Pathways of Photosynthesis in Non-Leaf Tissues



Abstract
Simple Summary
Abstract
1. Introduction
2. Sites of Non-Leaf Photosynthesis
2.1. Stems
2.2. Petioles
2.3. Flowers
2.4. Seeds
2.5. Fruits
2.6. Roots
3. Source of Carbon for Non-Leaf Photosynthesis
4. Dark Reactions of Photosynthesis in Non-Leaf Organs and Tissues
4.1. C3
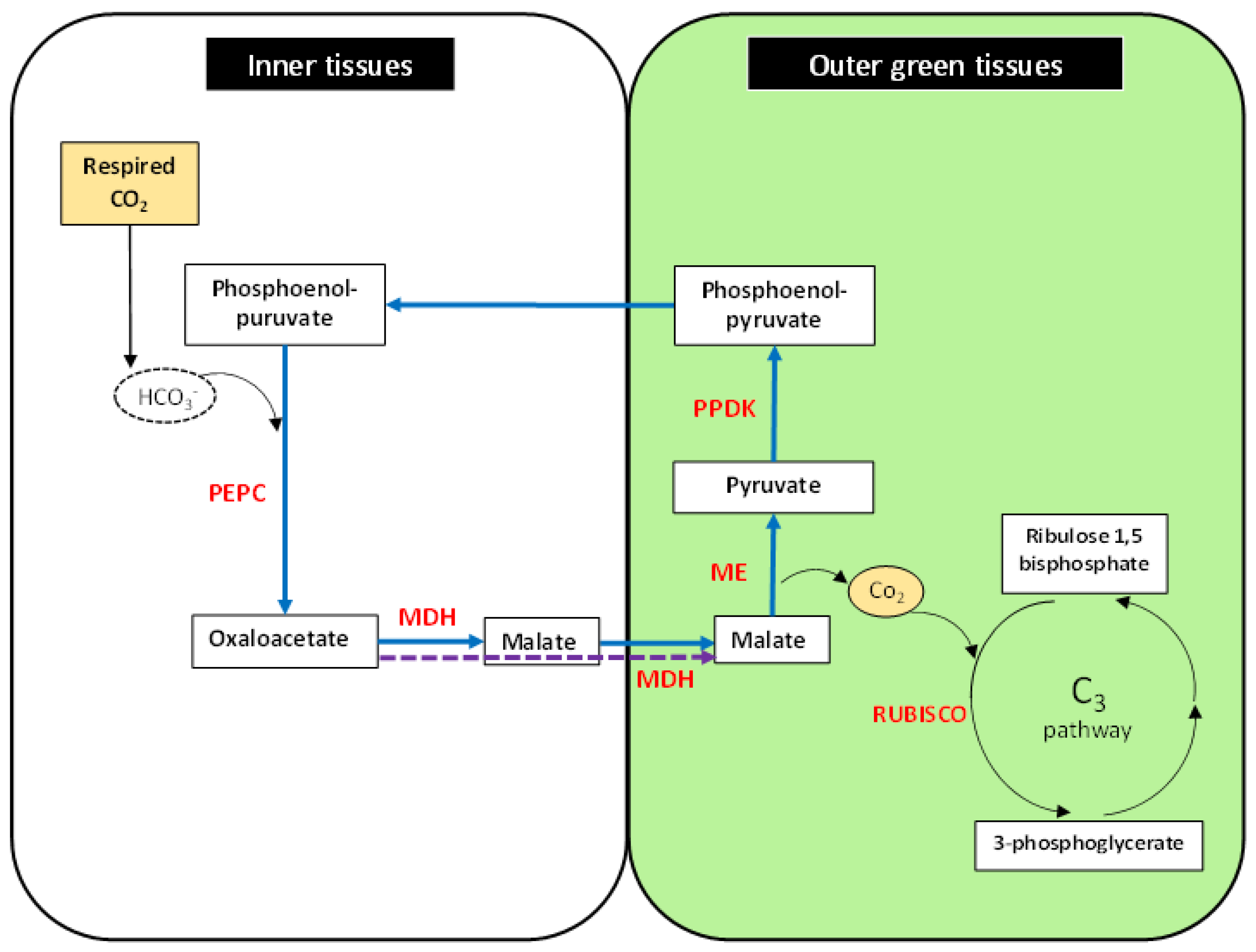
4.2. C4
4.3. CAM
4.4. Carbon Isotope Discrimination and Non-Leaf Photosynthesis
5. Importance and Function of Non-Leaf Photosynthesis
6. Evolutionary Considerations
7. Conclusions and Future Research Needs
Author Contributions
Funding
Conflicts of Interest
References
- Simkin, A.J.; Faralli, M.; Ramamoorthy, S.; Lawson, T. Photosynthesis in non-foliar tissues: Implications for yield. Plant J. 2020, 101, 1001–1015. [Google Scholar] [CrossRef]
- Jia, S.; Lv, J.; Jiang, S.; Liang, T.; Liu, C.; Jing, Z. Response of wheat ear photosynthesis and photosynthate carbon distribution to water deficit. Photosynthetica 2015, 53, 95–109. [Google Scholar] [CrossRef]
- Hibberd, J.M.; Quick, W.P. Characteristics of C-4 photosynthesis in stems and petioles of C-3 flowering plants. Nature 2002, 415, 451–454. [Google Scholar] [CrossRef]
- Werk, K.S.; Ehleringer, J.R. Photosynthesis by Flowers in Encelia-Farinosa and Encelia-Californica (Asteraceae). Oecologia 1983, 57, 311–315. [Google Scholar] [CrossRef]
- Smolikova, G.N.; Medvedev, S.S. Photosynthesis in the seeds of chloroembryophytes. Russ. J. Plant Physiol. 2016, 63, 1–12. [Google Scholar] [CrossRef]
- Eastmond, P.; Kolacna, L.; Rawsthorne, S. Photosynthesis by developing embryos of oilseed rape (Brassica napus L). J. Exp. Bot. 1996, 47, 1763–1769. [Google Scholar] [CrossRef]
- Sui, X.L.; Shan, N.; Hu, L.P.; Zhang, C.K.; Yu, C.Q.; Ren, H.Z.; Turgeon, R.; Zhang, Z.X. The complex character of photosynthesis in cucumber fruit. J. Exp. Bot. 2017, 68, 1625–1637. [Google Scholar] [CrossRef]
- Xu, H.L.; Gauthier, L.; Desjardins, Y.; Gosselin, A. Photosynthesis in leaves, fruits, stem and petioles of greenhouse-grown tomato plants. Photosynthetica 1997, 33, 113–123. [Google Scholar] [CrossRef]
- Anstis, P.J.P.; Northcote, D.H. Development of Chloroplasts from Amyloplasts in Potato Tuber Discs. New Phytol. 1973, 72, 449–464. [Google Scholar] [CrossRef]
- Henry, R.J.; Rangan, P.; Furtado, A. Functional cereals for production in new and variable climates. Curr. Opin. Plant Biol. 2016, 30, 11–18. [Google Scholar] [CrossRef]
- Henry, R. Innovations in Agriculture and Food Supply in Response to the COVID-19 Pandemic. Mol. Plant 2020, 13, 1095–1097. [Google Scholar] [CrossRef]
- Rangan, P.; Furtado, A.; Henry, R.J. New evidence for grain specific C-4 photosynthesis in wheat. Sci. Rep. 2016, 6, 31721. [Google Scholar] [CrossRef]
- Zieglerjons, A. Gas-Exchange of Ears of Cereals in Response to Carbon-Dioxide and Light.2. Occurrence of a C3-C4 Intermediate Type of Photosynthesis. Planta 1989, 178, 164–175. [Google Scholar] [CrossRef]
- Saveyn, A.; Steppe, K.; Ubierna, N.; Dawson, T.E. Woody tissue photosynthesis and its contribution to trunk growth and bud development in young plants. Plant Cell Environ. 2010, 33, 1949–1958. [Google Scholar] [CrossRef]
- Liu, J.X.; Gu, L.; Yu, Y.C.; Ju, G.S.; Sun, Z.Y. Stem Photosynthesis of Twig and Its Contribution to New Organ Development in Cutting Seedlings of Salix Matsudana Koidz. Forests 2018, 9, 207. [Google Scholar] [CrossRef]
- Badeck, F.W.; Tcherkez, G.; Nogues, S.; Piel, C.; Ghashghaie, J. Post-photo synthetic fractionation of stable carbon isotopes between plant organs—A widespread phenomenon. Rapid Commun. Mass Spectrom. 2005, 19, 1381–1391. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Condon, A.G.; Richards, R.A.; Rebetzke, G.J.; Farquhar, G.D. Breeding for high water-use efficiency. J. Exp. Bot. 2004, 55, 2447–2460. [Google Scholar] [CrossRef]
- Chen, X.; Gao, J.G.; Zhao, P.; McCarthy, H.R.; Zhu, L.W.; Ni, G.Y.; Ouyang, L. Tree Species with Photosynthetic Stems Have Greater Nighttime Sap Flux. Front. Plant Sci. 2018, 9, 30. [Google Scholar] [CrossRef]
- Wang, Q.; Lintunen, A.; Zhao, P.; Shen, W.J.; Salmon, Y.; Chen, X.; Ouyang, L.; Zhu, L.W.; Ni, G.Y.; Sun, D.; et al. Assessing Environmental Control of Sap Flux of Three Tree Species Plantations in Degraded Hilly Lands in South China. Forests 2020, 11, 206. [Google Scholar] [CrossRef]
- Rivera-Amado, C.; Trujillo-Negrellos, E.; Molero, G.; Reynolds, M.P.; Sylvester-Bradley, R.; Foulkes, M.J. Optimizing dry-matter partitioning for increased spike growth, grain number and harvest index in spring wheat. Field Crops Res. 2019, 240, 154–167. [Google Scholar] [CrossRef]
- AuBuchon-Elder, T.; Coneva, V.; Goad, D.M.; Jenkins, L.M.; Yu, Y.; Allen, D.K.; Kellogg, E.A. Sterile Spikelets Contribute to Yield in Sorghum and Related Grasses. Plant Cell 2020, 32, 3500–3518. [Google Scholar] [CrossRef]
- Wendling, I.; Brooks, P.R.; Trueman, S.J. Topophysis in Corymbia torelliana x C-citriodora seedlings: Adventitious rooting capacity, stem anatomy, and auxin and abscisic acid concentrations. New For. 2015, 46, 107–120. [Google Scholar] [CrossRef]
- Cowie, I.D.; Guymer, G.P. A new, rare species of Brachychiton from Fish River Station, Northern Territory. Aust. Syst. Bot. 2014, 27, 462–468. [Google Scholar] [CrossRef]
- Hoang, N.V.; Furtado, A.; Thirugnanasambandam, P.P.; Botha, F.C.; Henry, R.J. De novo assembly and characterizing of the culm-derived meta-transcriptome from the polyploid sugarcane genome based on coding transcripts. Heliyon 2018, 4, e00583. [Google Scholar] [CrossRef]
- Hibberd, J.M.; Furbank, R.T. Seeds of C-4 photosynthesis. Nat. Plants 2016, 2, 16172. [Google Scholar] [CrossRef]
- Leonardos, E.D.; Rauf, S.A.; Weraduwage, S.M.; Marilla, E.F.; Taylor, D.C.; Micallef-Bernard, B.J. Photosynthetic capacity of the inflorescence is a major contributor to daily-C-gain and the responsiveness of growth to elevated CO2 in Arabidopsis thaliana with repressed expression of mitochondrial-pyruvate-dehydrogenase-kinase. Environ. Exp. Bot. 2014, 107, 84–97. [Google Scholar] [CrossRef]
- Bazzaz, F.A.; Carlson, R.W. Photosynthetic Contribution of Flowers and Seeds to Reproductive Effort of an Annual Colonizer. New Phytol. 1979, 82, 223–232. [Google Scholar] [CrossRef]
- Lanoue, J.; Leonardos, E.D.; Grodzinski, B. Effects of Light Quality and Intensity on Diurnal Patterns and Rates of Photo-Assimilate Translocation and Transpiration in Tomato Leaves. Front. Plant Sci. 2018, 9, 756. [Google Scholar] [CrossRef]
- Lemoine, R.; La Camera, S.; Atanassova, R.; Dédaldéchamp, F.; Allario, T.; Pourtau, N.; Bonnemain, J.-L.; Laloi, M.; Coutos-Thévenot, P.; Maurousset, L.; et al. Source-to-sink transport of sugar and regulation by environmental factors. Front. Plant Sci. 2013, 4, 272. [Google Scholar] [CrossRef]
- Smolikova, G.; Kreslavski, V.; Shiroglazova, O.; Bilova, T.; Sharova, E.; Frolov, A.; Medvedev, S. Photochemical activity changes accompanying the embryogenesis of pea (Pisum sativum) with yellow and green cotyledons. Funct. Plant Biol. 2018, 45, 228–235. [Google Scholar] [CrossRef]
- Furbank, R.T.; White, R.; Palta, J.A.; Turner, N.C. Internal recycling of respiratory CO2 in pods of chickpea (Cicer arietinum L.): The role of pod wall, seed coat, and embryo. J. Exp. Bot. 2004, 55, 1687–1696. [Google Scholar] [CrossRef] [PubMed]
- Harris, M.; Mackender, R.O.; Smith, D.L. Photosynthesis of Cotyledons of Soybean Seedlings. New Phytol. 1986, 104, 319–329. [Google Scholar] [CrossRef]
- Meerow, A.W.; Snijman, D.A. Phylogeny of amaryllidaceae tribe amaryllideae based on nrDNA ITS sequences and morphology. Am. J. Bot. 2001, 88, 2321–2330. [Google Scholar] [CrossRef]
- Henry, R.J.; Rangan, P.; Furtado, A.; Busch, F.A.; Farquhar, G.D. Does C-4 Photosynthesis Occur in Wheat Seeds? Plant Physiol. 2017, 174, 1994–1995. [Google Scholar] [CrossRef]
- Cochrane, M.P.; Duffus, C.M. Morphology and Ultrastructure of Immature Cereal-Grains in Relation to Transport. Ann. Bot. 1979, 44, 67–72. [Google Scholar] [CrossRef]
- Zieglerjons, A. Gas-Exchange of Ears of Cereals in Response to Carbon-Dioxide and Light.1. Relative Contributions of Parts of the Ears of Wheat, Oat, and Barley to the Gas-Exchange of the Whole Organ. Planta 1989, 178, 84–91. [Google Scholar] [CrossRef]
- Blanke, M.M.; Lenz, F. Fruit photosynthesis. Plant Cell Environ. 1989, 12, 31–46. [Google Scholar] [CrossRef]
- Smille, R.M.; Hetherington, S.E.; Davies, W.J. Photosynthetic activity of the calyx, green shoulder, pericarp and locular parenchyma of tomato fruit. J. Exp. Bot. 1999, 50, 707–718. [Google Scholar] [CrossRef]
- Sma-Air, S.; Ritchie, R.J. Photosynthesis in a Vanda sp orchid with Photosynthetic Roots. J. Plant Physiol. 2020, 251, 153187. [Google Scholar] [CrossRef]
- Kocurek, M.; Kornas, A.; Wierzchnicki, R.; Luttge, U.; Miszalski, Z. Importance of stem photosynthesis in plant carbon allocation of Clusia minor. Trees Struct. Funct. 2020, 34, 1009–1020. [Google Scholar] [CrossRef]
- Hu, Y.Y.; Zhang, Y.L.; Luo, H.H.; Li, W.; Oguchi, R.; Fan, D.Y.; Chow, W.S.; Zhang, W.F. Important photosynthetic contribution from the non-foliar green organs in cotton at the late growth stage. Planta 2012, 235, 325–336. [Google Scholar] [CrossRef]
- Pyankov, V.I.; Voznesenskaya, E.V.; Kuz’min, A.N.; Ku, M.S.B.; Ganko, E.; Franceschi, V.R.; Black, C.C.; Edwards, G.E. Occurrence of C-3 and C-4 photosynthesis in cotyledons and leaves of Salsola species (Chenopodiaceae). Photosynth. Res. 2000, 63, 69–84. [Google Scholar] [CrossRef]
- Nutbeam, A.R.; Duffus, C.M. Evidence for C4 Photosynthesis in Barley Pericarp Tissue. Biochem. Biophys. Res. Commun. 1976, 70, 1198–1203. [Google Scholar] [CrossRef]
- Henry, R.J.; Furtado, A.; Rangan, P. Wheat seed transcriptome reveals genes controlling key traits for human preference and crop adaptation. Curr. Opin. Plant Biol. 2018, 45, 231–236. [Google Scholar] [CrossRef]
- Singal, H.R.; Sheoran, I.S.; Singh, R. Invitro Enzyme-Activities and Products of (Co2)-C-14 Assimilation in Flag Leaf and Ear Parts of Wheat (Triticum-Aestivum L). Photosynth. Res. 1986, 8, 113–122. [Google Scholar] [CrossRef]
- Imaizumi, N.; Samejima, M.; Ishihara, K. Characteristics of photosynthetic carbon metabolism of spikelets in rice. Photosynth. Res. 1997, 52, 75–82. [Google Scholar] [CrossRef]
- Wang, H.; Zhou, Q.P.; Mao, P.S. Ultrastructural and Photosynthetic Responses of Pod Walls in Alfalfa to Drought Stress. Int. J. Mol. Sci. 2020, 21, 4457. [Google Scholar] [CrossRef]
- Zhang, Y.; Mulpuri, S.; Liu, A.Z. Photosynthetic capacity of the capsule wall and its contribution to carbon fixation and seed yield in castor (Ricinus communis L.). Acta Physiol. Plant 2016, 38, 245. [Google Scholar] [CrossRef]
- Martin, C.E.; Mas, E.J.; Lu, C.; Ong, B.L. The photosynthetic pathway of the roots of twelve epiphytic orchids with CAM leaves. Photosynthetica 2010, 48, 42–50. [Google Scholar] [CrossRef]
- Ho, K.K.; Yeoh, H.H.; Hew, C.S. The Presence of Photosynthetic Machinery in Aerial Roots of Leafy Orchids. Plant Cell Physiol. 1983, 24, 1317–1321. [Google Scholar]
- Kriedemann, P. Photosynthetic Activity of Wheat Ear. Ann. Bot. 1966, 30, 349–363. [Google Scholar] [CrossRef]
- Dominguez, F.; Cejudo, F.J. Patterns of starchy endosperm acidification and protease gene expression in wheat grains following germination. Plant Physiol. 1999, 119, 81–87. [Google Scholar] [CrossRef]
- Hibberd, J.M.; Furbank, R.T. Fifty years of C-4 photosynthesis. Nature 2016, 538, 177–179. [Google Scholar] [CrossRef]
- Leegood, R.C. C4 photosynthesis: Principles of CO2 concentration and prospects for its introduction into C(3) plants. J. Exp. Bot. 2002, 53, 581–590. [Google Scholar] [CrossRef]
- Alvarez, H.M.; Hernndez, A. NADP(+)-dependent malic enzymes as a tool for improving oil production in Rhodococcus bacteria. New Biotechnol. 2018, 44, S89–S90. [Google Scholar] [CrossRef]
- Pedersen, O. Jack of all trades—C-4 photosynthesis, CAM and HCO3- use in the same tissue. A commentary on: ‘Structural basis for C-4 photosynthesis without Kranz anatomy in leaves of the submerged freshwater plant Ottelia alismoides’. Ann. Bot. 2020, 125, iv–vi. [Google Scholar] [CrossRef]
- Bort, J.; Brown, R.H.; Araus, J.L. Lack of C-4 Photosynthetic Metabolism in Ears of C-3 Cereals. Plant Cell Environ. 1995, 18, 697–702. [Google Scholar] [CrossRef]
- Rangan, P.; Furtado, A.; Henry, R.J. The transcriptome of the developing grain: A resource for understanding seed development and the molecular control of the functional and nutritional properties of wheat. BMC Genom. 2017, 18, 766. [Google Scholar] [CrossRef]
- Aoyagi, K.; Bassham, J.A.; Greene, F.C. Pyruvate Ortho-Phosphate Dikinase Gene-Expression in Developing Wheat Seeds. Plant Physiol. 1984, 75, 393–396. [Google Scholar] [CrossRef]
- Macnicol, P.K.; Jacobsen, J.V. Endosperm Acidification and Related Metabolic Changes in the Developing Barley-Grain. Plant Physiol. 1992, 98, 1098–1104. [Google Scholar] [CrossRef]
- Zhang, X.; Pu, P.; Tang, Y.; Zhang, L.X.; Lv, J.Y. C4 photosynthetic enzymes play a key role in wheat spike bracts primary carbon metabolism response under water deficit. Plant Physiol. Biochem. 2019, 142, 163–172. [Google Scholar] [CrossRef]
- Aoyagi, K.; Bassham, J.A. Appearance and Accumulation of C-4 Carbon Pathway Enzymes in Developing Wheat Leaves. Plant Physiol. 1986, 80, 334–340. [Google Scholar] [CrossRef]
- Harris, M.; Mackender, R.O.; Smith, D.L. Carboxylating Enzymes of Soybean Cotyledons. New Phytol. 1986, 104, 331–337. [Google Scholar] [CrossRef]
- Dawson, T.E.; Manbelli, S.; Plamboeck, A.H.; Templer, P.H.; Tu, K.P. Stable isotopes in plantecology. Annu. Rev. Ecol. Syst. 2002, 33, 507–559. [Google Scholar] [CrossRef]
- Hew, C.S.; Yong, J.W.H. The Physiology of Tropical Orchids in Relation to the Industry, 2nd ed.; World Scentific Press: Singapore, 2004; p. 388. [Google Scholar] [CrossRef]
- Hew, C.S.; Ng, C.K.Y.; Gouk, S.S.; Yong, J.W.H.; Wong, S.C. Variation in δ13C values for different plant parts of an Oncidium orchid. Photosynthetica 1996, 32, 135–139. [Google Scholar]
- Hu, Y.Y.; Zhang, Y.L.; Yi, X.P.; Zhan, D.X.; Luo, H.H.; Soon, C.W.; Zhang, W.F. The Relative Contribution of Non-Foliar Organs of Cotton to Yield and Related Physiological Characteristics Under Water Deficit. J. Integr. Agric. 2014, 13, 975–989. [Google Scholar] [CrossRef]
- Tsuchida, H.; Tamai, T.; Fukayama, H.; Agarie, S.; Nomura, M.; Onodera, H.; Ono, K.; Nishizawa, Y.; Lee, B.H.; Hirose, S.; et al. High level expression of C-4-specific NADP-malic enzyme in leaves and impairment of photoautotrophic growth in a C-3 plant, rice. Plant Cell Physiol. 2001, 42, 138–145. [Google Scholar] [CrossRef]
- Jansson, C.; Vogel, J.; Hazen, S.; Brutnell, T.; Mockler, T. Climate-smart crops with enhanced photosynthesis. J. Exp. Bot. 2018, 69, 3801–3809. [Google Scholar] [CrossRef]
- Dehigaspitiya, P.; Milham, P.; Ash, G.J.; Arun-Chinnappa, K.; Gamage, D.; Martin, A.; Nagasaka, S.; Seneweera, S. Exploring natural variation of photosynthesis in a site-specific manner: Evolution, progress, and prospects. Planta 2019, 250, 1033–1050. [Google Scholar] [CrossRef]
- Bräutigam, A.; Schlüter, U.; Eisenhut, M.; Gowik, U. On the Evolutionary Origin of CAM Photosynthesis. Plant Physiol. 2017, 174, 473–477. [Google Scholar] [CrossRef]
- Edwards, E.J. The distinct evolutionary trajectories of C4 and CAM photosynthesis. Integr. Comp. Biol. 2019, 59, E61. [Google Scholar]
- Sage, R.F.; Sage, T.L.; Kocacinar, F. Photorespiration and the Evolution of C4 Photosynthesis. Annu. Rev. Plant Biol. 2012, 63, 19–47. [Google Scholar] [CrossRef]
- Moreno-Villena, J.J.; Dunning, L.T.; Osborne, C.P.; Christin, P.A. Highly Expressed Genes Are Preferentially Co-Opted for C-4 Photosynthesis. Mol. Biol. Evol. 2018, 35, 94–106. [Google Scholar] [CrossRef]
- Silvera, K.; Santiago, L.S.; Winter, K. Distribution of crassulacean acid metabolism in orchids of Panama: Evidence of selection for weak and strong modes. Funct. Plant Biol. 2005, 32, 397–407. [Google Scholar] [CrossRef]
- Heyduk, K.; Moreno-Villena, J.J.; Gilman, I.S.; Christin, P.A.; Edwards, E.J. The genetics of convergent evolution: Insights from plant photosynthesis. Nat. Rev. Genet. 2019, 20, 485–493. [Google Scholar] [CrossRef]
- Edwards, G.E.; Franceschi, V.R.; Ku, M.S.B.; Voznesenskaya, E.V.; Pyankov, V.I.; Andreo, C.S. Compartmentation of photosynthesis in cells and tissues of C-4 plants. J. Exp. Bot. 2001, 52, 577–590. [Google Scholar] [CrossRef][Green Version]
- Voznesenskaya, E.V.; Franceschi, V.R.; Kiirats, O.; Freitag, H.; Edwards, G.E. Kranz anatomy is not essential for terrestrial C-4 plant photosynthesis. Nature 2001, 414, 543–546. [Google Scholar] [CrossRef]
- Sage, R.F. C-4 photosynthesis in terrestrial plants does not require Kranz anatomy. Trends Plant Sci. 2002, 7, 283–285. [Google Scholar] [CrossRef]
- Mao, Y.F.; Botella, J.R.; Liu, Y.G.; Zhu, J.K. Gene editing in plants: Progress and challenges. Natl. Sci. Rev. 2019, 6, 421–437. [Google Scholar] [CrossRef]
- Abberton, M.; Batley, J.; Bentley, A.; Bryant, J.; Cai, H.; Cockram, J.; Costa de Oliveira, A.; Cseke, L.J.; Dempewolf, H.; De Pace, C.; et al. Global agricultural intensification during climate change: A role for genomics. Plant Biotechnol. J. 2015, 14, 1095–1098. [Google Scholar] [CrossRef]
- Kole, C.; Muthamilarasan, M.; Henry, R.; Edwards, D.; Sharma, R.; Abberton, M.; Batley, J.; Bentley, A.; Blakeney, M.; Bryant, J.; et al. Application of genomics-assisted breeding for generation of climate resilient crops: Progress and prospects. Front. Plant Sci. 2015, 6, 563. [Google Scholar] [CrossRef]


{kind=link}
| Tissue | Species | Reference |
|---|---|---|
| Root | Potato tuber | [9] |
| Vanda (Orchidaceae) | [40] | |
| Oncidium | ||
| Chiloschista | ||
| Arachnis | ||
| Trunk/Stem | Tobacco | [3] |
| Prunis illicifolia | [13] | |
| Umbellularia californica | [13] | |
| Arctostaphylos amnzanita | [13] | |
| Salix matsudana | [14] | |
| Clusia minor (Clusiaceae) | [41] | |
| Cotton | [42] | |
| Petiole | Tobacco, Celery | [3] |
| Flower | Encelia spp (Asteraceae) | [4] |
| Giant ragweed (Ambrosia trifida) | [28] | |
| Salsola spp (Chenopodiaceae) | [43] | |
| Seed | Soybean | [15] |
| Barley (pericarp) | [44] | |
| Wheat | [12,45] | |
| Wheat (pericarp) | [46] | |
| Rice (ear) | [47] | |
| Alfalfa (pod walls) | [48] | |
| Castor (Rininus communis) | [49] | |
| Rape (Brassica napus) (embryo) | [6] | |
| Fruit | Cucumber | [7] |
| Tomato | [8] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henry, R.J.; Furtado, A.; Rangan, P. Pathways of Photosynthesis in Non-Leaf Tissues. Biology 2020, 9, 438. https://doi.org/10.3390/biology9120438
Henry RJ, Furtado A, Rangan P. Pathways of Photosynthesis in Non-Leaf Tissues. Biology. 2020; 9(12):438. https://doi.org/10.3390/biology9120438
Chicago/Turabian StyleHenry, Robert J., Agnelo Furtado, and Parimalan Rangan. 2020. "Pathways of Photosynthesis in Non-Leaf Tissues" Biology 9, no. 12: 438. https://doi.org/10.3390/biology9120438
APA StyleHenry, R. J., Furtado, A., & Rangan, P. (2020). Pathways of Photosynthesis in Non-Leaf Tissues. Biology, 9(12), 438. https://doi.org/10.3390/biology9120438