Melatonin and Sleep-Wake Rhythms before and after Ocular Lens Replacement in Elderly Humans

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Subjects
2.2. Experimental Design
2.3. Data Analysis
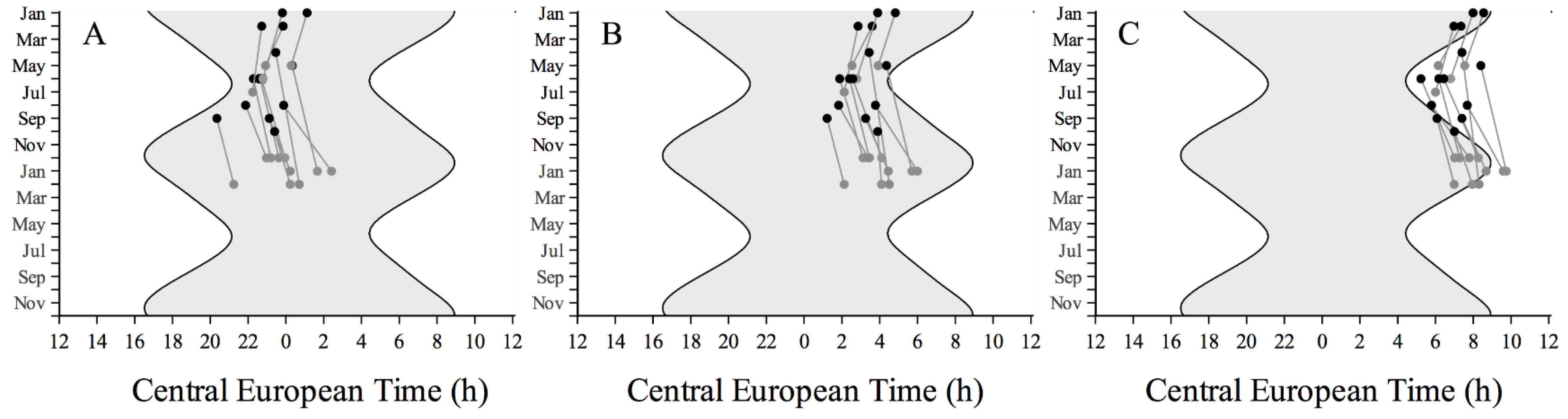
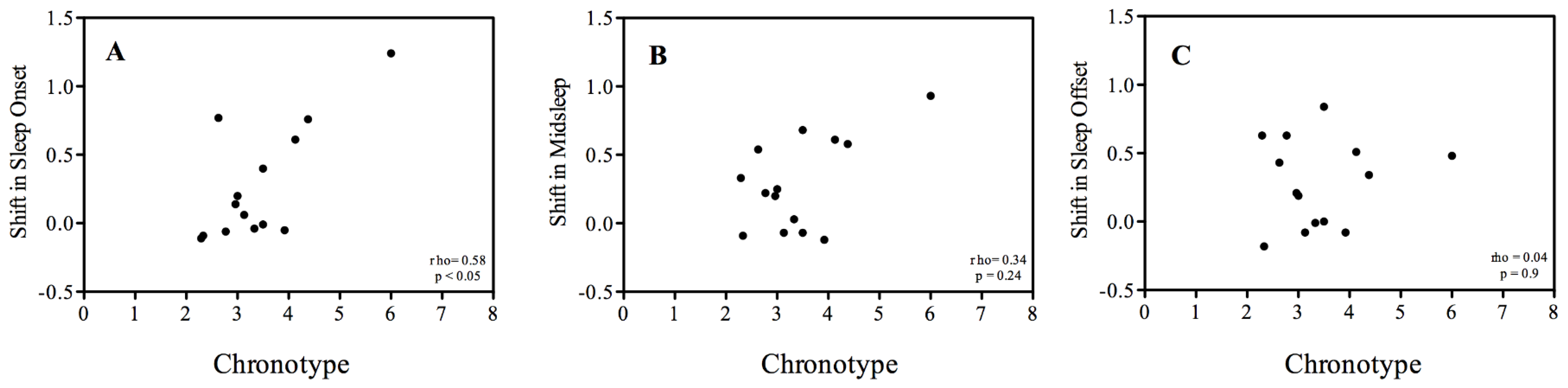
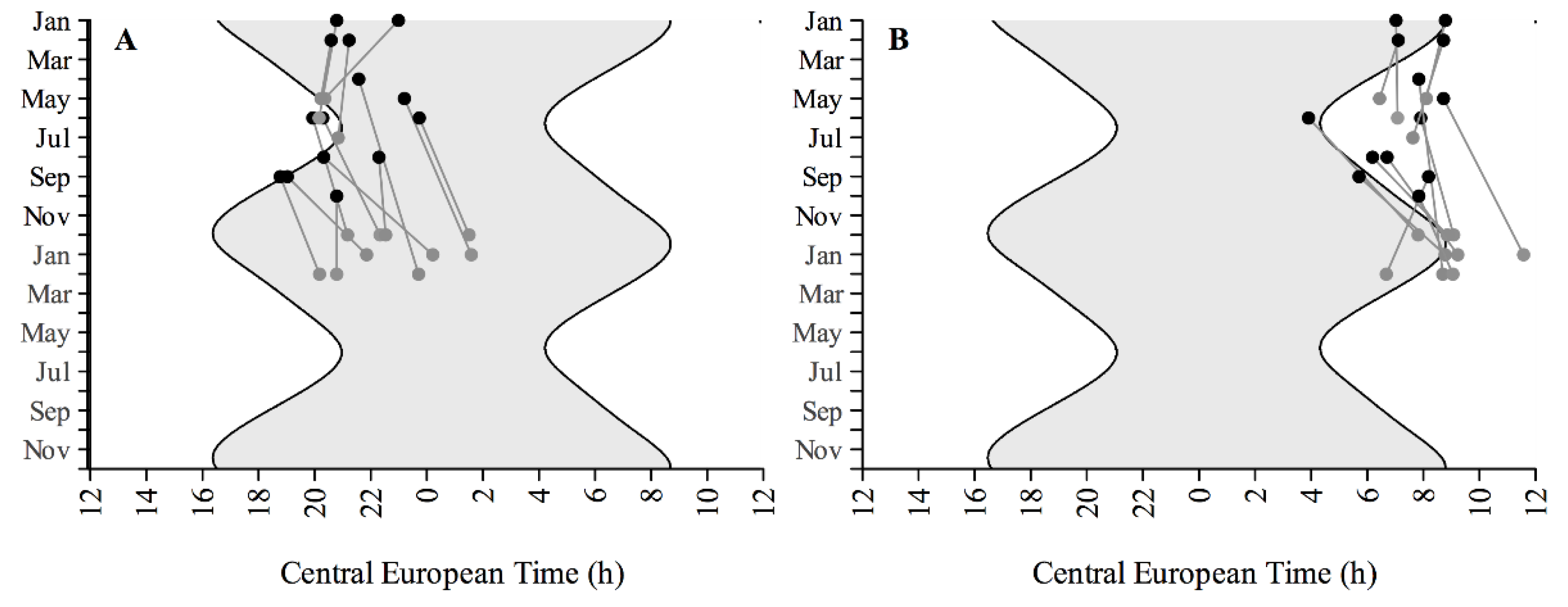
3. Results
3.1. Subjects
3.2. Sleep-Wake Rhythms



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Before Surgery | After Surgery | F | p | Difference | |
|---|---|---|---|---|---|
| Sleep Onset (CET) | 23:12 ± 1:06 | 23:28 ± 1:13 | 5.8 | < 0.05 | 16 ± 25 min |
| Midsleep (CET) | 03:10 ± 51 min | 3:28 ± 59 min | 10.3 | < 0.01 | 17 ± 19 min |
| Sleep Offset (CET) | 7:08 ± 49 min | 7:25 ± 50 min | 10.7 | < 0.01 | 17 ± 10 min |
| Sleep Duration (CET) | 7:56 ± 38 min | 7:56 ± 40 min | 0.006 | 0.94 | 0.5 ± 24 min |
| Sleep Onset Latency (min) | 10.5 ± 6 | 13 ± 9.8 | 2.26 | 0.16 | 2.5 ± 6 min |
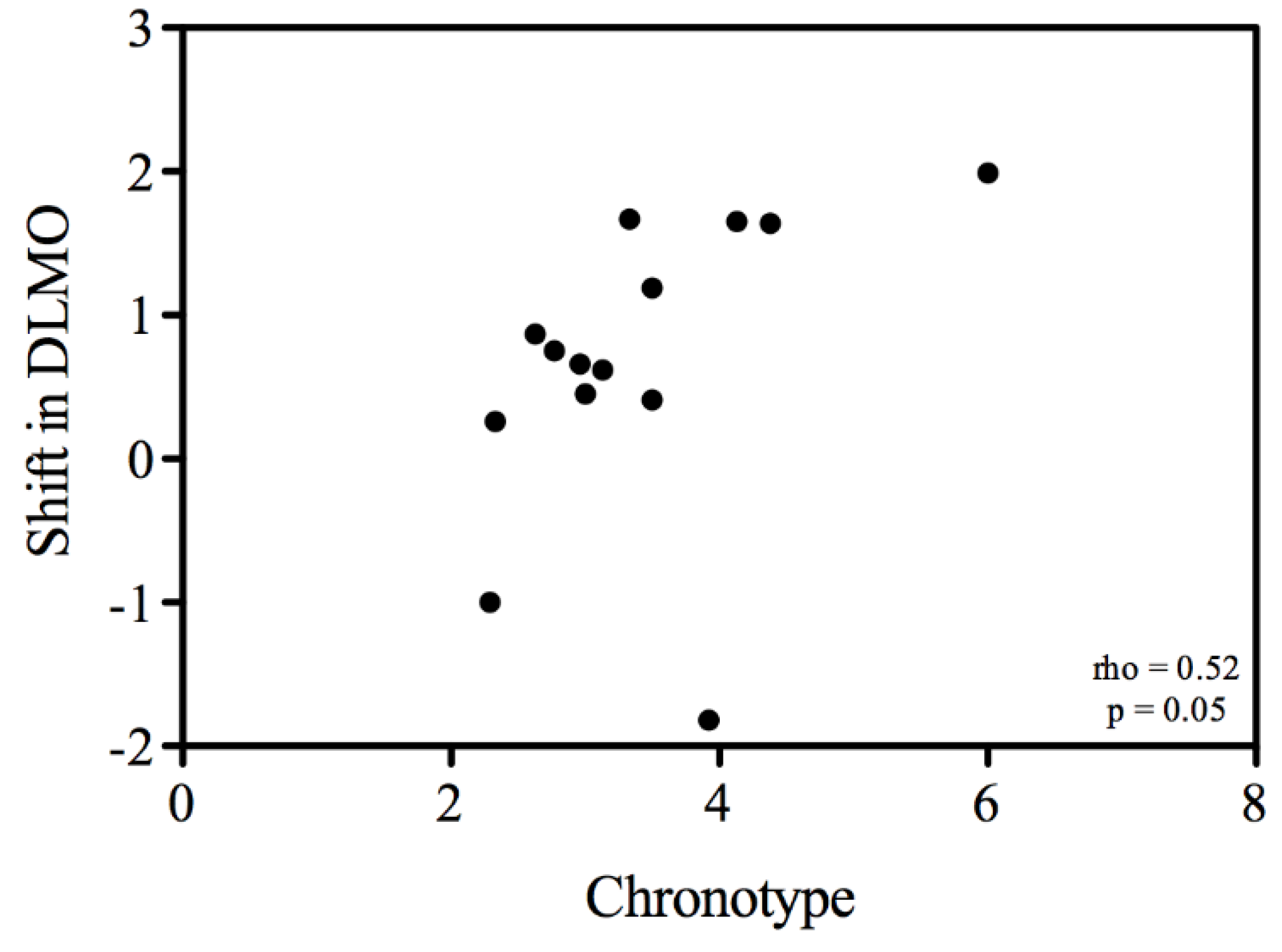
3.3. Melatonin Rhythms


| Before Surgery | After Surgery | F value | p Value | Difference | |
|---|---|---|---|---|---|
| DLMO | 21:14 h ± 1:30 h | 21:54 h ± 1:41 h | 5.69 | < 0.05 | 40 min ± 1:03 h |
| DLMOff | 07:20 h ± 1:12 h | 8:07 h ± 1:08 h | 4.57 | 0.05 | 46 min ± 1:19 h |
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brainard, G.C.; Hanifin, J.P.; Greeson, J.M.; Byrne, B.; Glickman, G.; Gerner, E.; Rollag, M.D. Action spectrum for melatonin regulation in humans: Evidence for a novel circadian photoreceptor. J. Neurosci. 2001, 21, 6405–6412. [Google Scholar] [PubMed]
- Warman, V.L.; Dijk, D.J.; Warman, G.R.; Arendt, J.; Skene, D.J. Phase advancing human circadian rhythms with short wavelength light. Neurosci. Lett. 2003, 342, 37–40. [Google Scholar] [CrossRef]
- Lockley, S.W.; Brainard, G.C.; Czeisler, C.A. High sensitivity of the human circadian melatonin rhythm to resetting by short wavelength light. J. Clin. Endocrinol. Metab. 2003, 88, 4502–4502. [Google Scholar] [CrossRef] [PubMed]
- Lockley, S.W.; Evans, E.E.; Scheer, F.A.J.L.; Brainard, G.C.; Czeisler, C.A.; Aeschbach, D. Short-wavelength sensitivity for the direct effects of light on alertness, vigilance, and the waking electroencephalogram in humans. Sleep 2006, 29, 161–168. [Google Scholar] [PubMed]
- Cajochen, C.; Munch, M.; Kobialka, S.; Kräuchi, K.; Steiner, R.; Oelhafen, P.; Orgul, S.; Wirz-Justice, A. High sensitivity of human melatonin, alertness, thermoregulation, and heart rate to short wavelength light. J. Clin. Endocrinol. Metab. 2005, 90, 1311–1316. [Google Scholar] [CrossRef] [PubMed]
- Revell, V.L.; Arendt, J.; Terman, M.; Skene, D.J. Short-wavelength sensitivity of the human circadian system to phase-advancing light. J. Biol. Rhythms 2005, 20, 270–272. [Google Scholar] [CrossRef] [PubMed]
- Provencio, I.; Rollag, M.D.; Castrucci, A.M. Photoreceptive net in the mammalian retina. This mesh of cells may explain how some blind mice can still tell day from night. Nature 2002. [Google Scholar] [CrossRef]
- Provencio, I.; Rodriguez, I.R.; Jiang, G.; Hayes, W.P.; Moreira, E.F.; Rollag, M.D. A novel human opsin in the inner retina. J. Neurosci. 2000, 20, 600–605. [Google Scholar] [PubMed]
- Hattar, S.; Liao, H.W.; Takao, M.; Berson, D.M.; Yau, K.W. Melanopsin-containing retinal ganglion cells: Architecture, projections, and intrinsic photosensitivity. Science 2002, 295, 1065–1070. [Google Scholar] [CrossRef] [PubMed]
- Berson, D.M.; Dunn, F.A.; Takao, M. Phototransduction by retinal ganglion cells that set the circadian clock. Science 2002, 295, 1070–1073. [Google Scholar] [CrossRef] [PubMed]
- Hattar, S.; Kumar, M.; Park, A.; Tong, P.; Tung, J.; Yau, K.W.; Berson, D.M. Central projections of melanopsin-expressing retinal ganglion cells in the mouse. J. Comp. Neurol. 2006, 497, 326–349. [Google Scholar] [CrossRef] [PubMed]
- Van Someren, E.J.W. Circadian and sleep disturbances in the elderly. Exp. Gerontol. 2000, 35, 1229–1237. [Google Scholar] [CrossRef]
- Duffy, J.F.; Zeitzer, J.M.; Czeisler, C.A. Decreased sensitivity to phase-delaying effects of moderate intensity light in older subjects. Neurobiol. Aging 2007, 28, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Klerman, E.B.; Duffy, J.F.; Dijk, D.J.; Czeisler, C.A. Circadian phase resetting in older people by ocular bright light exposure. J. Investig. Med. 2001, 49, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.F.; Zeitzer, J.M.; Rimmer, D.W.; Klerman, E.B.; Dijk, D.-J.; Czeisler, C.A. Peak of circadian melatonin rhythm occurs later within the sleep of older subjects. Am. J. Physiol. Endocrinol. Metab. 2001, 282, E297–E303. [Google Scholar] [CrossRef] [PubMed]
- Lewy, A.J. Melatonin and Human Chronobiology. Cold Spring Harb. Symp. Quant. Biol. 2007, 72, 623–636. [Google Scholar] [CrossRef] [PubMed]
- Van Norren, D.; Vos, J.J. Spectral transmission of the human ocular media. Vis. Res. 1974, 14, 1237–1244. [Google Scholar] [CrossRef]
- Bonnel, S.; Mohand-Said, S.; Sahel, J.-A. The aging of the retina. Exp. Gerontol. 2003, 38, 825–831. [Google Scholar] [CrossRef]
- Cavallotti, C.; Artico, M.; Pescosolido, N.; Feher, J. Age-related changes in rat retina. Jpn. J. Ophtalmol. 2001, 45, 68–75. [Google Scholar] [CrossRef]
- Katz, M.L.; Robison, W.G. Evidence of cell loss from the rat retina during senescence. Exp. Eye Res. 1986, 42, 293–304. [Google Scholar] [CrossRef]
- Deacon, S.; Arendt, J. Posture influences melatonin concentrations in plasma and saliva in humans. Neurosci. Lett. 1994, 167, 191–194. [Google Scholar] [CrossRef]
- Van de Kraats, J.; van Norren, D. Directional and nondirectional spectral reflection from the human fovea. J. Biomed. Opt. 2008. [Google Scholar] [CrossRef] [PubMed]
- Giménez, M.C.; Kanis, M.J.; Beersma, D.G.; van der Pol, B.A.; van Norren, D.; Gordijn, M.C. In vivo quantification of the retinal reflectance spectral composition in elderly subjects before and after cataract surgery: Implications for the non-visual effects of light. J. Biol. Rhythms 2010, 25, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Roenneberg, T.; Wirz-Justice, A.; Merrow, M. Life between clocks: Daily temporal patterns of human chronotypes. J. Biol. Rhythms 2003, 18, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Zavada, A.; Gordijn, M.C.M.; Beersma, D.G.M.; Daan, S.; Roenneberg, T. Comparison of the Munich chronotype questionnaire with the Horne-Ostberg’s morningness-eveningness score. Chronobiol. Int. 2005, 22, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Van Someren, E.J.; Swaab, D.F.; Colenda, C.C.; Cohen, W.; McCall, W.V.; Rosenquist, P.B. Bright light therapy: Improved sensitivity to its effects on rest-activity rhythms in Alzheimer patients by application of nonparametric methods. Chronobiol. Int. 1999, 16, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Witting, W.; Kwa, I.H.; Eikelenboom, P.; Mirmiran, M.; Swaab, D.F. Alterations in the circadian rest-activity rhythm in aging and Alzheimer’s disease. Biol. Psychiatry 1990, 27, 563–572. [Google Scholar] [CrossRef]
- Van Someren, E.J.W.; Nagtegaal, E. Improving melatonin circadian phase estimates. Sleep Med. 2007, 8, 590–601. [Google Scholar] [CrossRef] [PubMed]
- Lewy, A.J.; Cutler, N.L.; Sack, R.L. The endogenous melatonin profile as a marker for circadian phase position. J. Biol. Rhythms 1999, 14, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Kantermann, T.; Juda, M.; Merrow, M.; Roenneberg, T. The human circadian clock’s seasonal adjustment is disrupted by daylight saving time. Curr. Biol. 2007, 17, 1996–2000. [Google Scholar] [CrossRef] [PubMed]
- Hut, R.A. CircWave Version 1.4. Available online: http://www.euclock.org/results/item/circ-wave.html (accessed on 11 November 2015).
- Schmoll, C.; Lascaratos, G.; Dhillon, B.; Skene, D.; Riha, R.L. The role of retinal regulation of sleep in health and disease. Sleep Med. Rev. 2011, 15, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Charman, W.N. Age, lens transmittance, and the possible effects of light on melatonin suppression. Ophthalmic Physiol. Opt. 2003, 23, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Turner, P.L.; Mainster, M.A. Circadian photoreception: Ageing and the eye’s important role in systemic health. Br. J. Ophthalmol. 2008, 92, 1439–1444. [Google Scholar] [CrossRef] [PubMed]
- Turner, P.L.; van Someren, E.J.W.; Mainster, M.A. Sleep Med. Rev. 2010, 14, 269–280.
- Asplund, R.; Lindblad, B.E. Sleep and sleepiness 1 and 9 months after cataract surgery. Arch. Gerontol. Geriatr. 2004, 38, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Hosoe, K.; Hamada, T.; Morita, T. Change in sleep state of the elderly before and after cataract surgery. J. Physiol. Anthropol. 2010, 29, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Kessel, L.; Siganos, G.; Jørgensen, T.; Larsen, M. Sleep disturbances are related to decreased transmission of blue light to the retina caused by lens yellowing. Sleep 2011, 34, 1215–1219. [Google Scholar] [CrossRef] [PubMed]
- Alexander, I.; Cuthbertson, F.M.; Ratnarajan, G.; Safa, R.; Mellington, F.E.; Foster, R.G.; Downes, S.M.; Wulff, K. Impact of cataract surgery on sleep in patients receiving either ultraviolet-blocking or blue-filtering intraocular lens implants. Invest. Ophthal. Vis. Sci. 2014, 55, 4999–5004. [Google Scholar] [CrossRef] [PubMed]
- Brøndsted, A.E.; Sander, B.; Haargaard, B.; Lund-Andersen, H.; Jennum, P.; Gammeltoft, S.; Kessel, L. The effect of cataract surgery on circadian photoentrainment: A randomized trial of blue-blocking versus neutral intraocular lenses. Ophthalmology 2015, 122, 2115–2124. [Google Scholar] [CrossRef] [PubMed]
- Roenneberg, T.; Kuehnle, T.; Juda, M.; Kantermann, T.; Allebrandt, K.; Gordijn, M.; Merrow, M. Epidemiology of the human circadian clock. Sleep Med. Rev. 2007, 11, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Miles, L.E.; Dement, W.C. Sleep and aging. Sleep 1980, 3, 1–220. [Google Scholar] [PubMed]
- Myers, B.L.; Badia, P. Changes in circadian rhythms and sleep quality with aging: Mechanisms and interventions. Neurosci. Biobehav. Rev. 1995, 9, 553–571. [Google Scholar] [CrossRef]
- Duffy, J.F.; Dijk, D.J.; Klerman, E.B.; Czeisler, C.A. Later endogenous circadian temperature nadir relative to an earlier wake time in older people. Am. J. Physiol. 1998, 275, R1478–R1487. [Google Scholar] [PubMed]
- Duffy, J.F.; Czeisler, C.A. Age-related change in the relationship between circadian period, circadian phase, and diurnal preference in humans. Neurosci. Lett. 2002, 318, 117–120. [Google Scholar] [CrossRef]
- Cajochen, C.; Münch, M.; Knoblauch, V.; Blatter, K.; Wirz-Justice, A. Age-related changes in the circadian and homeostatic regulation of human sleep. Chronobiol. Int. 2006, 23, 461–474. [Google Scholar] [CrossRef] [PubMed]
- Czeisler, C.A.; Duffy, J.F.; Shanahan, T.L.; Brown, E.N.; Mitchell, J.F.; Rimmer, D.W.; Ronda, J.M.; Silva, E.J.; Allan, J.S.; Emens, J.S.; et al. Stability, precision, and near-24-h period of the human circadian pacemaker. Science 1999, 284, 2177–2181. [Google Scholar] [CrossRef] [PubMed]
- Boivin, D.B.; Duffy, J.F.; Kronauer, R.E.; Czeisler, C.A. Sleep, hormones, and circadian rhythms throughout the menstrual cycle in healthy women and women with premenstrual dysphoric disorder. Nature 1996, 2010, 540–542. [Google Scholar] [CrossRef] [PubMed]
- Zeitzer, J.M.; Khalsa, S.B.S.; Boivin, D.B.; Duffy, J.F.; Shanahan, T.L.; Kronauer, R.E.; Czeisler, C.A. Temporal dynamics of late-night photic stimulation of the human circadian timing system. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R839–R844. [Google Scholar] [CrossRef] [PubMed]
- Cajochen, C. Alerting effects of light. Sleep Med. Rev. 2007, 11, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Goulet, G.; Mongrain, V.; Desrosiers, C.; Paquet, J.; Dumont, M. Daily light exposure in morning-type and evening-type individuals. J. Biol. Rhythms 2007, 22, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Staples, V.S.L.; Archer, S.N.; Arber, S.; Skene, D.J. Daily light exposure profiles in older non-resident extreme morning and evening types. J. Sleep Res. 2009, 18, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Benloucif, S.; Green, K.; L’Hermite-Balériaux, M.; Weintraub, S.; Wolfe, L.F.; Zee, P.C. Responsiveness of the aging circadian clock to light. Neurobiol. Aging 2006, 27, 1870–1879. [Google Scholar] [CrossRef] [PubMed]
- Asplund, R.; Lindblad, B.E. The development of sleep in persons undergoing cataract surgery. Arch. Gerontol. Geriatr. 2002, 35, 179–187. [Google Scholar] [CrossRef]
- Pallesen, S.; Nordhus, I.H.; Nielsen, G.H.; Havik, O.E.; Kvale, G.; Johnsen, B.H.; Skjøtskift, S. Prevalence of insomnia in the adult Norwegian population. Sleep 2001, 24, 771–779. [Google Scholar] [PubMed]
- Friborg, O.; Bjorvatn, B.; Amponsah, B.; Pallesen, S. Associations between seasonal variations in day length (photoperiod), sleep timing, sleep quality and mood: A comparison between Ghana (5°) and Norway (69°). J. Sleep Res. 2011, 21, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Giménez, M.C.; Beersma, D.G.M.; Bollen, P.; van der Linden, M.L.; Gordijn, M.C.M. Effects of a chronic reduction of short-wavelength light input on melatonin and sleep patterns in humans: Evidence for adaptation. Chronobiol. Int. 2014, 31, 690–697. [Google Scholar] [CrossRef] [PubMed]
- Najjar, R.P.; Chiquet, C.; Teikari, P.; Cornut, P.-L.; Claustrat, B.; Denis, P.; Cooper, H.M.; Gronfier, C. Aging of non-visual spectral sensitivity to light in humans: Compensatory mechanisms? PLoS ONE 2014, 9, e85837. [Google Scholar] [CrossRef] [PubMed]
- Farajnia, S.; Michel, S.; Deboer, T.; VanderLeest, H.T.; Houben, T.; Rohling, J.H.T.; Ramkisoensing, A.; Yasenkov, R.; Meijer, J.H. Evidence for neuronal desynchrony in the aged suprachiasmatic nucleus clock. J. Neurosci. 2012, 32, 5891–5899. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giménez, M.; Beersma, D.; Daan, S.; Pol, B.V.d.; Kanis, M.; Van Norren, D.; Gordijn, M. Melatonin and Sleep-Wake Rhythms before and after Ocular Lens Replacement in Elderly Humans. Biology 2016, 5, 12. https://doi.org/10.3390/biology5010012
Giménez M, Beersma D, Daan S, Pol BVd, Kanis M, Van Norren D, Gordijn M. Melatonin and Sleep-Wake Rhythms before and after Ocular Lens Replacement in Elderly Humans. Biology. 2016; 5(1):12. https://doi.org/10.3390/biology5010012
Chicago/Turabian StyleGiménez, Marina, Domien Beersma, Serge Daan, Bert Van der Pol, Martijn Kanis, Dick Van Norren, and Marijke Gordijn. 2016. "Melatonin and Sleep-Wake Rhythms before and after Ocular Lens Replacement in Elderly Humans" Biology 5, no. 1: 12. https://doi.org/10.3390/biology5010012
APA StyleGiménez, M., Beersma, D., Daan, S., Pol, B. V. d., Kanis, M., Van Norren, D., & Gordijn, M. (2016). Melatonin and Sleep-Wake Rhythms before and after Ocular Lens Replacement in Elderly Humans. Biology, 5(1), 12. https://doi.org/10.3390/biology5010012