Melatonin Inhibits Transforming Growth Factor-β1-Induced Epithelial–Mesenchymal Transition in AML12 Hepatocytes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Cell Treatments
2.2. Western Blot Analysis
2.3. Quantitative Real-Time Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.4. Evaluation of Intracellular ROS
2.5. Statistical Analysis
3. Results
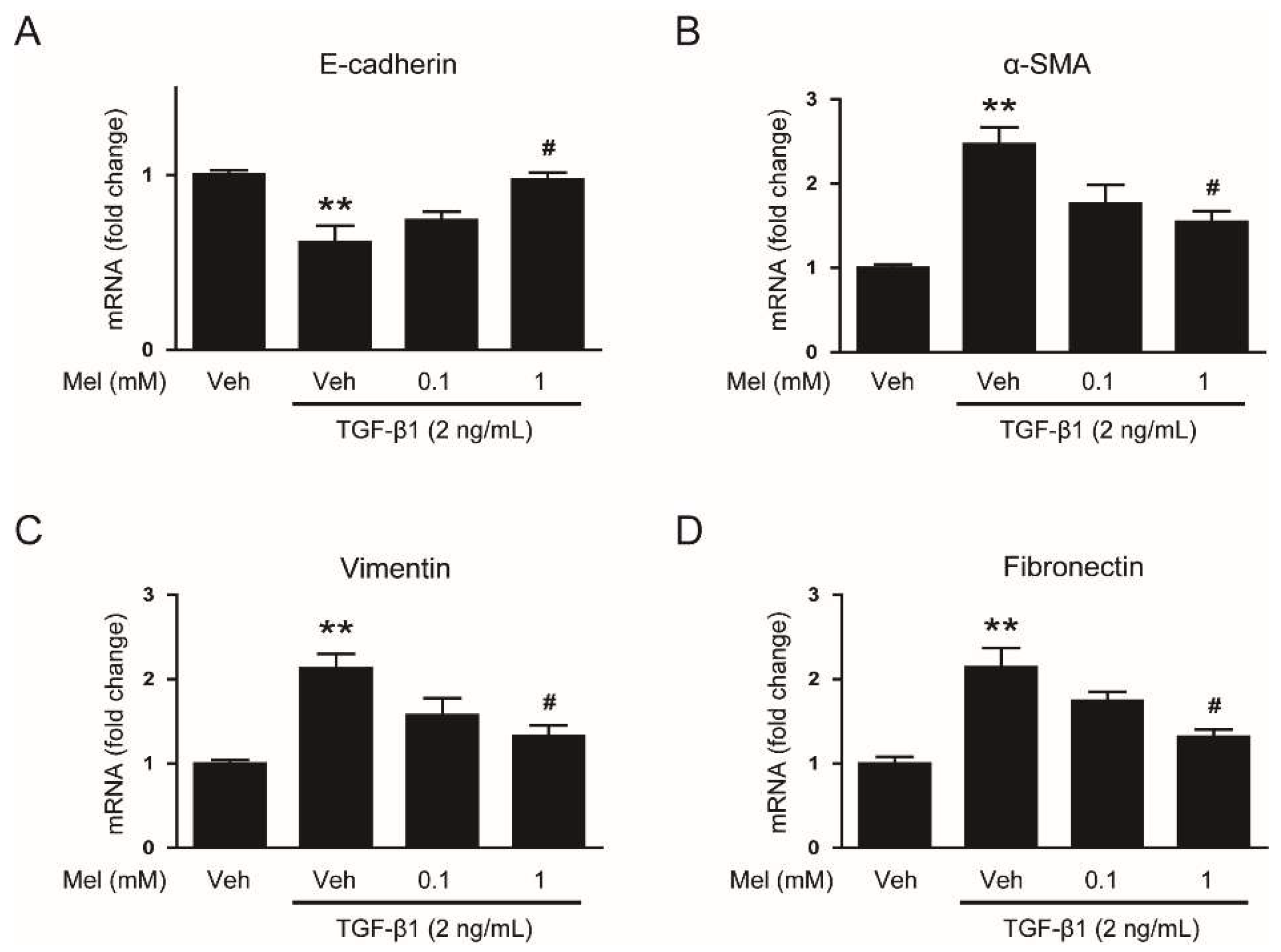
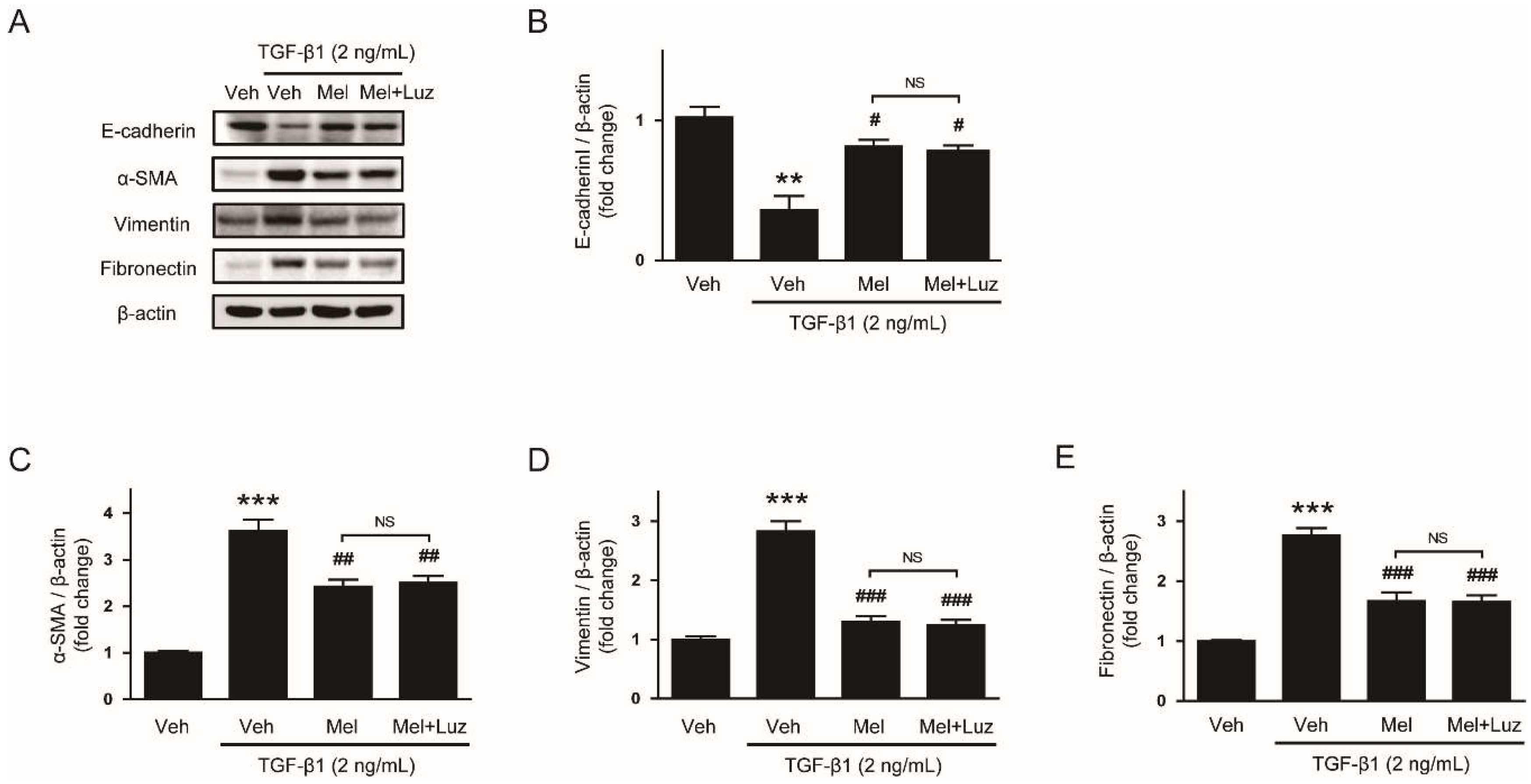
3.1. Melatonin Prevents TGF-β1-Stimulated EMT in AML12 Hepatocytes
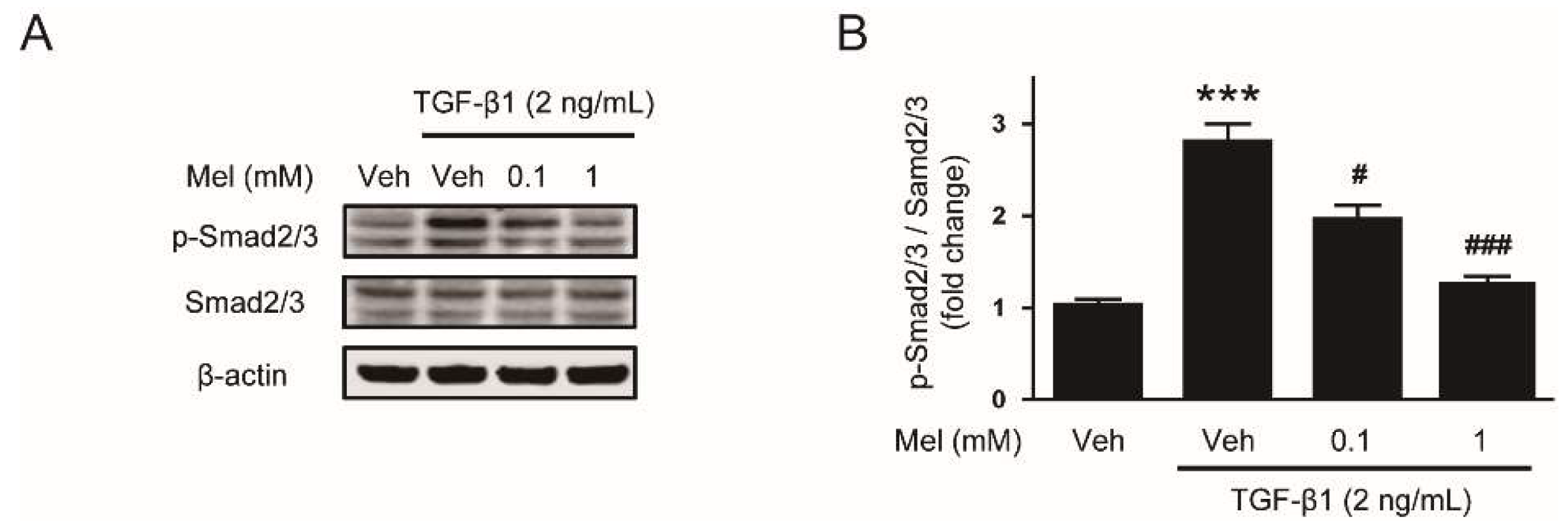
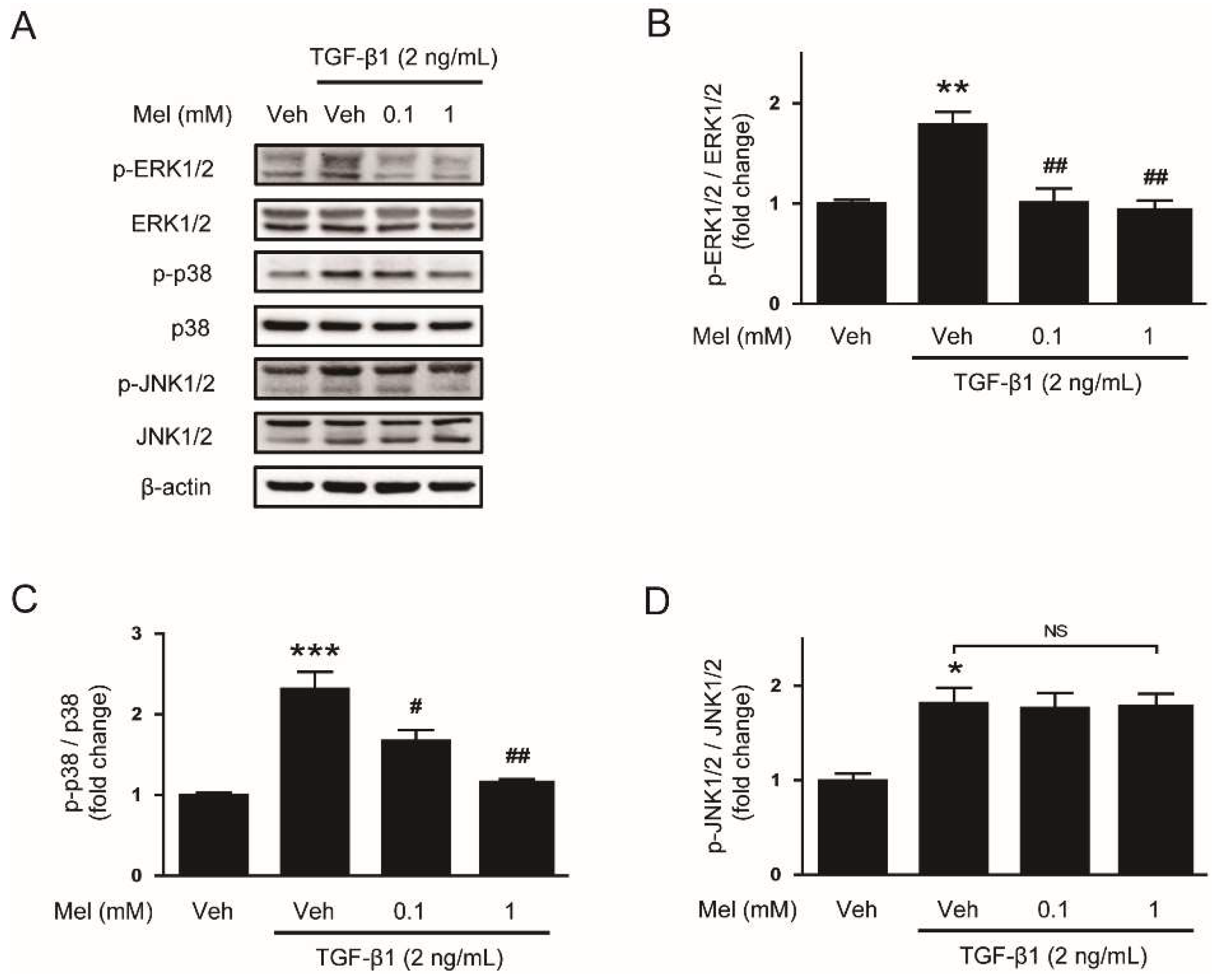
3.2. Melatonin Attenuates TGF-β1-Stimulated Smad and MAPK Signaling Pathways
3.3. Suppressive Effect of Melatonin on EMT Stimulated by TGF-β1 Is Independent of Its Membrane Receptors
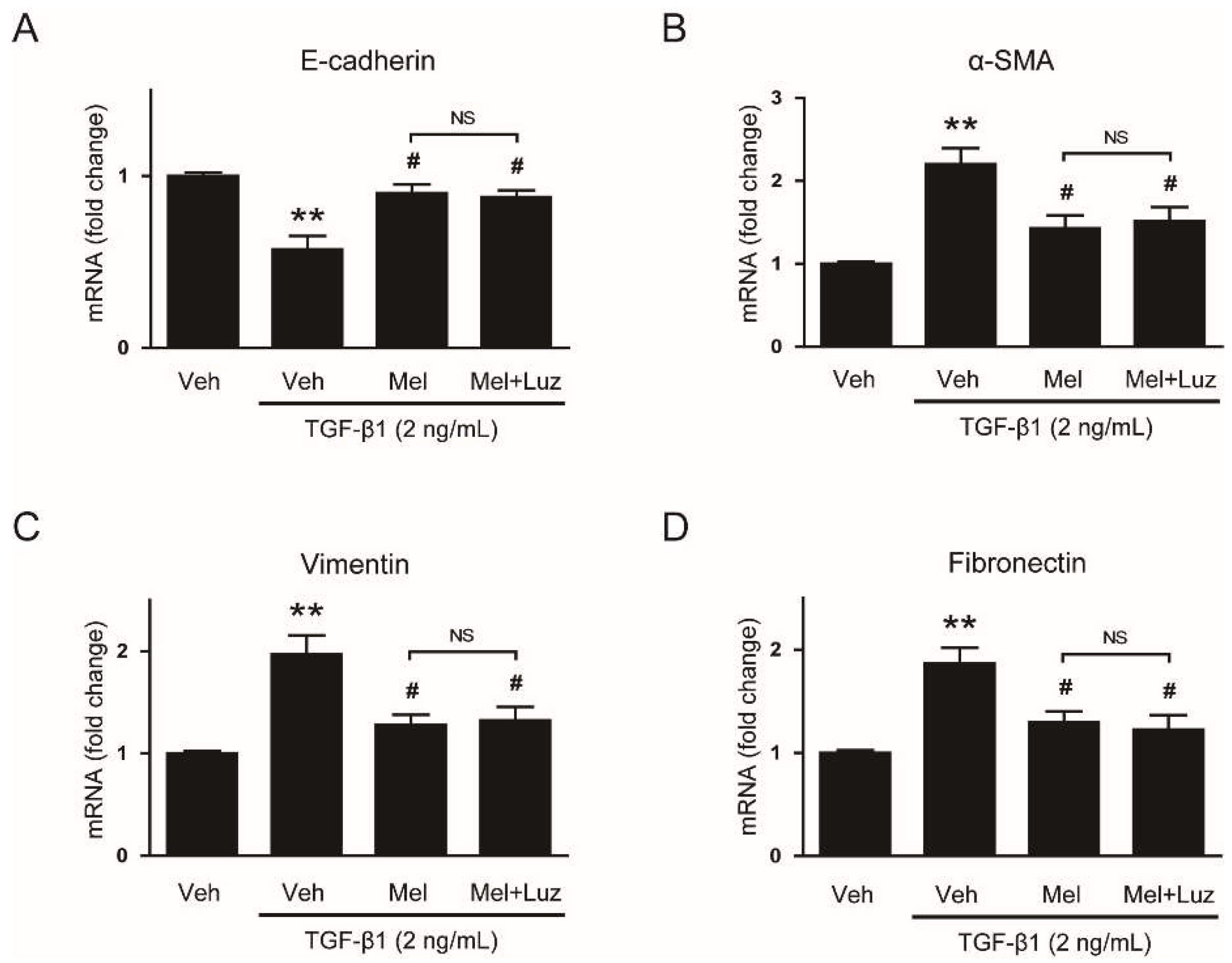
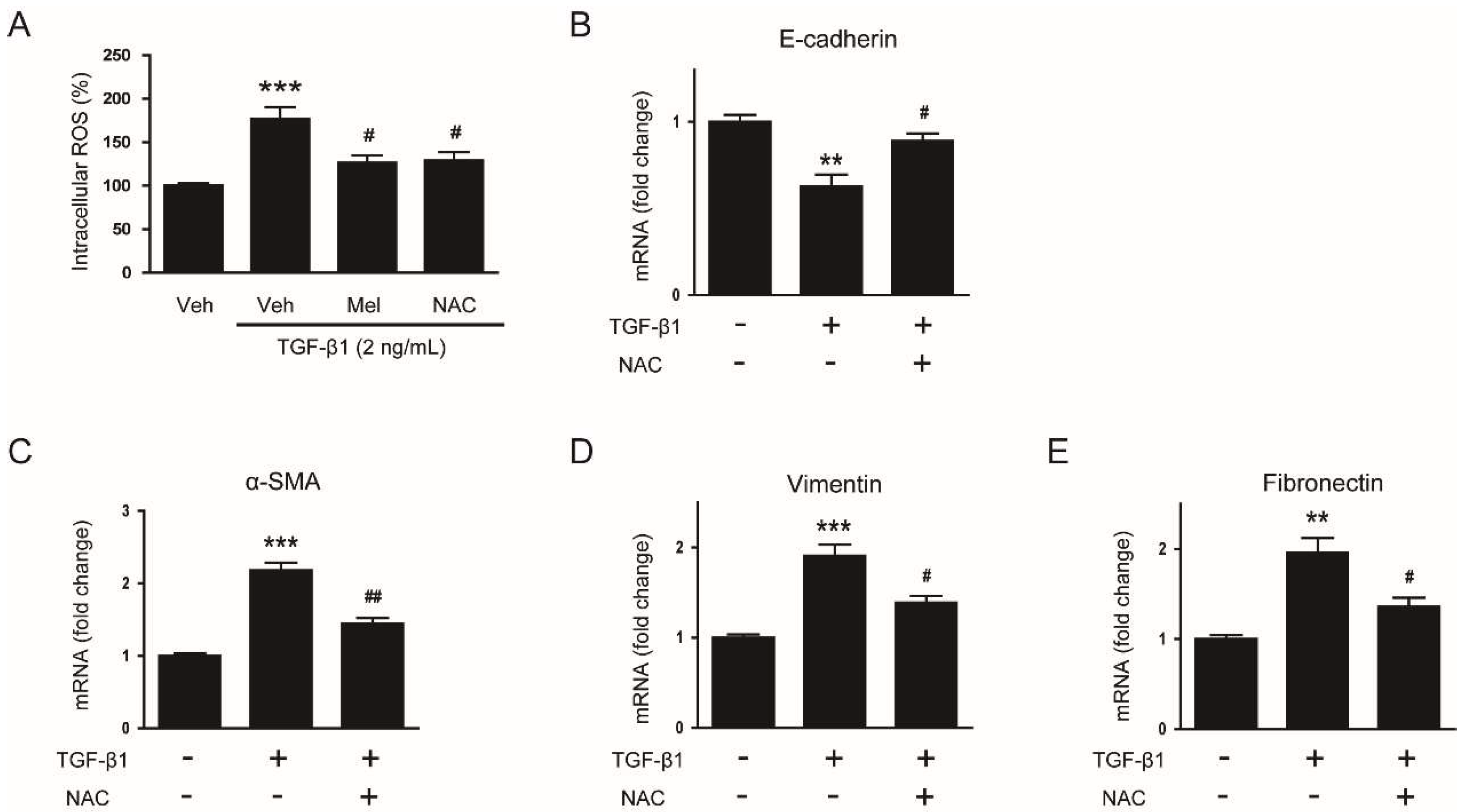
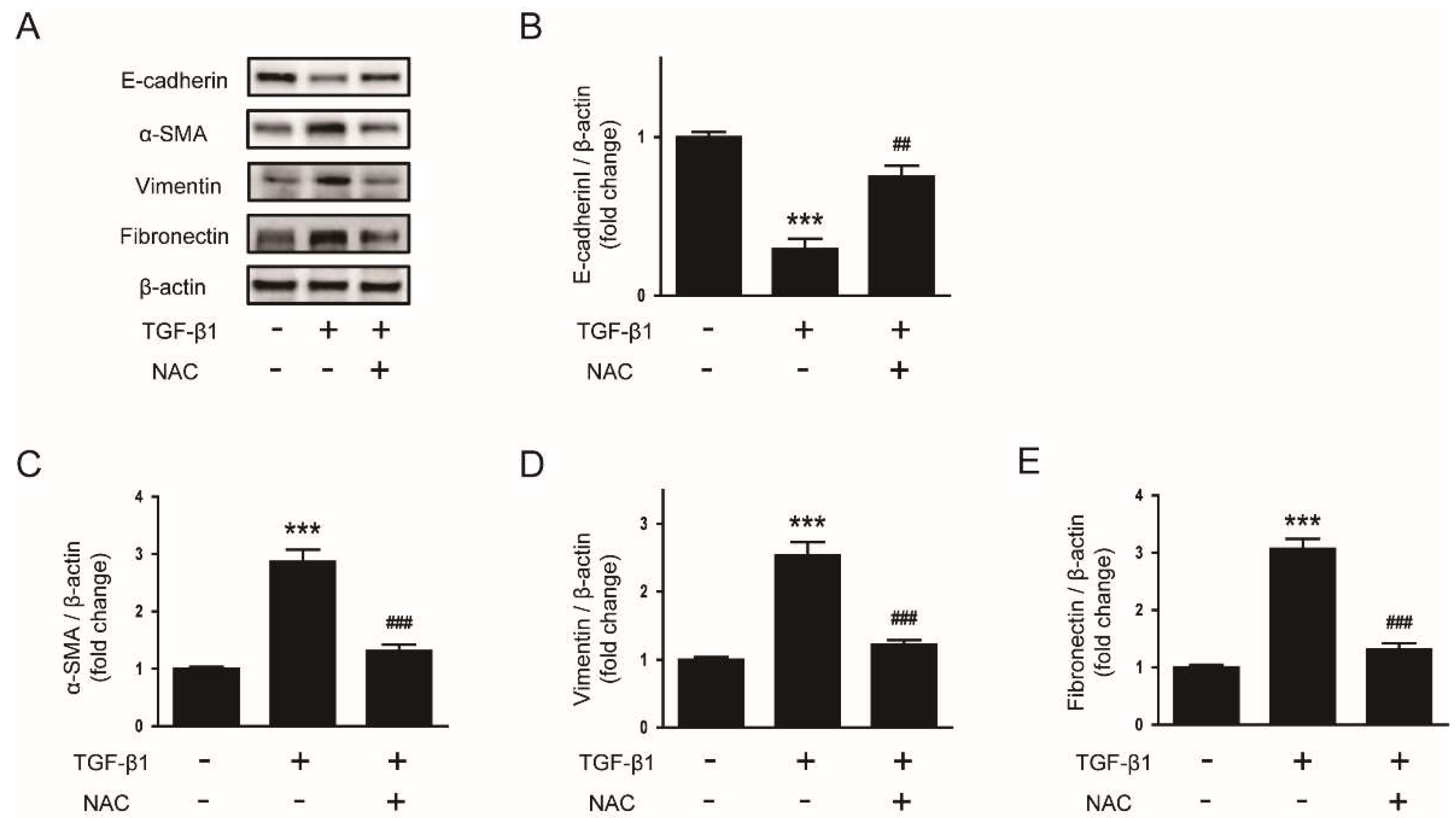
3.4. Suppressive Effect of Melatonin on EMT Stimulated by TGF-β1 Is Mediated by Suppressing ROS-Dependent Mechanisms
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mallat, A.; Lotersztajn, S. Cellular mechanisms of tissue fibrosis. 5. Novel insights into liver fibrosis. Am. J. Physiol.Cell Physiol. 2013, 305, C789–C799. [Google Scholar] [CrossRef] [PubMed]
- Zeisberg, M.; Yang, C.; Martino, M.; Duncan, M.B.; Rieder, F.; Tanjore, H.; Kalluri, R. Fibroblasts derive from hepatocytes in liver fibrosis via epithelial to mesenchymal transition. J. Biol. Chem. 2007, 282, 23337–23347. [Google Scholar] [CrossRef] [PubMed]
- Dooley, S.; Hamzavi, J.; Ciuclan, L.; Godoy, P.; Ilkavets, I.; Ehnert, S.; Ueberham, E.; Gebhardt, R.; Kanzler, S.; Geier, A.; et al. Hepatocyte-specific Smad7 expression attenuates TGF-beta-mediated fibrogenesis and protects against liver damage. Gastroenterology 2008, 135, 642–659. [Google Scholar] [CrossRef] [PubMed]
- Nitta, T.; Kim, J.S.; Mohuczy, D.; Behrns, K.E. Murine cirrhosis induces hepatocyte epithelial mesenchymal transition and alterations in survival signaling pathways. Hepatology 2008, 48, 909–919. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.S.; Diehl, A.M. Epithelial-to-mesenchymal transitions in the liver. Hepatology 2009, 50, 2007–2013. [Google Scholar] [CrossRef] [PubMed]
- Taura, K.; Miura, K.; Iwaisako, K.; Osterreicher, C.H.; Kodama, Y.; Penz-Osterreicher, M.; Brenner, D.A. Hepatocytes do not undergo epithelial-mesenchymal transition in liver fibrosis in mice. Hepatology 2010, 51, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Chu, A.S.; Diaz, R.; Hui, J.J.; Yanger, K.; Zong, Y.; Alpini, G.; Stanger, B.Z.; Wells, R.G. Lineage tracing demonstrates no evidence of cholangiocyte epithelial-to-mesenchymal transition in murine models of hepatic fibrosis. Hepatology 2011, 53, 1685–1695. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Galano, A. Melatonin: Exceeding expectations. Physiology 2014, 29, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. The potential of phytomelatonin as a nutraceutical. Molecules 2018, 23, 238. [Google Scholar] [CrossRef] [PubMed]
- Bonomini, F.; Borsani, E.; Favero, G.; Rodella, L.F.; Rezzani, R. Dietary melatonin supplementation could be a promising preventing/therapeutic approach for a variety of liver diseases. Nutrients 2018, 10, 1135. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Ma, Z.; Jiang, S.; Fan, C.; Deng, C.; Yan, X.; Di, S.; Lv, J.; Reiter, R.J.; Yang, Y. Melatonin: The dawning of a treatment for fibrosis? J. Pineal Res. 2016, 60, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Crespo, I.; San-Miguel, B.; Fernández, A.; Ortiz de Urbina, J.; González-Gallego, J.; Tuñón, M.J. Melatonin limits the expression of profibrogenic genes and ameliorates the progression of hepatic fibrosis in mice. Transl. Res. 2015, 165, 346–357. [Google Scholar] [CrossRef] [PubMed]
- San-Miguel, B.; Crespo, I.; Sánchez, D.I.; González-Fernández, B.; Ortiz de Urbina, J.J.; Tuñón, M.J.; González-Gallego, J. Melatonin inhibits autophagy and endoplasmic reticulum stress in mice with carbon tetrachloride-induced fibrosis. J. Pineal. Res. 2015, 59, 151–162. [Google Scholar] [CrossRef] [PubMed]
- González-Fernández, B.; Sánchez, D.I.; Crespo, I.; San-Miguel, B.; Álvarez, M.; Tuñón, M.J.; González-Gallego, J. Inhibition of the SphK1/S1P signaling pathway by melatonin in mice with liver fibrosis and human hepatic stellate cells. Biofactors 2017, 43, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Czechowska, G.; Celinski, K.; Korolczuk, A.; Wojcicka, G.; Dudka, J.; Bojarska, A.; Reiter, R.J. Protective effects of melatonin against thioacetamide-induced liver fibrosis in rats. J. Physiol. Pharmacol. 2015, 66, 567–579. [Google Scholar] [PubMed]
- Das, N.; Mandala, A.; Naaz, S.; Giri, S.; Jain, M.; Bandyopadhyay, D.; Reiter, R.J.; Roy, S.S. Melatonin protects against lipid-induced mitochondrial dysfunction in hepatocytes and inhibits stellate cell activation during hepatic fibrosis in mice. J. Pineal Res. 2017, 62, 12404. [Google Scholar] [CrossRef] [PubMed]
- Colares, J.R.; Schemitt, E.G.; Hartmann, R.M.; Licks, F.; Soares, M.D.; Bosco, A.D.; Marroni, N.P. Antioxidant and anti-inflammatory action of melatonin in an experimental model of secondary biliary cirrhosis induced by bile duct ligation. World J. Gastroenterol. 2016, 22, 8918–8928. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Meng, F.; Zhou, T.; Han, Y.; Kennedy, L.; Venter, J.; Francis, H.; DeMorrow, S.; Onori, P.; Invernizzi, P. Prolonged darkness reduces liver fibrosis in a mouse model of primary sclerosing cholangitis by miR-200b down-regulation. FASEB J. 2017, 31, 4305–4324. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Park, J.H.; Kim, K.; Jo, J.; Leem, J.; Park, K.K. Pharmacological inhibition of caspase-1 ameliorates cisplatin-induced nephrotoxicity through suppression of apoptosis, oxidative stress, and inflammation in mice. Mediat. Inflamm. 2018, 2018, 6571676. [Google Scholar] [CrossRef] [PubMed]
- Fabregat, I.; Moreno-Càceres, J.; Sánchez, A.; Dooley, S.; Dewidar, B.; Giannelli, G.; Ten Dijke, P.; IT-LIVER Consortium. TGF-β signalling and liver disease. FEBS J. 2016, 283, 2219–2232. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.E. Non-Smad signaling pathways of the TGF-β family. Cold Spring Harb. Perspect. Biol. 2017, 9, a022129. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.J.; Lin, Y.; Zhao, T.Z.; Zhang, C.K.; Zhang, T.; Chen, X.L.; Ding, J.Q.; Chang, T.; Zhang, Z.; Sun, C.; et al. Melatonin suppresses neuropathic pain via MT2-dependent and -independent pathways in dorsal root ganglia neurons of mice. Theranostics 2017, 7, 2015–2032. [Google Scholar] [CrossRef] [PubMed]
- Tamarindo, G.H.; Ribeiro, D.L.; Gobbo, M.G.; Guerra, L.H.A.; Rahal, P.; Taboga, S.R.; Gadelha, F.R.; Góes, R.M. Melatonin and docosahexaenoic acid decrease proliferation of PNT1A prostate benign cells via modulation of mitochondrial bioenergetics and ROS production. Oxid. Med. Cell. Longev. 2019, 2019, 5080798. [Google Scholar] [CrossRef] [PubMed]
- Pei, H.F.; Hou, J.N.; Wei, F.P.; Xue, Q.; Zhang, F.; Peng, C.F.; Yang, Y.; Tian, Y.; Feng, J.; Du, J.; et al. Melatonin attenuates postmyocardial infarction injury via increasing Tom70 expression. J. Pineal Res. 2017, 62, 12371. [Google Scholar] [CrossRef] [PubMed]
- Maarman, G.; Blackhurst, D.; Thienemann, F.; Blauwet, L.; Butrous, G.; Davies, N.; Sliwa, K.; Lecour, S. Melatonin as a preventive and curative therapy against pulmonary hypertension. J. Pineal Res. 2015, 59, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Yeung, H.M.; Hung, M.W.; Lau, C.F.; Fung, M.L. Cardioprotective effects of melatonin against myocardial injuries induced by chronic intermittent hypoxia in rats. J. Pineal Res. 2015, 58, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Sun, J.; Su, W.; Shan, H.; Zhang, B.; Wang, Y.; Shabanova, A.; Shan, H.; Liang, H. Melatonin protects against lung fibrosis by regulating the Hippo/YAP pathway. Int. J. Mol. Sci. 2018, 19, 1118. [Google Scholar] [CrossRef] [PubMed]
- Ogeturk, M.; Kus, I.; Kavakli, A.; Oner, J.; Kukner, A.; Sarsilmaz, M. Reduction of carbon tetrachloride-induced nephropathy by melatonin administration. Cell Biochem. Funct. 2005, 23, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Ozbek, E.; Ilbey, Y.O.; Ozbek, M.; Simsek, A.; Cekmen, M.; Somay, A. Melatonin attenuates unilateral ureteral obstruction-induced renal injury by reducing oxidative stress, iNOS, MAPK, and NF-kB expression. J. Endourol. 2009, 23, 1165–1173. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, N.; Yan, S.; Lu, Y.; Miao, X.; Gu, Z.; Shao, Y. Melatonin attenuates renal fibrosis in diabetic mice by activating the AMPK/PGC1α signaling pathway and rescuing mitochondrial function. Mol. Med. Rep. 2019, 19, 1318–1330. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.R.; Kim, K.H.; An, H.J.; Kim, J.Y.; Lee, S.J.; Han, S.M.; Pak, S.C.; Park, K.K. Apamin inhibits hepatic fibrosis through suppression of transforming growth factor β1-induced hepatocyte epithelial-mesenchymal transition. Biochem. Biophys. Res. Commun. 2014, 450, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Yoon, J.; Lee, K.Y.; Park, B. Effects of geniposide on hepatocytes undergoing epithelial-mesenchymal transition in hepatic fibrosis by targeting TGFβ/Smad and ERK-MAPK signaling pathways. Biochimie 2015, 113, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Yoon, J. Schizandrin inhibits fibrosis and epithelial-mesenchymal transition in transforming growth factor-β1-stimulated AML12 cells. Int. Immunopharmacol. 2015, 25, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Son, J.Y.; Kim, K.S.; Park, Y.J.; Kim, H.R.; Park, J.H.; Kim, K.B.; Lee, K.Y.; Kang, K.W.; Kim, I.S.; et al. Estrogen deficiency potentiates thioacetamide-induced hepatic fibrosis in Sprague-Dawley rats. Int. J. Mol. Sci. 2019, 20, 3709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Tang, X.; Yang, M.; Zhang, S.; Li, S.; Chen, Y.; Liu, M.; Guo, Y.; Lu, M. Interleukin 10 gene-modified bone marrow-derived dendritic cells attenuate liver fibrosis in mice by inducing regulatory T cells and inhibiting the TGF-β/Smad signaling pathway. Mediat. Inflamm. 2019, 2019, 4652596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.; Li, R.; Cai, M.; Huang, J.; Huang, W.; Guo, Y.; Yang, L.; Yang, G.; Lan, T.; Zhu, K. Andrographolide ameliorates liver fibrosis in mice: Involvement of TLR4/NF-κB and TGF-β1/Smad2 signaling pathways. Oxid. Med. Cell. Longev. 2018, 2018, 7808656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhao, H.; Li, H.; Cao, W.; Wang, F.; Zhang, T.; Wang, S.W. Protective effects of Amarogentin against carbon tetrachloride-induced liver fibrosis in mice. Molecules 2017, 22, 754. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Xie, F.; Zhang, Q.; Cui, Z.; Cheng, X.; Zhong, F.; He, K.; Zhou, J. Advanced oxidation protein products induce hepatocyte epithelial-mesenchymal transition via a ROS-dependent, TGF-β/Smad signaling pathway. Cell Biol. Int. 2017, 41, 842–853. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.L.; Huang, S.; Ma, X.X.; Zhang, W.Y.; Wang, D.; Jin, S.Y.; Zhang, Y.P.; Li, Y.; Li, X. Angiotensin(1-7) attenuated Angiotensin II-induced hepatocyte EMT by inhibiting NOX-derived H2O2-activated NLRP3 inflammasome/IL-1β/Smad circuit. Free Radic. Biol. Med. 2016, 97, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.H.; Tian, H.Y.; Gao, X.; Lei, W.W.; Hu, Y.; Wang, D.M.; Pan, X.C.; Yu, M.L.; Xu, G.J.; Zhao, F.K.; et al. Ferritin heavy chain-mediated iron homeostasis and subsequent increased reactive oxygen species production are essential for epithelial-mesenchymal transition. Cancer Res. 2009, 69, 5340–5348. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′→3′) | Product Size (bp) |
|---|---|---|
| E-cadherin | Forward: GACAGAAACGAGACTGGGTCA Reverse: CCGGTGATGCTGTAGAAAACC | 130 |
| α-SMA1 | Forward: GTCCCAGACATCAGGGAGTAA Reverse: TCGGATACTTCAGCGTCAGGA | 102 |
| Vimentin | Forward: GATCGATGTGGACGTTTCCAA Reverse: GTTGGCAGCCTCAGAGAGGT | 145 |
| Fibronectin | Forward: CGAGGTGACAGAGACCACAA Reverse: CTGGAGTCAAGCCAGACACA | 149 |
| GAPDH2 | Forward: ACTCCACTCACGGCAAATTC Reverse: TCTCCATGGTGGTGAAGACA | 171 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.-Y.; Park, J.-H.; Kim, K.; Leem, J.; Park, K.-K. Melatonin Inhibits Transforming Growth Factor-β1-Induced Epithelial–Mesenchymal Transition in AML12 Hepatocytes. Biology 2019, 8, 84. https://doi.org/10.3390/biology8040084
Kim J-Y, Park J-H, Kim K, Leem J, Park K-K. Melatonin Inhibits Transforming Growth Factor-β1-Induced Epithelial–Mesenchymal Transition in AML12 Hepatocytes. Biology. 2019; 8(4):84. https://doi.org/10.3390/biology8040084
Chicago/Turabian StyleKim, Jung-Yeon, Jae-Hyung Park, Kiryeong Kim, Jaechan Leem, and Kwan-Kyu Park. 2019. "Melatonin Inhibits Transforming Growth Factor-β1-Induced Epithelial–Mesenchymal Transition in AML12 Hepatocytes" Biology 8, no. 4: 84. https://doi.org/10.3390/biology8040084
APA StyleKim, J.-Y., Park, J.-H., Kim, K., Leem, J., & Park, K.-K. (2019). Melatonin Inhibits Transforming Growth Factor-β1-Induced Epithelial–Mesenchymal Transition in AML12 Hepatocytes. Biology, 8(4), 84. https://doi.org/10.3390/biology8040084