Comparative Ergot Alkaloid Elaboration by Selected Plectenchymatic Mycelia of Claviceps purpurea through Sequential Cycles of Axenic Culture and Plant Parasitism


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sources of Experimental Ergot
2.2. Cultivation of Axenic Mycelia
2.3. Analysis of Ergot Alkaloids
2.4. Cultivation of Host Plants
2.5. Analysis of Ergot Oils
3. Results
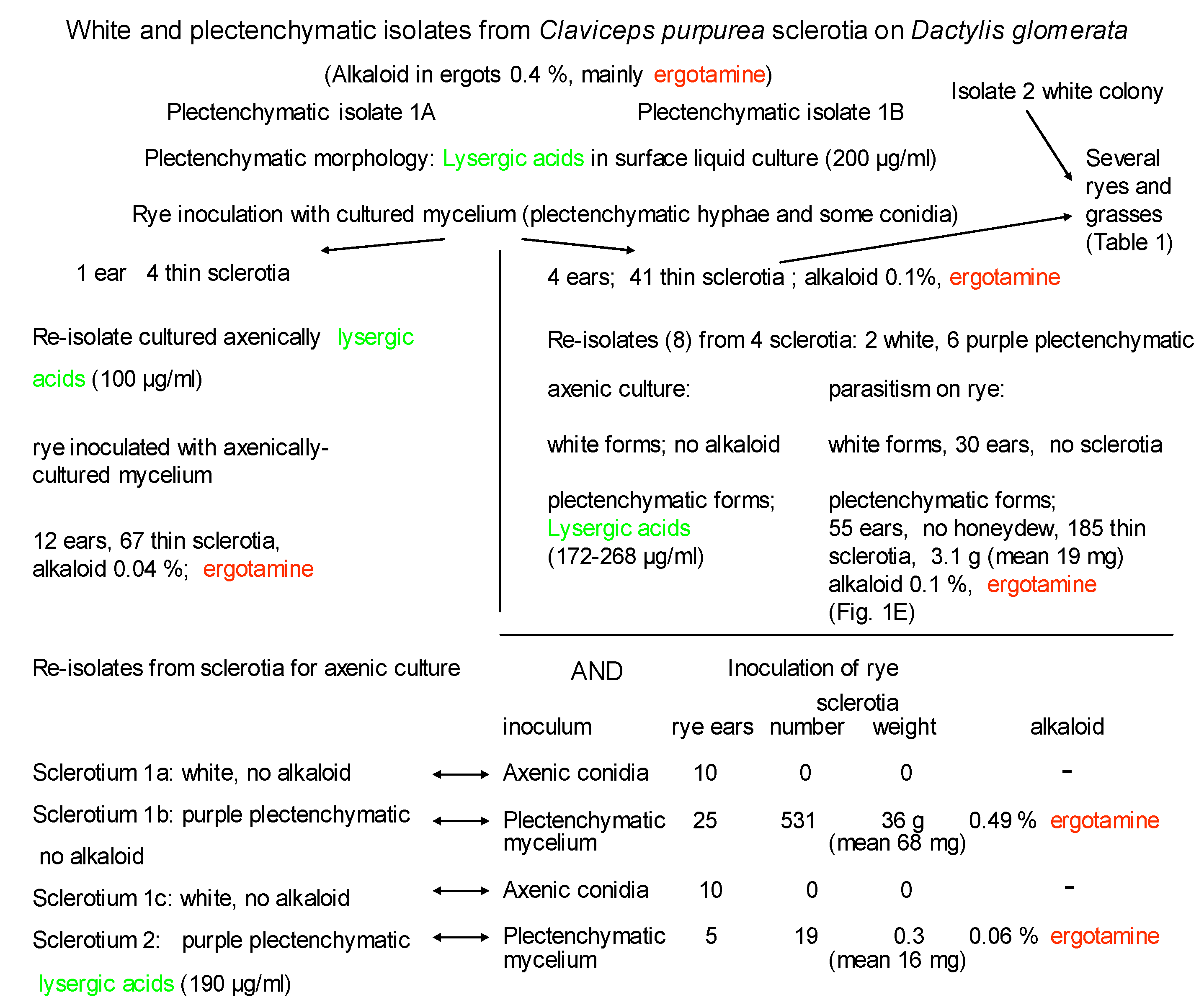
3.1. Ergot from Dactylis glomerata
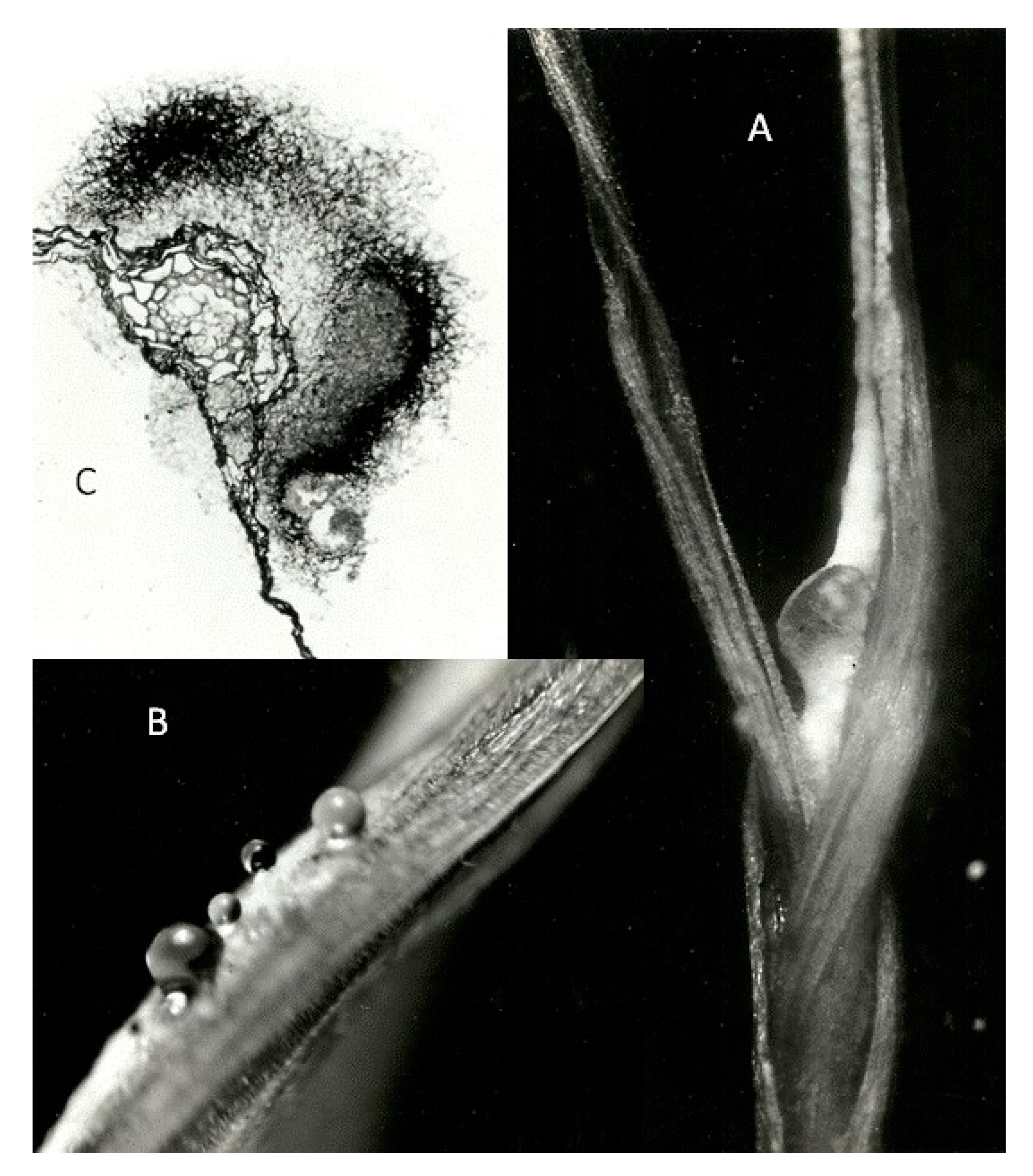
3.2. The Seedling Plumule Apex As a Putative Site for Ergot Parasitism
3.3. Ergot from Alopecurus myosuroides
4. Discussion
Funding
Conflicts of Interest
References
- Dunn, P. John Chassar Moir (1900–1977) and the discovery of ergometrine. Arch. Dis. Child Fetal Neonatal Ed. 2002, 87, F152–F154. [Google Scholar] [CrossRef]
- Arcamone, F.; Chain, E.B.; Ferretti, A.; Minghetti, A.; Pennella, P.; Tonolo, A.; Vero, L. Production of a new lysergic acid derivative in submerged culture by a strain of Claviceps paspali Stevens & Hall. Proc. Roy. Soc. Lond. B 1961, 155, 26–54. [Google Scholar]
- Tonolo, A.; Scotti, T.; Vero-Barcellona, L. Morphological observations on different species of Claviceps Tul. grown in submerged culture. Sci. Rep. Ist. Super. Sanita 1961, 1, 404–422. [Google Scholar]
- Mantle, P.G.; Tonolo, A. Relationship between the morphology of Claviceps purpurea and the production of alkaloids. Trans. Br. Mycol. Soc. 1968, 51, 499–505. [Google Scholar] [CrossRef]
- Tonolo, A. Production of peptide alkaloids in submerged culture by a strain of Claviceps purpurea (Fr.) Tul. Nat. Lond. 1966, 209, 1134. [Google Scholar] [CrossRef]
- Hofmann, A. Historical view on ergot. Pharmacology 1978, 16 (Suppl. 1), 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amici, A.M.; Minghetti, A.; Scotti, T.; Spalla, C.; Tognoli, L. Production of ergotamine by a strain of Claviceps purpurea (Fr.) Tul. Experientia 1966, 22, 415–416. [Google Scholar] [CrossRef] [PubMed]
- Mantle, P.G. Development of alkaloid production in vitro by a strain of Claviceps purpurea from Spartina townsendii. Trans. Br. Mycol. Soc. 1969, 62, 381–392. [Google Scholar] [CrossRef]
- Mantle, P.G. Studies on Claviceps purpurea (Fr.) Tul. parasitic on Phragmites communis Trin. Ann. Appl. Biol. 1969, 63, 425–434. [Google Scholar] [CrossRef]
- Mantle, P.G.; Morris, L.J.; Hall, S.W. Fatty acid composition of sphacelial and sclerotial growth forms of Claviceps purpurea in relation to the production of ergoline alkaloids in culture. Trans. Br. Mycol. Soc. 1969, 53, 441–447. [Google Scholar] [CrossRef]
- Mantle, P.G. Detection of ergot (Claviceps purpurea) in a dairy feed component by gas chromatography and mass spectrometry. J. Dairy Sci. 1996, 79, 1988–1991. [Google Scholar] [CrossRef]
- Amici, A.M.; Scotti, T.; Spalla, C.; Tognoli, L. Heterokaryosis and alkaloid production in Claviceps purpurea. Appl. Microbiol. 1967, 15, 611–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spalla, C. Genetic problems in production of ergot alkaloids in saprophytic andparasitic conditions. In Genetics of Industrial Microorganisms; Vanek, Z., Hostalek, Z., Cudlin, J., Eds.; Academia: Prague, Czech Republic, 1973; pp. 393–403. [Google Scholar]
- Esser, K.; Tudzynski, P. Genetics of the ergot fungus Claviceps purpurea. Theor. Appl. Genet. 1978, 53, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Miedaner, T.; Geiger, H.H. Biology, genetics, and management of ergot (Claviceps spp.) in rye, sorghum, and pearl millet. Toxins 2015, 7, 659–678. [Google Scholar] [CrossRef] [Green Version]
- Mantle, P.G.; Shaw, S.; Doling, D.A. Role of weed grasses in the aetiology of ergot disease in wheat. Ann. Appl. Biol. 1977, 86, 339–351. [Google Scholar] [CrossRef]
- Stoll, A.; Brack, A.; Hofmann, A.; Kobel, H. Process for the Preparation of Ergotamine, Ergotaminine and Ergometrine by Saprophytic Culture of Ergot (Claviceps purpurea [fr] tul.) In Vitro and Isolation of the Alkaloids Thus Produced. U.S. Patent 2,809,920, 15 October 1957. [Google Scholar]
- Gerhards, N.; Neubauer, L.; Tudzynski, P.; Shu-Ming, L. Biosynthetic pathways of ergot alkaloids. Toxins 2014, 6, 3281–3295. [Google Scholar] [CrossRef] [PubMed]
- Florea, S.; Panaccione, D.G.; Schardl, C.L. Ergot alkaloids of the family Clavicipitaceae. Phytopathology 2017, 107, 504–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castagnoli, N.; Mantle, P.G. Occurrence of D-lysergic acid and 6-methyl-ergol-8-ene-8-carboxylic acid in cultures of Claviceps purpurea. Nature 1966, 211, 859–860. [Google Scholar] [CrossRef]
- Abou-Chaar, C.I.; Brady, L.R.; Tyler, V.E. Occurrence of lysergic acid in cultures of Claviceps. Lloydia 1961, 24, 89–93. [Google Scholar]
- Mantle, P.G. Emergence and phytopathological properties of a new strain of Claviceps purpurea (Fr.) Tul. on rye. Ann. Appl. Biol. 1967, 60, 353–356. [Google Scholar] [CrossRef]
- Mantle, P.G. An unusual parasitic association between Claviceps purpurea and rye. Trans. Br. Mycol. Soc. 1972, 59, 327–330. [Google Scholar] [CrossRef]
- Pazoutova, S.; Pesikova, K.; Chudickova, M.; Srutka, P.; Kolarik, M. Delimitation of cryptic species inside Claviceps purpurea. Fun. Biol. 2015, 119, 7–26. [Google Scholar] [CrossRef]
- Roberts, A.; Beaumont, C.; Manzarpour, A.; Mantle, P. Purpurolic acid: A new natural alkaloid from Claviceps purpurea (Fr,) Tul. Fun. Biol. 2015, 120, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Corbett, K.; Dickerson, A.G.; Mantle, P.G. Metabolic studies on Claviceps purpurea during parasitic development on rye. J. Gen. Microbiol. 1974, 84, 39–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, B.I.; Mantle, P.G. Host infection by Claviceps purpurea. Trans. Br. Mycol. Soc. 1980, 75, 77–90. [Google Scholar] [CrossRef]
- Shaw, B.I.; Mantle, P.G. Parasitic differentiation of Claviceps purpurea. Trans. Br. Mycol. Soc. 1980, 75, 117–121. [Google Scholar] [CrossRef]
- Dickerson, A.G.; Mantle, P.G.; Nisbet, L.J.; Shaw, B.I. A role for β-glucanases in the parasitism of cereals by Claviceps purpurea. Physiol. Plant Path. 1978, 12, 55–62. [Google Scholar] [CrossRef]
- Oeser, B.; Kind, S.; Schurack, S.; Schmutzer, T.; Tudzynski, P.; Hinsch, J. Cross-talk of the biotrophic pathogen Claviceps purpurea and its host Secale cereale. BMC Genom. 2017, 18, 273. [Google Scholar] [CrossRef] [Green Version]
- Dickerson, A.G. A β-D-fructofuranosidase from Claviceps purpurea. Biochem. J. 1972, 129, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Woods, A.J.; Bradley Jones, J.; Mantle, P.G. An outbreak of gangrenous ergotism in cattle. Vet. Rec. 1966, 78, 742–749. [Google Scholar] [CrossRef]
- Mantle, P.G. Fatty acid composition of triglyceride oils from sphacelial, sclerotial and stromatal tissues of Claviceps purpurea and Claviceps sulcata. Trans. Br. Mycol. Soc. 1972, 59, 325–327. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host Plant | Inoculum | Infloresc-Ences Inoculated | Total Ergots | Ergot | Principal Alkaloid (% w/w) | |
|---|---|---|---|---|---|---|
| Weight (g) | Mean (mg) | |||||
| Fourex rye | PM | 48 | 310 Figure 2a | 26.35 | 85 | 0.34 ergotoxines |
| Winter rye | PM | 48 | 988 | 35.93 | 36 | 0.24 ergotoxines |
| Petkus rye | PM | 1 | 43 * Figure 2c | 1.2 | 28 | 0.23 ergotoxines |
| S. sylvestre | PM | 2 | 22 | 0.25 | 11 | 0.25 ergotoxines |
| S. africanum | PM | 4 | 17 | 0.42 | 25 | 0.28 ergotoxines |
| Triticale | PM | 1 | 19 Figure 2b | 1.2 | 63 | 0.30 ergotoxines |
| A. elatius | PM | 13 | 50 | 0.64 | 13 | 0.45 ergotoxines |
| D. glomerata | PM | 6 | 114 | 0.54 | 5 | 0.29 ergotoxines |
| Winter rye | CC | 18 | 138 * Figure 2d | 1.90 | 14 | 0.08 ergotamine |
| D. glomerata | CC | Many | 2500 | 10.25 | 4 | 0.08 ergotamine |
| 14:0 | 16:0 | 16:1 | 18:0 | 18:1 | 18:2 | 18:3 | OH-18:1 | |
|---|---|---|---|---|---|---|---|---|
| A. myosuroides isolate | ||||||||
| Parasitic sclerotia on rye | 0.3 | 27.8 | 4.2 | 2.7 | 20.5 | 7.0 | 0.7 | 36.8 |
| Axenic plectenchyma in submerged fermentation | 0.2 | 23.0 | 2.7 | 6.5 | 20.6 | 13.7 | 3.2 | 30.1 |
| Tonolo 1966 isolate | ||||||||
| Submerged. Alkaloid +ve | 0.1 | 13.2 | 5.2 | 1.3 | 25.1 | 30.9 | 0.7 | 23.5 |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mantle, P. Comparative Ergot Alkaloid Elaboration by Selected Plectenchymatic Mycelia of Claviceps purpurea through Sequential Cycles of Axenic Culture and Plant Parasitism. Biology 2020, 9, 41. https://doi.org/10.3390/biology9030041
Mantle P. Comparative Ergot Alkaloid Elaboration by Selected Plectenchymatic Mycelia of Claviceps purpurea through Sequential Cycles of Axenic Culture and Plant Parasitism. Biology. 2020; 9(3):41. https://doi.org/10.3390/biology9030041
Chicago/Turabian StyleMantle, Peter. 2020. "Comparative Ergot Alkaloid Elaboration by Selected Plectenchymatic Mycelia of Claviceps purpurea through Sequential Cycles of Axenic Culture and Plant Parasitism" Biology 9, no. 3: 41. https://doi.org/10.3390/biology9030041
APA StyleMantle, P. (2020). Comparative Ergot Alkaloid Elaboration by Selected Plectenchymatic Mycelia of Claviceps purpurea through Sequential Cycles of Axenic Culture and Plant Parasitism. Biology, 9(3), 41. https://doi.org/10.3390/biology9030041