1. Introduction
The subclass Scuticociliatia (Protozoa, Ciliophora) includes several families of free-living ciliates that are abundant in marine habitats. Some scuticociliate species, including
P. dicentrarchi, have acquired the capacity to infect fish, causing high mortalities in some cultured fish species, including flatfish [
1,
2,
3,
4,
5]. However, there is no clear picture of what makes this scuticociliate a fish pathogen. It is generally thought that
P. dicentrarchi enters the fish through external lesions and then proliferates in the blood and in most internal organs where it causes important histopathological changes [
1,
3,
6,
7,
8,
9]. Although the mechanisms that
P. dicentrarchi uses to invade fish tissues are not known, it has been suggested that during infection the ciliate can release proteases, which are considered virulence factors [
10]. Ciliate proteases can destroy components of the turbot humoral immune response [
11,
12] and modify fish leukocyte functions [
13,
14], thus providing a mechanism for circumventing the fish immune system. In addition,
P. dicentrarchi has developed other mechanisms that appear to be important in relation to parasitism. For example, it possesses mitochondria with two respiratory pathways: the cytochrome pathway, which exists in all aerobic organisms, and an alternative, cyanide-insensitive oxidase pathway, which enables the ciliate to survive and proliferate under normoxic and hypoxic conditions [
15] and thus to become adapted to differences in oxygen levels in the host and seawater. In addition,
P. dicentrarchi has potent antioxidant defence mechanisms that may be important during infection, including several superoxide dismutases that help the ciliate resist the reactive oxygen species released by the host [
16]. Finally, it has been suggested that
P. dicentrarchi can release its extrusome contents to create a protective barrier against soluble factors of the host immune system [
17].
Several components of
P. dicentrarchi stimulate fish leucocytes, thereby increasing respiratory burst, degranulation and the expression of pro-inflammatory cytokines. Despite the high level of stimulation, the toxic substances produced by the fish leucocytes do not seem to be sufficient to kill the parasite [
11,
18]. However, a turbot NK-lysin (an effector molecule of cytotoxic lymphocytes) has been reported to have very high antiparasitic activity, thus directly affecting the viability of
P. dicentrarchi [
19]. The innate humoral and adaptive immune responses appear to be crucial in defending the fish against this parasite. In this respect, the complement system plays a key role in the fish defence against this pathogen, as it displays potent antiparasitic activity, especially when activated by the classical pathway [
11,
20]. In addition to complement, the fish coagulation system also plays an important role in immobilizing and killing the parasite [
21].
Little is known about how the fish immune system provides protection against parasites or even how it recognises the parasites (particularly ciliates) or which immune signalling pathways are stimulated. Pardo et al. [
22,
23] investigated how
P. dicentrarchi stimulates the immune system of fish, by analysing the changes in gene expression in organs of the turbot immune system (spleen, liver and kidney) during a
P. dicentrarchi infection. These researchers found that many genes regulating substances involved in the immune response, including chemokines, chemotaxins, complement, immunoglobulins, major histocompatibility complex, interferon, lectins, cytochrome P450 and lysozyme, were differentially expressed and that the response was strongest in the spleen at 1- and 3- days post-infection. Most transcriptomic studies of the immune response in fish infected with ciliates have been carried out with ectoparasites such as
Cryptocaryon irritans and
Ichthyophthirius multifiliis [
24,
25,
26,
27], usually with an infection time longer than 12 h. In addition, previous transcriptomic studies have analysed the response in either the parasite or in the cells of the fish immune system, but not in both. The scuticociliate
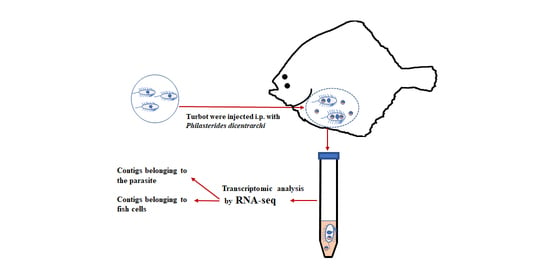
P. dicentrarchi can overcome the external barriers of fish and proliferate in the blood and in internal organs, causing a systemic infection. The initial interaction between the ciliate and fish immune system may be crucial to the success of infection. In the present study, we used transcriptomic technologies to analyse the interactions between
P. dicentrarchi and the turbot immune system during the very early stages of infection. Parasites were injected intraperitoneally and the transcriptomic response was evaluated in both fish cells and parasites found in the peritoneal cavity at several different times. This infection model enabled us to obtain sufficient numbers of host cells and parasites for the analysis.
4. Discussion
In the present study, we evaluated gene expression in the ciliate parasite
P. dicentrarchi at 1, 2 and 4 hpi, relative to the gene expression in samples processed at 0 h, during experimental infection in the turbot
S. maximus, to identify the genes participating in this process, including those potentially involved in virulence or in resistance to the host immune system. The most strongly regulated genes in
P. dicentrarchi during infection were the ABC transporters, which were mainly UR. Protozoan ABC transporters are involved in nutrient transport but they also protect cells from both internally produced and exogenous toxins, and some have been associated with antiparasitic drug resistance as well virulence and oxidative stress [
35,
36,
37]. These transporters are very abundant in ciliates, and 165 ABC transporter genes were identified in the macronuclear genome of the ciliate
Tetrahymena thermophila [
38]. In the present study, the
P. dicentrarchi ABC transporter G family genes showed a strong, early response during infection, indicating that some ABC transporters are very important in this process. It is not known whether they are involved in ciliate resistance against the attack of fish immune system or whether they have other roles in infection.
Very few free-living ciliate species can invade the fish, resist attack by the immune system and cause systemic infection. However,
P. dicentrarchi is capable of infecting cultured fish and causing massive mortalities. Thus, the ciliates must be able to resist the attack from the fish immune system and also to destroy fish tissue, for which proteases may be important. In the present study, we observed an increase in the expression of several genes of the leishmanolysin family, which are membrane-bound metalloproteases capable of degrading and cleaving many biological molecules [
39]. Leishmanolysin GP63 is a major surface protein and is considered one of the main virulence factors in the human pathogen
Leishmania [
40]. GP63 degrades a large number of proteins, including complement [
41], and down-regulation of gene expression makes the parasites more susceptible to complement-mediated lysis [
42]. The complement system is considered important in defence against
P. dicentrarchi [
11,
20], and although complement levels are lowered in the serum of infected turbot [
12], it is not known whether leishmanolysins participate in the process. Our results agree with those obtained in previous studies, which reported high level of leishmanolysin expression in a
P. dicentrarchi isolate (I1) obtained from turbot infections [
43], and UR of several leishmanolysin genes in
Miamiensis avidus fed a cell line [
44], suggesting that these molecules play a role in cell degradation. Thus, although the role of each regulated leishmanolysin gene in the success of
P. dicentrachi infection remains to be established, it appears that leishmanolysins are probably involved in ciliate virulence.
Ciliate proteases, other than leishmanolysins, are considered virulence factors in
P. dicentrarchi [
10,
12,
13,
14]. However, several members of the cathepsin family were DR during the early stages of
P. dicentrarchi infection. Interestingly, other genes whose proteins are also located in the lysosome were also DR. Because some ciliates contained phagocytosed turbot cells, release of lysosomal enzymes probably occurred during the early stages of infection. However, the expression of several cathepsin genes also did not increase in the scuticociliate
Anophryoides haemophila during an infection in American lobster [
45]. Although the expression of these genes will probably be enhanced at later stages in order to degrade the endocytosed material, the expression of these enzymes may be delayed during infection due to other priorities of the cells.
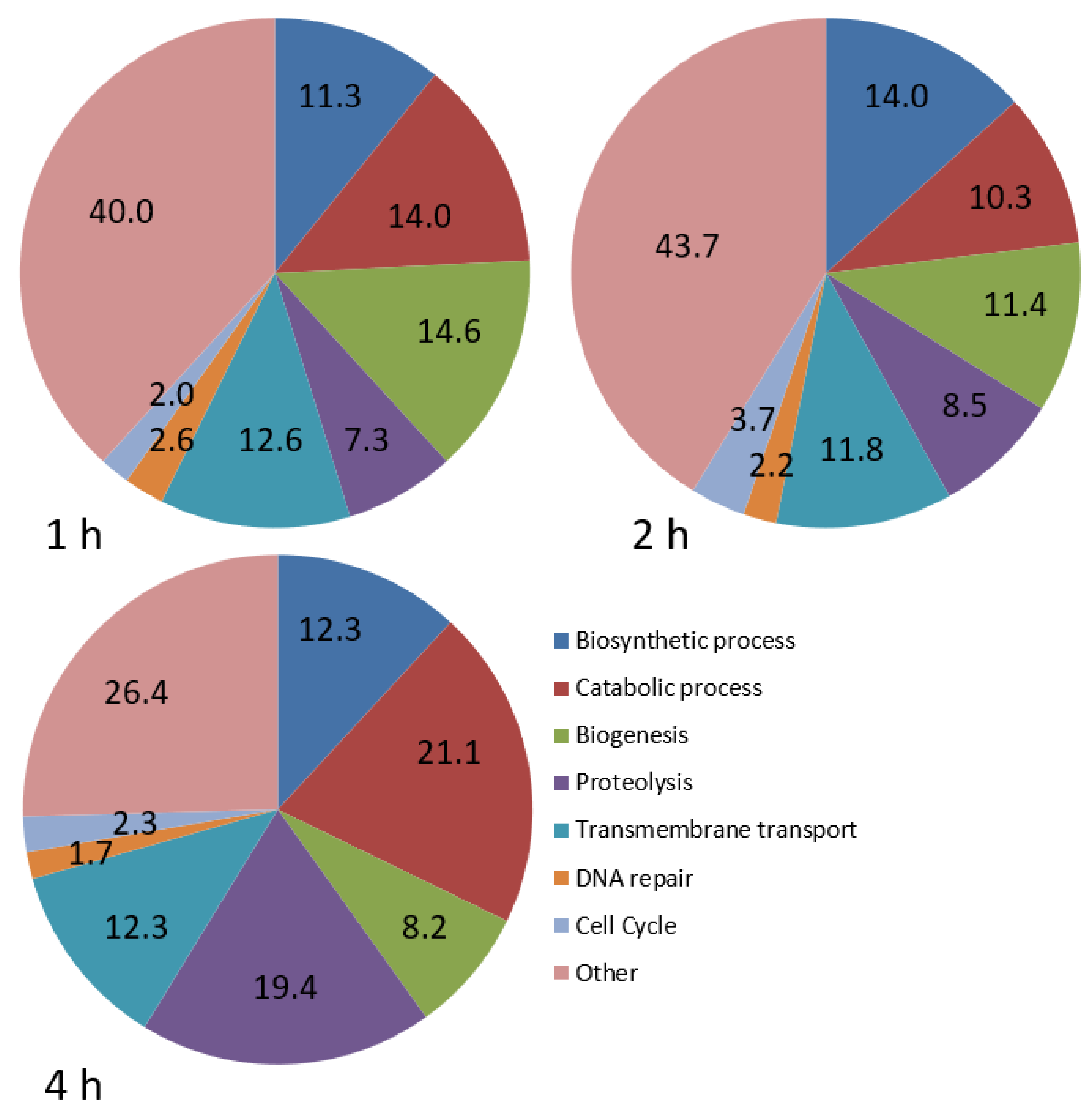
Many genes associated with rDNA transcription, ribosome biogenesis or rRNA synthesis were UR in
P. dicentrarchi during the early stages of infection, indicating a focus on ribosome production and protein synthesis in the ciliate. However, except for
kinesin family member 1, the
tubulin alpha and
beta genes and genes coding for several dyneins and kinesins were DR in
P. dicentrarchi during infection at 2 and/or 4 hpi. Tubulins and the associated motor proteins play many roles in eukaryotic cells, and particularly in ciliates, including cell division or cell movement. Genes involved in cell division, such as
cyclin-dependent kinase 1 (
cdk1) and
cyclin B, were also DR in
P. dicentrarchi during infection.
Cdk1 is a key protein that regulates mitotic entry and spindle assembly and its activation depends on
cyclin B [
46]. Our findings suggest that cell division is not activated during the early stages of infection. However, other genes associated with the cell cycle, such as
cdk2, were strongly UR.
Cdk2 activation leads to DNA replication in the cell, and it is also involved in DNA damage and the DNA repair response [
47]. Some ciliates probably suffered DNA damage because of attack from the turbot immune system, thus explaining the increase in
cdk2 expression. Several genes involved in glycolysis, the tricarboxylic acid (TCA) cycle and the mitochondrial respiratory chain were DR at 2 and (particularly) 4 hpi. If the
P. dicentrarchi mitochondrial genome is similar to that of
Tetrahymena pyriformis and
Paramecium aurelia [
48], the genes of the respiratory chain will be located in the mitochondrial genome, while those of the TCA cycle and glycolysis will be located in the nucleus. Unfortunately, little is known about how the expression of these genes is regulated in ciliates during infection. Many ciliate mitochondria may be altered because of interactions between the ciliate and the fish immune system. Previous studies have shown that turbot phagocytes are strongly stimulated by contact with
P. dicentrarchi, releasing a high amount of reactive oxygen species (ROS) [
11]. Leucocytes do not seem to produce sufficiently high concentrations of toxic substances to kill the parasite, but some ciliate components may be affected, as an increase in ROS levels is associated with alterations in the
P. dicentrachi mitochondria [
49]. Due to retrograde regulation, mitochondrial dysfunctions may affect the expression of enzymes involved in the TCA cycle, as observed in cells in other organisms [
50], thus explaining the DR of these genes.
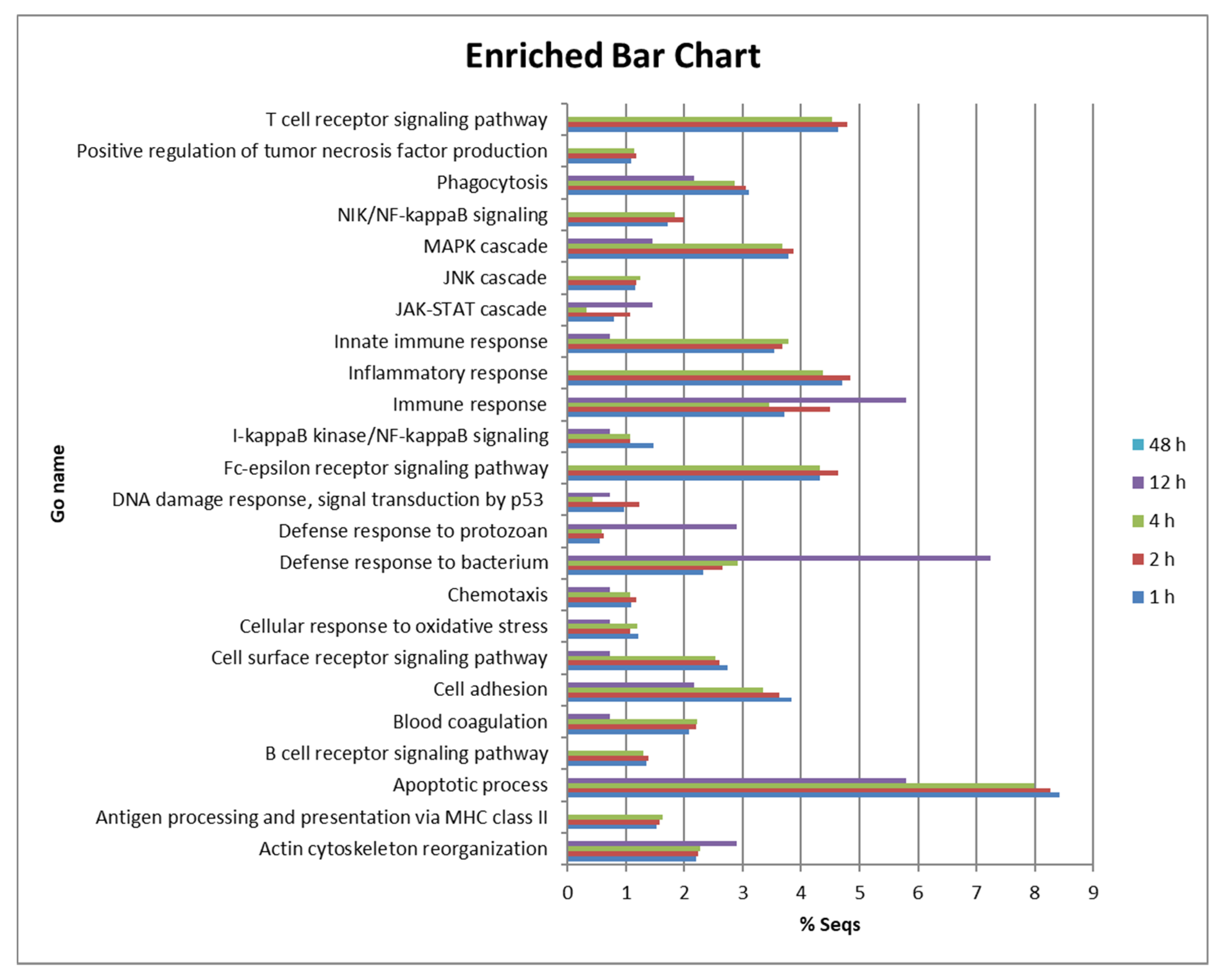
The present study shows that intraperitoneal injection of turbot with
P. dicentrarchi induced an acute inflammatory response and enhanced the expression of genes involved in inflammatory pathways, inflammatory cytokines or genes coding for enzymes involved in the synthesis of prostaglandins and leukotrienes. Recruitment of inflammatory cells, which is part of the inflammatory response [
51], was not evaluated in the present study but has been demonstrated in previous studies [
21]. Recognition of the pathogen by pattern recognition receptors is an important event in the inflammatory process. High levels of expression of
tlr5 and, to a lesser extent, of
tlr13 and
tlr2, occurred in turbot peritoneal cells although
tlr2 and
tlr13 expression began earlier.
Tlr2 expression increased in some organs of channel catfish (
Ictalurus punctatus) and orange-spotted grouper (
Epinephelus coioides) during infection with the ciliate ectoparasites
I. multifiliis and
C. irritans respectively [
24,
52,
53]. Studies in mammals have shown that glycosylphosphatidylinositol (GPI) is a potent activator of TLR2 in several protozoan parasites and that GPI is recognised by this receptor [
54,
55]. It has also been suggested that TLR2 mediates activation of the MAPK and NF-κB pathways [
56]. An increase in the expression of genes involved in these pathways was also observed in the present study, indicating that turbot TLR2 is activated during
P. dicentrarchi infection and that it may be important in the recognition of some ciliate molecules. However, because
P. dicentrarchi causes extensive damage in tissues and TLR2 can also be activated by several damage-associated molecular patterns [
57], an increase in the expression of this receptor associated with any of these molecules cannot be ruled out. Expression of
tlr13 and particularly
tlr5 was strongly enhanced in turbot peritoneal cells, peaking at 4 hpi and lasting until at least 12 hpi. TLR13 is located in endolysosomes and recognises bacterial 23s ribosomal RNA in mice, while TLR5 is located at the plasma membrane and recognises bacterial flagellin [
58]. Although TRL13 has been associated with the resistance of some insects to parasites [
59], it is not known whether it plays a role in fish parasite infections. Fish
tlr5 was also strongly up-regulated in some tissues of
E. coioides and in the Tibetan highland fish (
Gymnocypris przewalskii) in response to infection with the ciliates
C. irritans [
24,
60] and
I. multifiliis [
61] respectively, although, as far as we know, no particular role for this receptor in parasite recognition has been reported. The UR of
tlr5 found in the present study may be a consequence of prior stimulation of other cell receptors. In this respect, we cannot rule out unspecific stimulation of TLRs generated by a substance released by the parasite or even by a mechanical effect generated by the contact between the parasite membrane (which is moving all the time due to the movement of the cilia) and the leukocyte membranes.
Regarding the stimulation of inflammatory pathways, many components of NF-κB, MAPK and JAK/STAT pathways were UR. Some of those genes, such as
myd88, which was UR at 1, 2 and 4 hpi, have a positive effect on the pathways; however, others, such as
nfkbia, socs1 and
socs3, which were UR at 1, 2, 4 and 12 hpi, exert a negative effect in order to limit the inflammatory response [
62]. NF-κB, MAPK and JAK/STAT are involved in inflammatory processes, inducing expression of genes that regulate the inflammatory response, although they can also participate in other processes [
63,
64,
65]. Several chemokines and their receptors were UR in turbot peritoneal cells during
P. dicentrarchi infection. Among these, the strongest regulation was found in
cxcl8 and their receptors
cxcr1 and
cxcr2, which can be expressed by several types of leukocytes, especially neutrophils;
cxcl8 expression has been associated with neutrophil recruitment [
66]. This chemokine also induces chemotaxis of neutrophils and, to a lesser extent, lymphocytes and macrophages in turbot [
24,
67]. These results support those demonstrating intense migration of neutrophils to the turbot peritoneal cavity during
P. dicentrarchi infection [
21]. Several interleukins and their receptors, as well as several members of the TNF family and their receptors, were also strongly UR in turbot peritoneal cells, particularly the proinflammatory cytokines IL1b and TNFa. These and other inflammatory cytokines are produced by leukocytes, due to TLR stimulation, and promote inflammation [
58]. In addition, molecules such as TNFa can modulate multiple signalling pathways, some of which are related to the immune response during inflammation and also other responses, such as cell death [
68]. The role of both IL1b and TNFa in regulating the inflammatory response is well conserved in fish [
69]. Several other interleukins, interleukin receptors and members of the TNF superfamily that participate in the regulation of the inflammatory response are also strongly expressed in turbot peritoneal cells, including
il27b, il6ra, il6rb, il22r2, il3b2, il3b2, tnfaip8l2b, tnfaip2 and others, some of which, such as
tnfaip8l2b, are negative regulators of the immune response [
70], indicating the complexity of the response generated. Some of the interleukin genes such as
il10 and
il10r were UR at 12 hpi and at 1, 2, 4 and 12 hpi, respectively. IL10 displays potent anti-inflammatory and regulatory activity in most immune processes during infection [
71], suggesting an anti-inflammatory state in fish capable of controlling the infection. Increased expression of
il12b was observed at 12 and 48 hpi, although to a lower extent than for
il10. However, there was no increase in the expression of the complementary subunit or the receptors. In mammals, IL12B is secreted by phagocytic cells and acts on T and NK cells, inducing IFN-γ production and generating a proinflammatory state [
72]. An increase in IFN
-γ has been observed in leukocytes of other flatfish species stimulated with recombinant Il12 [
73]. However,
ifn-γ expression did not increase in turbot peritoneal leukocytes in the present study, suggesting that in this case, an increase in
il12b expression will not generate a proinflammatory state.
Several genes of the interferon family were UR, although to a much lesser extent than those related to other cytokines.
Interferon-induced double-stranded RNA-activated kinase was one of the genes in which DE was highest, being up-regulated at 1, 2, 4 and 12 hpi. This gene is a key component of host innate immunity that restricts viral replication and propagation but also participates in the stress response, being crucial for cell survival and proliferation, functions that are beyond the viral response [
74]. Most of the other DE genes were interferon regulatory factors (IRFs). Some of these factors are involved in TLR signalling pathways, inducing the expression of
type 1 ifn or proinflammatory cytokine genes [
75]. IRFs have been shown to play several roles in parasite infection, particularly
irf8, which appeared UR at 1, 2, 4 and 12 hpi in turbot peritoneal cells during
P. dicentrarchi infection and which regulates the production of proinflammatory cytokines during malarial infections [
76]. However, IRFs are also involved in differentiation and apoptosis of immune cells in mammals [
75] and in fish [
77]. All of these events seem to occur in the turbot peritoneal cavity, and different IRFs may participate in any of them.
Many genes associated with apoptosis were highly UR in turbot peritoneal cells during initial infection with
P. dicentrachi. Regulation of most genes occurred at 1, 2, 4 hpi, peaking at 4 hpi, as well as at 12 hpi in some. Previous studies have shown that
P. dicentrarchi proteases induce apoptosis in turbot cells [
14]. However, increased expression of genes related to apoptosis or in the number of apoptotic cells in turbot peritoneal cells after injection with other stimuli have also been observed [
31,
78]. Other studies have also shown that large numbers of neutrophils migrate to the peritoneal cavity after injection of
P. dicentrarchi [
21] and many of these cells seem to initiate the expression of genes leading to apoptosis. In later stages, these cells can be phagocytosed by macrophages, contributing to the resolution of inflammation [
79]. On the basis of these findings, it appears that most of the gene expression associated with apoptosis in turbot peritoneal cells occurs in neutrophils. However, it has been shown that other stimuli also cause apoptosis in fish peritoneal macrophages [
80], indicating that other cell types may also be involved in this process.
Increased expression of several T and B lymphocyte related genes was also observed in the turbot peritoneal cells. Thus, e.g.,
igm and
igd heavy chains,
cd79a and
cd22, which are expressed in B lymphocytes, were UR. This regulation may be related to the recruitment of B cells to the peritoneal cavity, as shown in other fish species after injection with different stimuli [
81]. Several genes related to cytotoxicity were also DE.
Granzyme A was UR at 1, 2 and 4 hpi and
granzyme b and
perforin-1 were UR only at 1 hpi. These genes are produced by cytotoxic lymphocytes in mammals and fish [
82,
83]. Information about how these molecules interact with parasites in fish is scarce. However, in mammals, it has been observed that in addition to killing parasite-infected cells, these cytotoxic granule effectors can kill parasites [
84]. It is not known whether these molecules can kill
P. dicentrarchi. However, a turbot NK-lysin, an effector molecule of cytotoxic T lymphocytes and NK-cells, has been shown to be toxic for
P. dicentrarchi, although infection did not clearly affect mRNA expression [
19]. On the basis of these observations, these molecules from cytotoxic cells appear highly likely to play a role in
P. dicentrarchi infection.
Finally, several genes related to the coagulation and the complement systems were also DE in turbot during
P. dicentrarchi infection. Several complement genes have been found to be expressed in immune organs of turbot during a
P. dicentrarchi infection [
23], as well as locally in
E. coioides infected with the ciliate
C. irritans [
24] and also in the skin or liver of common carp (
Cyprinus carpio) infected with
I. multifiliis [
85]. The findings of the present study are consistent with the aforementioned findings in suggesting that peritoneal cells express complement and that the expression is regulated during infection. Similar observations have been made for genes involved in coagulation. Both the complement and the coagulation systems play important roles in defending turbot against
P. dicentrarchi [
20,
21], and the complement or coagulation proteins released by leukocytes at the site of infection may represent an important additional source of these molecules.
How does the immune response generated by
P. dicentrarchi in turbot compare with that induced by other parasites in fish? The immune response induced in fish by different parasites varies widely. Thus, some parasites do not elicit a host response, while others induce a more or less intense inflammatory response [
86,
87]. Previous transcriptomic studies carried out in fish infected with ciliate parasites have shown that these parasites usually generate an inflammatory response. However, the sampling times were longer than used in the present study, and the responses are therefore not necessarily comparable. Transcriptomic analysis, involving a microarray enriched in immune genes, of spleen, kidney and liver of turbot infected with
P. dicentrarchi, revealed strong up-regulation of many immune response genes [
22,
23] at 1 dpi, but a less intense response than observed in turbot peritoneal cavity in the present study. Sampling times after experimental infection with ciliates were 12 h [
88], 24 h [
25] or 3 dpi [
24] in fish infected experimentally with
C. irritans. In the ectoparasite
I. multifiliis, sampling was conducted 2 dpi [
61] or 8 dpi [
27]. However, regardless of the differences between the sampling times in the present and previous studies, ectoparasite ciliates such as
C. irritants also induced a potent inflammatory response on the skin of infected fish, with up-regulation of genes involved in innate immunity, genes coding for TLRs, TLR signalling pathways, chemokines and chemokine receptors and genes associated with complement activation [
24]. Similarly,
I. multifiliis generated a massive immune response in the gills of rainbow trout (
Oncorhynchus mykiss) [
27].

 ,
, 



{kind=link}
{kind=link}
{kind=link}