Structural Equation Modelling Reveals That Nutrients and Physicochemistry Act Additively on the Dynamics of a Microcosm-Based Biotic Community



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Sampling
2.2. Pairwise Correlations among Variables
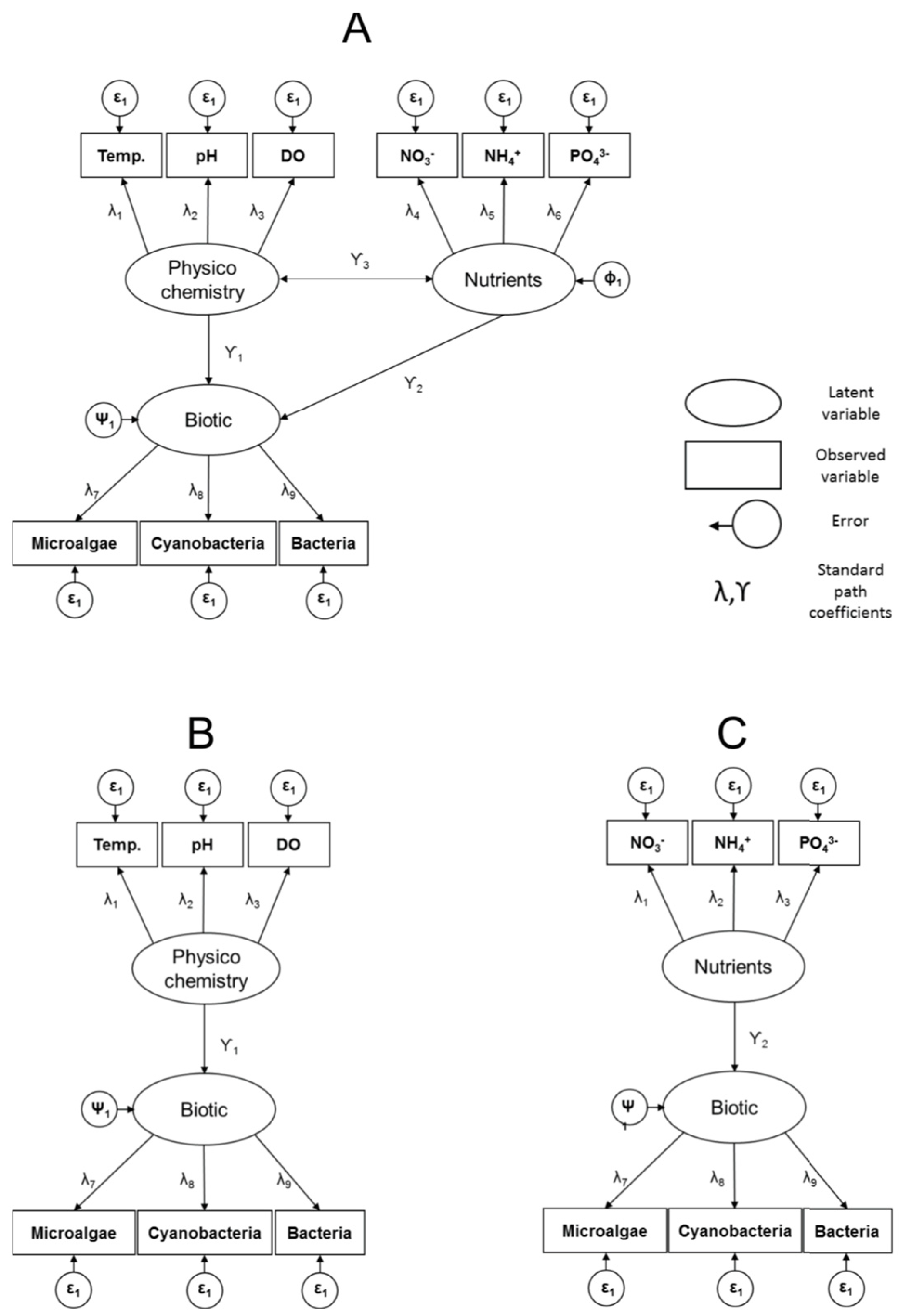
2.3. Quantifying Direct and Indirect Effects among Functional Biology, Nutrients, and Physicochemistry
3. Results
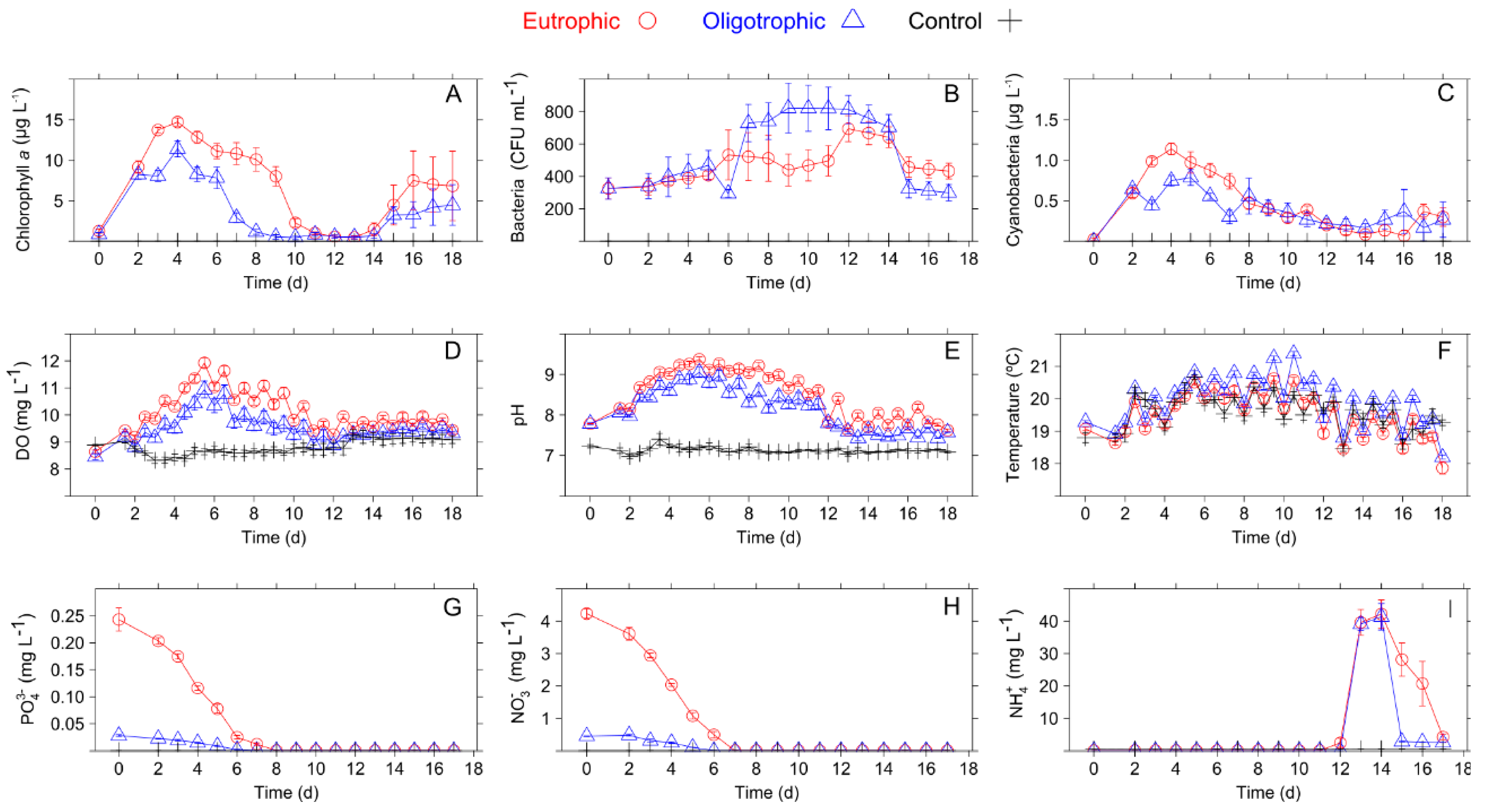
3.1. Pairwise Correlations among Variables
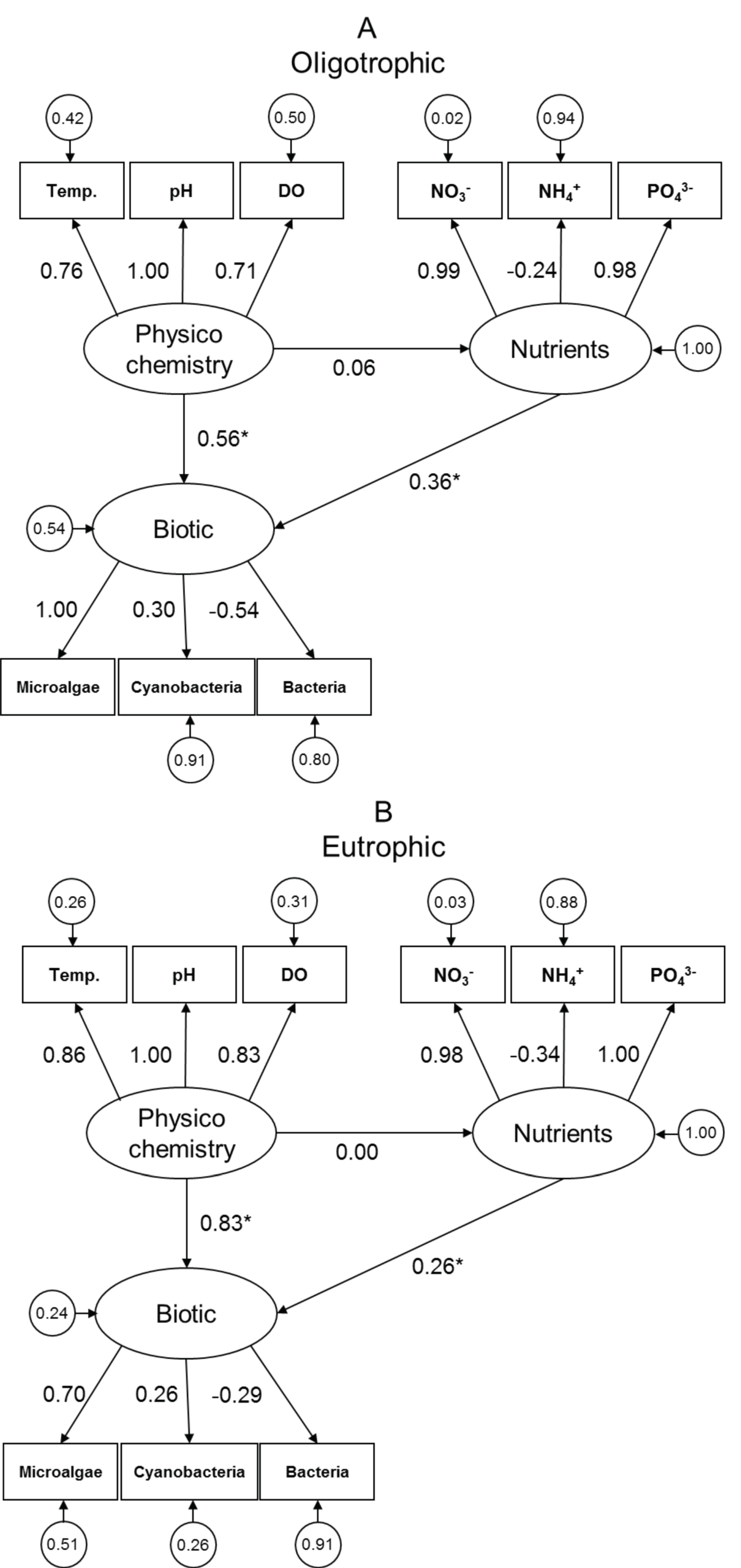
3.2. SEM Analysis: Quantifying Direct and Indirect Effects among Latent Variables Biotic, Nutrients, and Physicochemistry
4. Discussion
4.1. Comparison of Observed Univariate Relationships with Previous Studies
4.2. Utilising SEM to Quantify Direct and Indirect Effects among Functional Biology, Nutrients, and Physicochemistry
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| NaHCO3 | 192 |
| MnCl2·4H2O | 0.18 |
| MgSO47H2O | 115 |
| KCl | 0.45 |
| H2SeO3 | 0.0016 |
| Ca(NO3)24H2O | 0.8 |
| NH4Cl | 1 |
| KH2PO4 | 0.025 |
| K2PO4 | 0.025 |
| ZnSO4·7H2O | 0.022 |
| Na2EDTA.2H2O | 0.5 |
| H3BO3 | 0.114 |
| FeSO4·7H2O | 0.05 |
| CuSO4·5H2O | 0.016 |
| CoCl2·6H2O | 0.016 |
| (NH4)6Mo7O24·4H2O | 0.011 |

References
- Anderson, D.M.; Cembella, A.D.; Hallegraeff, G.M. Progress in Understanding Harmful Algal Blooms: Paradigm Shifts and New Technologies for Research, Monitoring, and Management. Annu. Rev. Mar. Sci. 2012, 4, 143–176. [Google Scholar] [CrossRef]
- Heisler, J.; Glibert, P.M.; Burkholder, J.M.; Anderson, D.M.; Cochlan, W.; Dennison, W.C.; Dortch, Q.; Gobler, C.J.; Heil, C.A.; Humphries, E.; et al. Eutrophication and harmful algal blooms: A scientific consensus. Harmful Algae 2008, 8, 3–13. [Google Scholar] [CrossRef]
- Smith, V.H.; Schindler, D.W. Eutrophication science: Where do we go from here? Trends Ecol. Evol. 2009, 24, 201–207. [Google Scholar] [CrossRef]
- Jöhnk, K.D.; Huisman, J.; Sharples, J.; Sommeijer, B.; Visser, P.M.; Stroom, J.M. Summer heatwaves promote blooms of harmful cyanobacteria. Glob. Chang. Biol. 2008, 14, 495–512. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Blooms Like It Hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef]
- Kosten, S.; Huszar, V.L.M.; Bécares, E.; Costa, L.S.; van Donk, E.; Hansson, L.-A.; Jeppesen, E.; Kruk, C.; Lacerot, G.; Mazzeo, N.; et al. Warmer climates boost cyanobacterial dominance in shallow lakes. Glob. Chang. Biol. 2012, 18, 118–126. [Google Scholar] [CrossRef]
- Muylaert, K.; Sabbe, K.; Vyverman, W. Spatial and Temporal Dynamics of Phytoplankton Communities in a Freshwater Tidal Estuary (Schelde, Belgium). Estuar. Coast. Shelf Sci. 2000, 50, 673–687. [Google Scholar] [CrossRef]
- Rigosi, A.; Carey, C.C.; Ibelings, B.W.; Brookes, J.D. The interaction between climate warming and eutrophication to promote cyanobacteria is dependent on trophic state and varies among taxa. Limnol. Oceanogr. 2014, 59, 99–114. [Google Scholar] [CrossRef]
- Bhattacharya, R.; Osburn, C.L. Multivariate Analyses of Phytoplankton Pigment Fluorescence from a Freshwater River Network. Environ. Sci. Technol. 2017, 51, 6683–6690. [Google Scholar] [CrossRef]
- Shipley, B. Cause and Correlation in Biology: A User’s Guide to Path Analysis, Structural Equations and Causal Inference with R, 2nd ed.; Cambridge University Press: Cambridge, UK, 2016; ISBN 978-1-107-44259-7. [Google Scholar]
- Lefcheck, J.S. piecewiseSEM: Piecewise structural equation modelling in r for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Stenegren, M.; Berg, C.; Padilla, C.C.; David, S.-S.; Montoya, J.P.; Yager, P.L.; Foster, R.A. Piecewise Structural Equation Model (SEM) Disentangles the Environmental Conditions Favoring Diatom Diazotroph Associations (DDAs) in the Western Tropical North Atlantic (WTNA). Front. Microbiol. 2017, 8, 810. [Google Scholar] [CrossRef]
- Kruk, M.; Kobos, J.; Nawrocka, L.; Parszuto, K. Positive and negative feedback loops in nutrient phytoplankton interactions related to climate dynamics factors in a shallow temperate estuary (Vistula Lagoon, southern Baltic). J. Mar. Syst. 2018, 180, 49–58. [Google Scholar] [CrossRef]
- Crossetti, L.O.; de Campos Bicudo, D.; Bini, L.M.; Dala-Corte, R.B.; Ferragut, C.; de Mattos Bicudo, C.E. Phytoplankton species interactions and invasion by Ceratium furcoides are influenced by extreme drought and water-hyacinth removal in a shallow tropical reservoir. Hydrobiologia 2019, 831, 71–85. [Google Scholar] [CrossRef]
- Russo, D.A.; Couto, N.; Beckerman, A.P.; Pandhal, J. A Metaproteomic Analysis of the Response of a Freshwater Microbial Community under Nutrient Enrichment. Front. Microbiol. 2016, 7, 1172. [Google Scholar] [CrossRef]
- International Organization for Standardization. Water Quality—Determination of Phosphorus—Ammonium Molybdate Spectrometric Method; ISO 6878:2004; ISO: Geneva, Switzerland, 2004. [Google Scholar]
- Salvesen, I.; Vadstein, O. Evaluation of plate count methods for determination of maximum specific growth rate in mixed microbial communities, and its possible application for diversity assessment. J. Appl. Microbiol. 2000, 88, 442–448. [Google Scholar] [CrossRef]
- Geissmann, Q. OpenCFU, a New Free and Open-Source Software to Count Cell Colonies and Other Circular Objects. PLoS ONE 2013, 8, e54072. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Rosseel, Y. lavaan: An R Package for Structural Equation Modeling. J. Stat. Softw. 2012, 48, 1–36. [Google Scholar] [CrossRef]
- Humphreys, R.K.; Puth, M.-T.; Neuhäuser, M.; Ruxton, G.D. Underestimation of Pearson’s product moment correlation statistic. Oecologia 2019, 189, 1–7. [Google Scholar] [CrossRef]
- Raven, J.A.; Gobler, C.J.; Hansen, P.J. Dynamic CO2 and pH levels in coastal, estuarine, and inland waters: Theoretical and observed effects on harmful algal blooms. Harmful Algae 2019, in press. [Google Scholar] [CrossRef]
- Paerl, H.W.; Hall, N.S.; Calandrino, E.S. Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic-induced change. Sci. Total Environ. 2011, 409, 1739–1745. [Google Scholar] [CrossRef]
- Elliott, J.A. Is the future blue-green? A review of the current model predictions of how climate change could affect pelagic freshwater cyanobacteria. Water Res. 2012, 46, 1364–1371. [Google Scholar] [CrossRef]
- Zhang, M.; Duan, H.; Shi, X.; Yu, Y.; Kong, F. Contributions of meteorology to the phenology of cyanobacterial blooms: Implications for future climate change. Water Res. 2012, 46, 442–452. [Google Scholar] [CrossRef]
- Tadonléke, R.D. Evidence of warming effects on phytoplankton productivity rates and their dependence on eutrophication status. Limnol. Oceanogr. 2010, 55, 973–982. [Google Scholar] [CrossRef]
- Domis, L.N.D.S.; de Waal, D.B.V.; Helmsing, N.R.; Donk, E.V.; Mooij, W.M. Community stoichiometry in a changing world: Combined effects of warming and eutrophication on phytoplankton dynamics. Ecology 2014, 95, 1485–1495. [Google Scholar] [CrossRef]
- Jankowiak, J.; Hattenrath-Lehmann, T.; Kramer, B.J.; Ladds, M.; Gobler, C.J. Deciphering the effects of nitrogen, phosphorus, and temperature on cyanobacterial bloom intensification, diversity, and toxicity in western Lake Erie. Limnol. Oceanogr. 2019, 64, 1347–1370. [Google Scholar] [CrossRef]
- Paerl, H.W. Controlling harmful cyanobacterial blooms in a climatically more extreme world: Management options and research needs. J. Plankton Res. 2017, 39, 763–771. [Google Scholar] [CrossRef]
- Lürling, M.; Mello, M.M.; van Oosterhout, F.; de Senerpont Domis, L.; Marinho, M.M. Response of Natural Cyanobacteria and Algae Assemblages to a Nutrient Pulse and Elevated Temperature. Front. Microbiol. 2018, 9, 1851. [Google Scholar] [CrossRef] [Green Version]


| Variable 1 | Variable 2 | Oligotrophic | Eutrophic |
|---|---|---|---|
| Bacteria | NO3− | −0.40 | −0.46 * |
| PO43− | −0.44 * | −0.45 * | |
| NH4+ | 0.22 | 0.36 | |
| Temp. | 0.27 | −0.03 | |
| DO | −0.08 | 0.00 | |
| pH | 0.02 | −0.15 | |
| Microalgae | NO3− | 0.38 | 0.20 |
| PO43− | 0.40 | 0.21 | |
| NH4+ | −0.31 | −0.37 | |
| Temp. | 0.17 | 0.29 | |
| DO | 0.56 * | 0.49 * | |
| pH | 0.58 * | 0.55 * | |
| Cyanobacteria | NO3− | 0.06 | 0.14 |
| PO43− | 0.06 | 0.17 | |
| NH4+ | −0.23 | −0.41 | |
| Temp. | 0.37 | 0.50 * | |
| DO | 0.33 | 0.65 * | |
| pH | 0.41 | 0.74 * | |
| Temperature | NO3− | 0.08 | 0.19 |
| PO43− | 0.07 | 0.16 | |
| NH4+ | −0.37 | −0.48 * | |
| DO | NO3− | −0.19 | −0.33 |
| PO43− | −0.13 | −0.31 | |
| NH4+ | −0.06 | −0.22 | |
| pH | NO3− | 0.04 | 0.03 |
| PO43− | 0.10 | 0.00 | |
| NH4+ | −0.36 | −0.57 * |
| Model Comparison | Condition | DF Difference | χ2 Difference | p-Value |
|---|---|---|---|---|
| A vs. B | Oligotrophic | 16 | 71.558 | <0.001 |
| Eutrophic | 16 | 73.305 | <0.001 | |
| A vs. C | Oligotrophic | 16 | 46.881 | <0.001 |
| Eutrophic | 17 | 76.334 | <0.001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Russo, D.A.; Ferguson, A.; Beckerman, A.P.; Pandhal, J. Structural Equation Modelling Reveals That Nutrients and Physicochemistry Act Additively on the Dynamics of a Microcosm-Based Biotic Community. Biology 2019, 8, 87. https://doi.org/10.3390/biology8040087
Russo DA, Ferguson A, Beckerman AP, Pandhal J. Structural Equation Modelling Reveals That Nutrients and Physicochemistry Act Additively on the Dynamics of a Microcosm-Based Biotic Community. Biology. 2019; 8(4):87. https://doi.org/10.3390/biology8040087
Chicago/Turabian StyleRusso, David A., Andrew Ferguson, Andrew P. Beckerman, and Jagroop Pandhal. 2019. "Structural Equation Modelling Reveals That Nutrients and Physicochemistry Act Additively on the Dynamics of a Microcosm-Based Biotic Community" Biology 8, no. 4: 87. https://doi.org/10.3390/biology8040087
APA StyleRusso, D. A., Ferguson, A., Beckerman, A. P., & Pandhal, J. (2019). Structural Equation Modelling Reveals That Nutrients and Physicochemistry Act Additively on the Dynamics of a Microcosm-Based Biotic Community. Biology, 8(4), 87. https://doi.org/10.3390/biology8040087