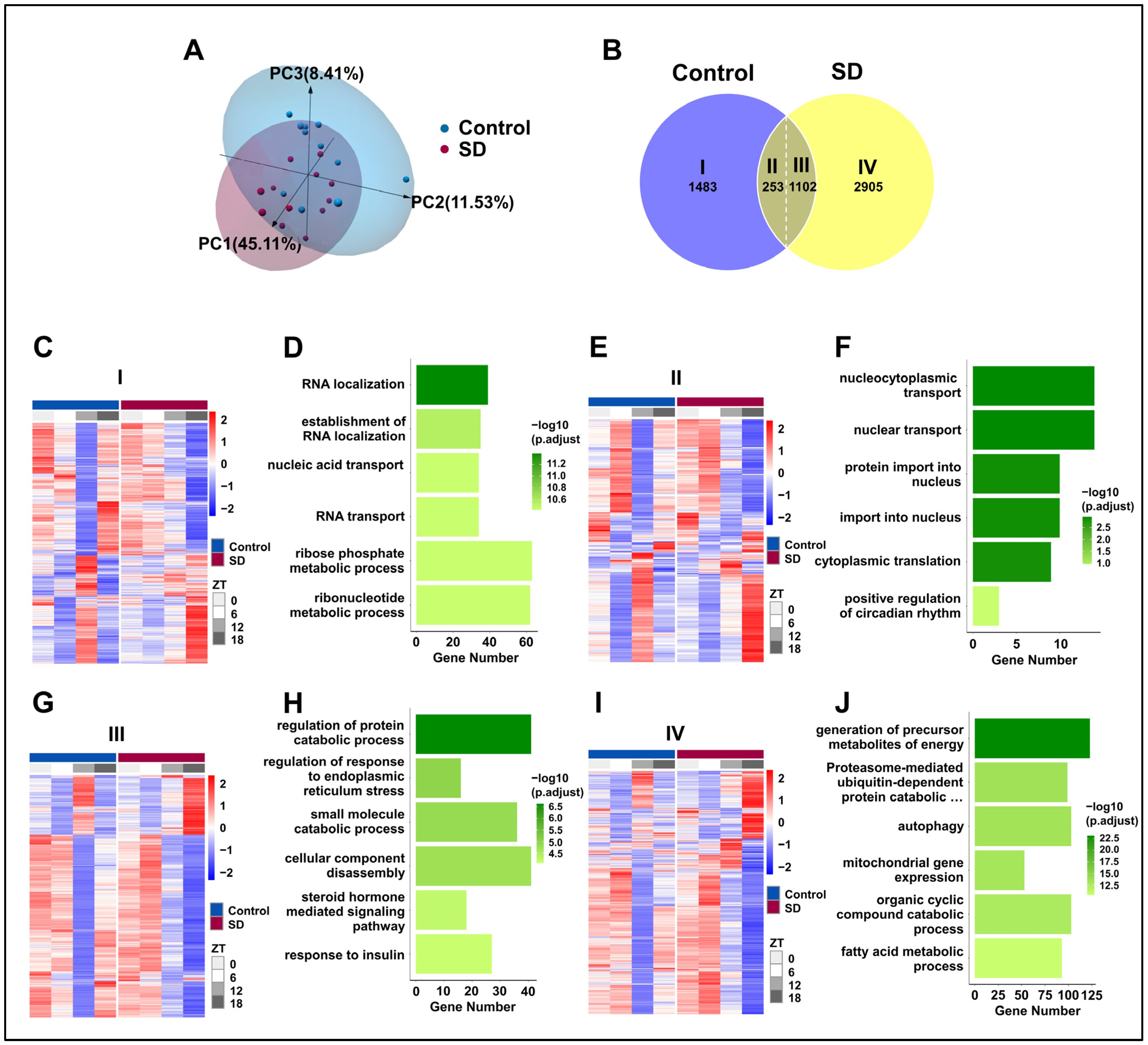
3.4. Prolonged Sleep Deprivation Alters the Liver Transcriptome Profiles of Mice across Different Time Points
3.5. Prolonged Sleep Deprivation Induces an Increase in Rhythmic Gene Expression
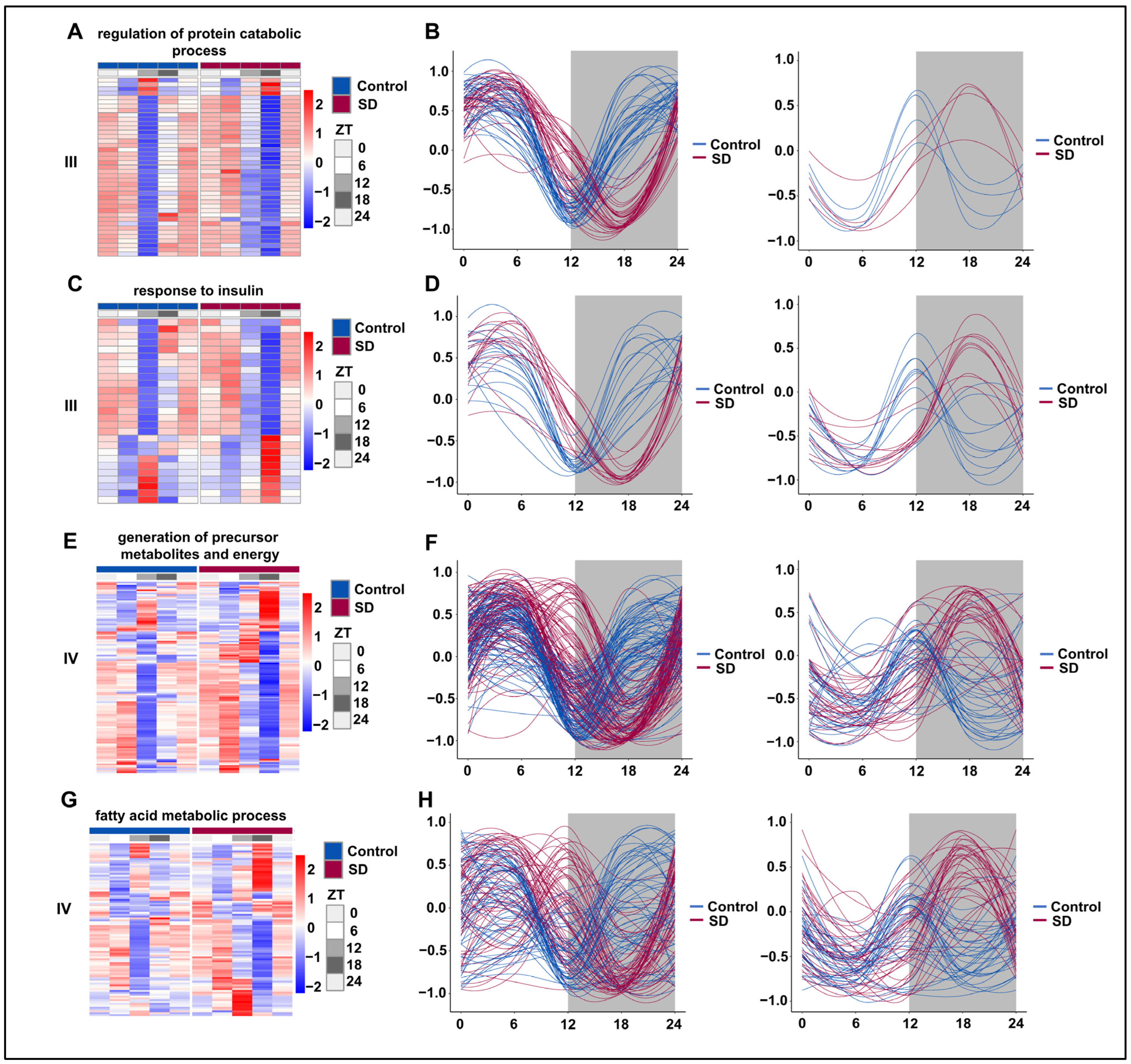
3.6. Prolonged Sleep Deprivation Resets the Rhythmicity of Multiple Metabolic Pathways
3.7. Prolonged Sleep Deprivation Synchronizes the Expression of a Large Number of Metabolic Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bass, J.; Lazar, M.A. Circadian time signatures of fitness and disease. Science 2016, 354, 994–999. [Google Scholar] [CrossRef]
- Adlanmerini, M.; Carpenter, B.J.; Remsberg, J.R.; Aubert, Y.; Peed, L.C.; Richter, H.J.; Lazar, M.A. Circadian lipid synthesis in brown fat maintains murine body temperature during chronic cold. Proc. Natl. Acad. Sci. USA 2019, 116, 18691–18699. [Google Scholar] [CrossRef]
- Panda, S.; Hogenesch, J.B.; Kay, S.A. Circadian rhythms from flies to human. Nature 2002, 417, 329–335. [Google Scholar] [CrossRef]
- Takahashi, J.S.; Shimomura, K.; Kumar, V. Searching for genes underlying behavior: Lessons from circadian rhythms. Science 2008, 322, 909–912. [Google Scholar] [CrossRef]
- Yurgel, M.E.; Masek, P.; DiAngelo, J.; Keene, A.C. Genetic dissection of sleep-metabolism interactions in the fruit fly. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2015, 201, 869–877. [Google Scholar] [CrossRef]
- Carneiro-Barrera, A.; Amaro-Gahete, F.J.; Jurado-Fasoli, L.; Sáez-Roca, G.; Martín-Carrasco, C.; Tinahones, F.J.; Ruiz, J.R. Effect of a Weight Loss and Lifestyle Intervention on Dietary Behavior in Men with Obstructive Sleep Apnea: The INTERAPNEA Trial. Nutrients 2022, 14, 2731. [Google Scholar] [CrossRef]
- Jia, J.; Tao, W.; Chen, T.; Zhong, Q.; Sun, J.; Xu, Y.; Sui, X.; Chen, C.; Zhang, Z. SIRT6 Improves Hippocampal Neurogenesis Following Prolonged Sleep Deprivation through Modulating Energy Metabolism in Developing rats. Mol. Neurobiol. 2023, 61, 883–899. [Google Scholar] [CrossRef]
- Magalhães, P.; Pereira, B.; Oliveira, A.; Santos, D.; Núñez, J.C.; Rosário, P. The Mediator Role of Routines on the Relationship between General Procrastination, Academic Procrastination and Perceived Importance of Sleep and Bedtime Procrastination. Int. J. Environ. Res. Public Health 2021, 18, 7796. [Google Scholar] [CrossRef]
- Hall, W.L. The emerging importance of tackling sleep-diet interactions in lifestyle interventions for weight management. Br. J. Nutr. 2022, 128, 561–568. [Google Scholar] [CrossRef]
- Huang, W.; Ramsey, K.M.; Marcheva, B.; Bass, J. Circadian rhythms, sleep, and metabolism. J. Clin. Investig. 2011, 121, 2133–2141. [Google Scholar] [CrossRef]
- Wright, K.P., Jr.; Drake, A.L.; Frey, D.J.; Fleshner, M.; Desouza, C.A.; Gronfier, C.; Czeisler, C.A. Influence of sleep deprivation and circadian misalignment on cortisol, inflammatory markers, and cytokine balance. Brain Behav. Immun. 2015, 47, 24–34. [Google Scholar] [CrossRef]
- Ramsey, K.M.; Bass, J. Circadian clocks in fuel harvesting and energy homeostasis. Cold Spring Harb. Symp. Quant. Biol. 2011, 76, 63–72. [Google Scholar] [CrossRef]
- Perogamvros, L.; Pépin, J.L.; Thorens, G.; Mégevand, P.; Claudel, E.; Espa, F.; Besson, M.; Cervena, K.; Janssens, J.P.; Lador, F. Baclofen-Associated Onset of Central Sleep Apnea in Alcohol Use Disorder: A Case Report. Respiration 2015, 90, 507–511. [Google Scholar] [CrossRef]
- Sengupta, A.; Rhoades, S.D.; Kim, E.J.; Nayak, S.; Grant, G.R.; Meerlo, P.; Weljie, A.M. Sleep restriction induced energy, methylation and lipogenesis metabolic switches in rat liver. Int. J. Biochem. Cell Biol. 2017, 93, 129–135. [Google Scholar] [CrossRef]
- Leproult, R.; Van Cauter, E. Role of sleep and sleep loss in hormonal release and metabolism. Endocr. Dev. 2010, 17, 11–21. [Google Scholar] [CrossRef]
- Kim, T.W.; Jeong, J.H.; Hong, S.C. The impact of sleep and circadian disturbance on hormones and metabolism. Int. J. Endocrinol. 2015, 2015, 591729. [Google Scholar] [CrossRef]
- Pejovic, S.; Vgontzas, A.N.; Basta, M.; Tsaoussoglou, M.; Zoumakis, E.; Vgontzas, A.; Bixler, E.O.; Chrousos, G.P. Leptin and hunger levels in young healthy adults after one night of sleep loss. J. Sleep Res. 2010, 19, 552–558. [Google Scholar] [CrossRef]
- Melinda, K.; Zsolt, G.; László-István, B.; Zoltán, K.I. The Antidepressants and the Metabolic Syndrome. Bull. Med. Sci. 2018, 91, 89–98. [Google Scholar] [CrossRef]
- Sumer, M.M. Stroke and Sleep Disorders. Examines Phys. Med. Rehabil. 2018. [Google Scholar] [CrossRef]
- Möller-Levet, C.S.; Archer, S.N.; Bucca, G.; Laing, E.E.; Slak, A.; Kabiljo, R.; Lo, J.C.; Santhi, N.; von Schantz, M.; Smith, C.P.; et al. Effects of insufficient sleep on circadian rhythmicity and expression amplitude of the human blood transcriptome. Proc. Natl. Acad. Sci. USA 2013, 110, E1132–E1141. [Google Scholar] [CrossRef]
- Franken, P.; Dijk, D.J. Sleep and circadian rhythmicity as entangled processes serving homeostasis. Nat. Rev. Neurosci. 2024, 25, 43–59. [Google Scholar] [CrossRef]
- Lu, Y.; Liu, B.; Ma, J.; Yang, S.; Huang, J. Disruption of Circadian Transcriptome in Lung by Acute Sleep Deprivation. Front. Genet. 2021, 12, 664334. [Google Scholar] [CrossRef]
- Brüning, F.; Noya, S.B.; Bange, T.; Koutsouli, S.; Rudolph, J.D.; Tyagarajan, S.K.; Cox, J.; Mann, M.; Brown, S.A.; Robles, M.S. Sleep-wake cycles drive daily dynamics of synaptic phosphorylation. Science 2019, 366, eaav3617. [Google Scholar] [CrossRef]
- Barclay, J.L.; Husse, J.; Bode, B.; Naujokat, N.; Meyer-Kovac, J.; Schmid, S.M.; Lehnert, H.; Oster, H. Circadian desynchrony promotes metabolic disruption in a mouse model of shiftwork. PLoS ONE 2012, 7, e37150. [Google Scholar] [CrossRef]
- Kim, S.J.; Hotta-Hirashima, N.; Asano, F.; Kitazono, T.; Iwasaki, K.; Nakata, S.; Komiya, H.; Asama, N.; Matsuoka, T.; Fujiyama, T.; et al. Kinase signalling in excitatory neurons regulates sleep quantity and depth. Nature 2022, 612, 512–518. [Google Scholar] [CrossRef]
- Parhizkar, S.; Gent, G.; Chen, Y.; Rensing, N.; Gratuze, M.; Strout, G.; Sviben, S.; Tycksen, E.; Zhang, Q.; Gilmore, P.E.; et al. Sleep deprivation exacerbates microglial reactivity and Aβ deposition in a TREM2-dependent manner in mice. Sci. Transl. Med. 2023, 15, eade6285. [Google Scholar] [CrossRef]
- Fenzl, T.; Romanowski, C.P.N.; Flachskamm, C.; Honsberg, K.; Boll, E.; Hoehne, A.; Kimura, M. Fully automated sleep deprivation in mice as a tool in sleep research. J. Neurosci. Methods 2007, 166, 229–235. [Google Scholar] [CrossRef]
- Naylor, E.; Harmon, H.; Gabbert, S.; Johnson, D. Automated sleep deprivation: Simulated gentle handling using a yoked control. Sleep 2010, 12, 5–12. [Google Scholar]
- Naylor, E.; Aillon, D.V.; Barrett, B.S.; Wilson, G.S.; Johnson, D.A.; Johnson, D.A.; Harmon, H.P.; Gabbert, S.; Petillo, P.A. Lactate as a biomarker for sleep. Sleep 2012, 35, 1209–1222. [Google Scholar] [CrossRef]
- Ahnaou, A.; Raeymaekers, L.; Steckler, T.; Drinkenbrug, W.H. Relevance of the metabotropic glutamate receptor (mGluR5) in the regulation of NREM-REM sleep cycle and homeostasis: Evidence from mGluR5 (-/-) mice. Behav. Brain Res. 2015, 282, 218–226. [Google Scholar] [CrossRef]
- Tahara, Y.; Kuroda, H.; Saito, K.; Nakajima, Y.; Kubo, Y.; Ohnishi, N.; Seo, Y.; Otsuka, M.; Fuse, Y.; Ohura, Y.; et al. In vivo monitoring of peripheral circadian clocks in the mouse. Curr. Biol. 2012, 22, 1029–1034. [Google Scholar] [CrossRef]
- Zhang, Z.; Cheng, L.; Ma, J.; Wang, X.; Zhao, Y. Chronic Cold Exposure Leads to Daytime Preference in the Circadian Expression of Hepatic Metabolic Genes. Front. Physiol. 2022, 13, 865627. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Y.; Shi, C.; Huang, Z.; Zhang, Y.; Li, S.; Li, Y.; Ye, J.; Yu, C.; Li, Z.; et al. SOAPnuke: A MapReduce acceleration-supported software for integrated quality control and preprocessing of high-throughput sequencing data. Gigascience 2018, 7, 1–6. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2014; Volume 1. [Google Scholar]
- Liao, Y.; Smyth, G.K.; Shi, W. The R package Rsubread is easier, faster, cheaper and better for alignment and quantification of RNA sequencing reads. Nucleic Acids Res. 2019, 47, e47. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Hughes, M.E.; Hogenesch, J.B.; Kornacker, K. JTK_CYCLE: An efficient nonparametric algorithm for detecting rhythmic components in genome-scale data sets. J. Biol. Rhythm. 2010, 25, 372–380. [Google Scholar] [CrossRef]
- Whittaker, D.S.; Akhmetova, L.; Carlin, D.; Romero, H.; Welsh, D.K.; Colwell, C.S.; Desplats, P. Circadian modulation by time-restricted feeding rescues brain pathology and improves memory in mouse models of Alzheimer’s disease. Cell Metab. 2023, 35, 1704–1721.e1706. [Google Scholar] [CrossRef]
- Parsons, R.; Parsons, R.; Garner, N.; Oster, H.; Rawashdeh, O. CircaCompare: A method to estimate and statistically support differences in mesor, amplitude and phase, between circadian rhythms. Bioinformatics 2020, 36, 1208–1212. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Đukanović, N.; La Spada, F.; Emmenegger, Y.; Niederhäuser, G.; Preitner, F.; Franken, P. Depriving Mice of Sleep also Deprives of Food. Clocks Sleep 2022, 4, 37–51. [Google Scholar] [CrossRef]
- Hoekstra, M.M.; Jan, M.; Katsioudi, G.; Emmenegger, Y.; Franken, P. The sleep-wake distribution contributes to the peripheral rhythms in PERIOD-2. Elife 2021, 10, e69773. [Google Scholar] [CrossRef]
- Curie, T.; Maret, S.; Emmenegger, Y.; Franken, P. In Vivo Imaging of the Central and Peripheral Effects of Sleep Deprivation and Suprachiasmatic Nuclei Lesion on PERIOD-2 Protein in Mice. Sleep 2015, 38, 1381–1394. [Google Scholar] [CrossRef]
- Aviram, R.; Dandavate, V.; Manella, G.; Golik, M.; Asher, G. Ultradian rhythms of AKT phosphorylation and gene expression emerge in the absence of the circadian clock components Per1 and Per2. PLoS Biol. 2021, 19, e3001492. [Google Scholar] [CrossRef]
- Idevall-Hagren, O.; Dickson, E.J.; Hille, B.; Toomre, D.K.; De Camilli, P. Optogenetic control of phosphoinositide metabolism. Proc. Natl. Acad. Sci. USA 2012, 109, E2316–E2323. [Google Scholar] [CrossRef]
- Chaput, J.P.; McHill, A.W.; Cox, R.C.; Broussard, J.L.; Dutil, C.; da Costa, B.G.G.; Sampasa-Kanyinga, H.; Wright, K.P., Jr. The role of insufficient sleep and circadian misalignment in obesity. Nat. Rev. Endocrinol. 2023, 19, 82–97. [Google Scholar] [CrossRef]
- Chaput, J.P.; Dutil, C.; Featherstone, R.; Ross, R.; Giangregorio, L.; Saunders, T.J.; Janssen, I.; Poitras, V.J.; Kho, M.E.; Ross-White, A.; et al. Sleep duration and health in adults: An overview of systematic reviews. Appl. Physiol. Nutr. Metab. 2020, 45, S218–S231. [Google Scholar] [CrossRef]
- Boege, H.L.; Bhatti, M.Z.; St-Onge, M.P. Circadian rhythms and meal timing: Impact on energy balance and body weight. Curr. Opin. Biotechnol. 2021, 70, 1–6. [Google Scholar] [CrossRef]
- Diessler, S.; Jan, M.; Emmenegger, Y.; Guex, N.; Middleton, B.; Skene, D.J.; Ibberson, M.; Burdet, F.; Götz, L.; Pagni, M.; et al. A systems genetics resource and analysis of sleep regulation in the mouse. PLoS Biol. 2018, 16, e2005750. [Google Scholar] [CrossRef]
- Zitting, K.M.; Vujovic, N.; Yuan, R.K.; Isherwood, C.M.; Medina, J.E.; Wang, W.; Buxton, O.M.; Williams, J.S.; Czeisler, C.A.; Duffy, J.F. Human Resting Energy Expenditure Varies with Circadian Phase. Curr. Biol. 2018, 28, 3685–3690.e3683. [Google Scholar] [CrossRef]
- Ohdo, S.; Koyanagi, S.; Matsunaga, N. Chronopharmacological strategies: Intra- and inter-individual variability of molecular clock. Adv. Drug Deliv. Rev. 2010, 62, 885–897. [Google Scholar] [CrossRef]
- Jung, C.M.; Melanson, E.L.; Frydendall, E.J.; Perreault, L.; Eckel, R.H.; Wright, K.P. Energy expenditure during sleep, sleep deprivation and sleep following sleep deprivation in adult humans. J. Physiol. 2011, 589, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Markwald, R.R.; Melanson, E.L.; Smith, M.R.; Higgins, J.; Perreault, L.; Eckel, R.H.; Wright, K.P., Jr. Impact of insufficient sleep on total daily energy expenditure, food intake, and weight gain. Proc. Natl. Acad. Sci. USA 2013, 110, 5695–5700. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Zhuo, K.; Wang, Y.; Wang, X.; Zhao, Y. Prolonged Sleep Deprivation Induces a Reprogramming of Circadian Rhythmicity with the Hepatic Metabolic Transcriptomic Profile. Biology 2024, 13, 532. https://doi.org/10.3390/biology13070532
Liu S, Zhuo K, Wang Y, Wang X, Zhao Y. Prolonged Sleep Deprivation Induces a Reprogramming of Circadian Rhythmicity with the Hepatic Metabolic Transcriptomic Profile. Biology. 2024; 13(7):532. https://doi.org/10.3390/biology13070532
Chicago/Turabian StyleLiu, Shiyan, Kailin Zhuo, Yiming Wang, Xiaomei Wang, and Yingying Zhao. 2024. "Prolonged Sleep Deprivation Induces a Reprogramming of Circadian Rhythmicity with the Hepatic Metabolic Transcriptomic Profile" Biology 13, no. 7: 532. https://doi.org/10.3390/biology13070532