


Anti-Melanogenic and Anti-Oxidative Effects of Nostoc verrucosum (ashitsuki) Extracts


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of N. verrucosum Extracts
2.1.1. Collection of N. verrucosum Colonies
2.1.2. Preparation of N. verrucosum HE
2.1.3. Preparation of N. verrucosum ME
2.1.4. Preparation of N. verrucosum AE
2.2. Cell Culture
2.3. Assay of 2,2’-Azino-bis (3-ethylbenzothiazoline-6-sulfonic Acid (ABTS) Radical Scavenging Activity
2.4. Assay of Tyrosinase Activity
2.5. Assay of Cell Proliferation
2.6. Determination of Melanin Content
2.7. DOPA Staining
2.8. Western Blotting
2.9. Quantitative Reverse-Transcription Polymerase Chain Reaction
2.10. Statistical Analysis
3. Results
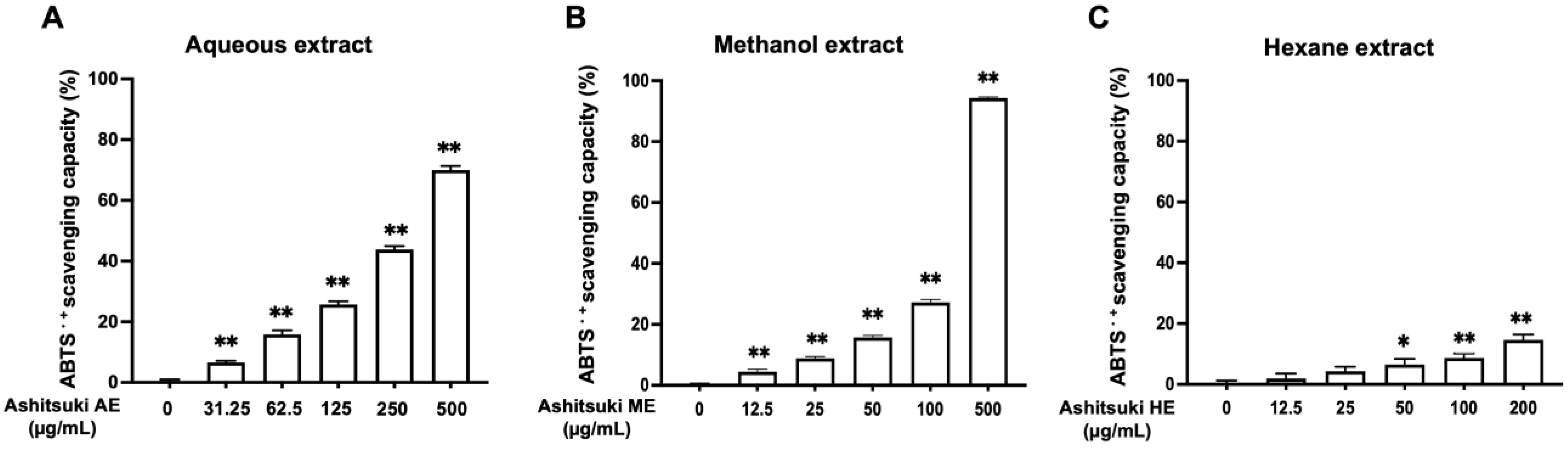
3.1. Antioxidant Activity
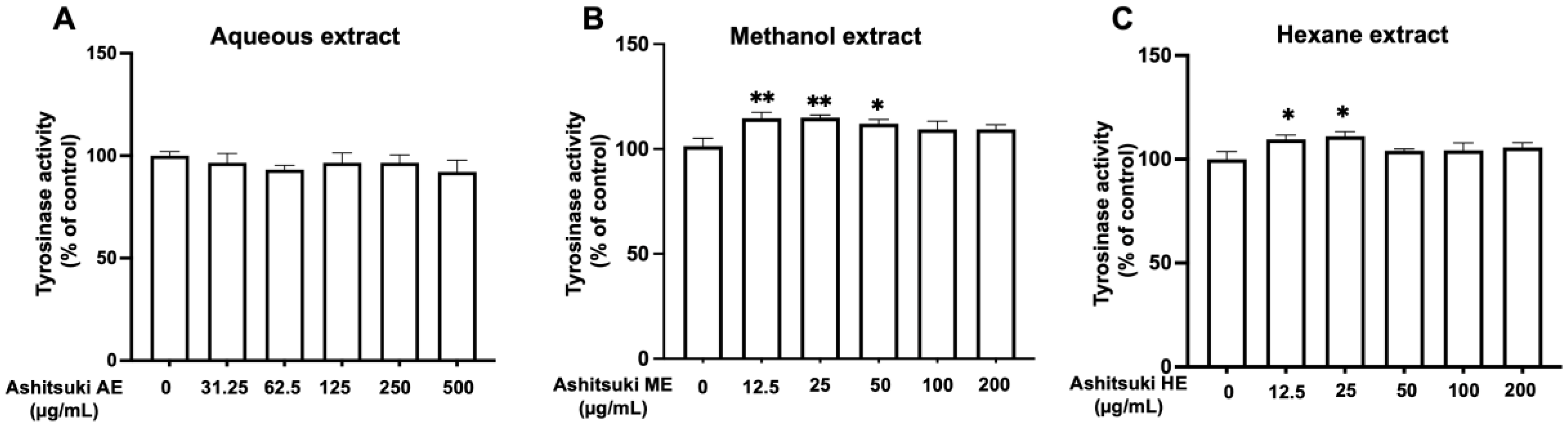
3.2. Mushroom Tyrosinase Activity
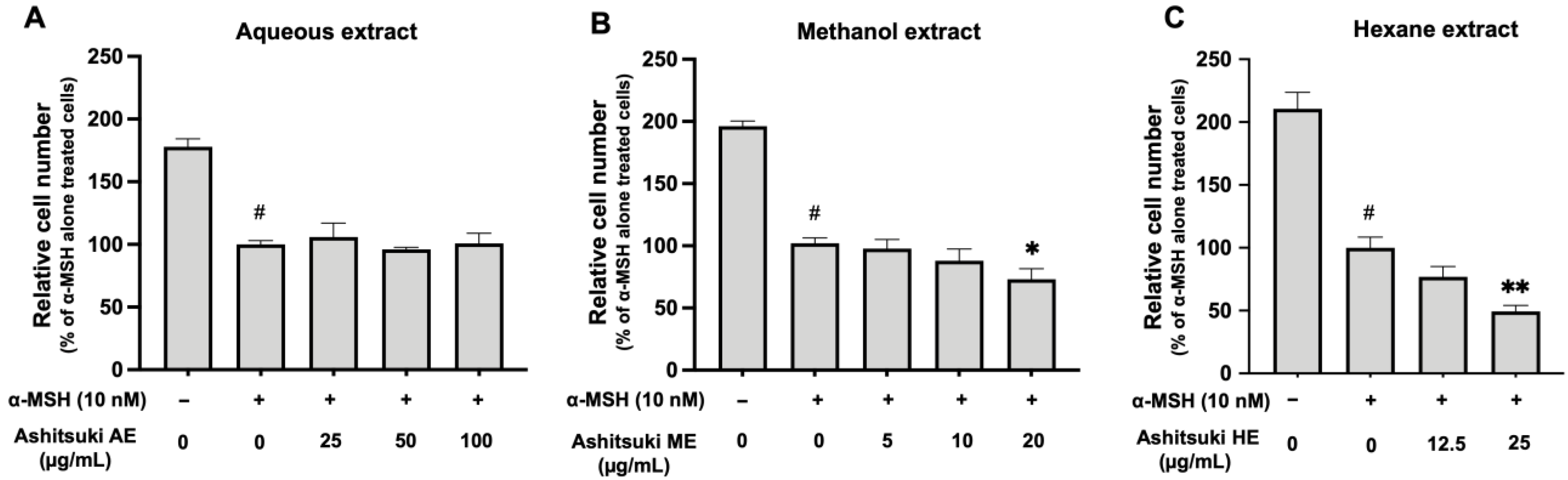
3.3. Cell Proliferation
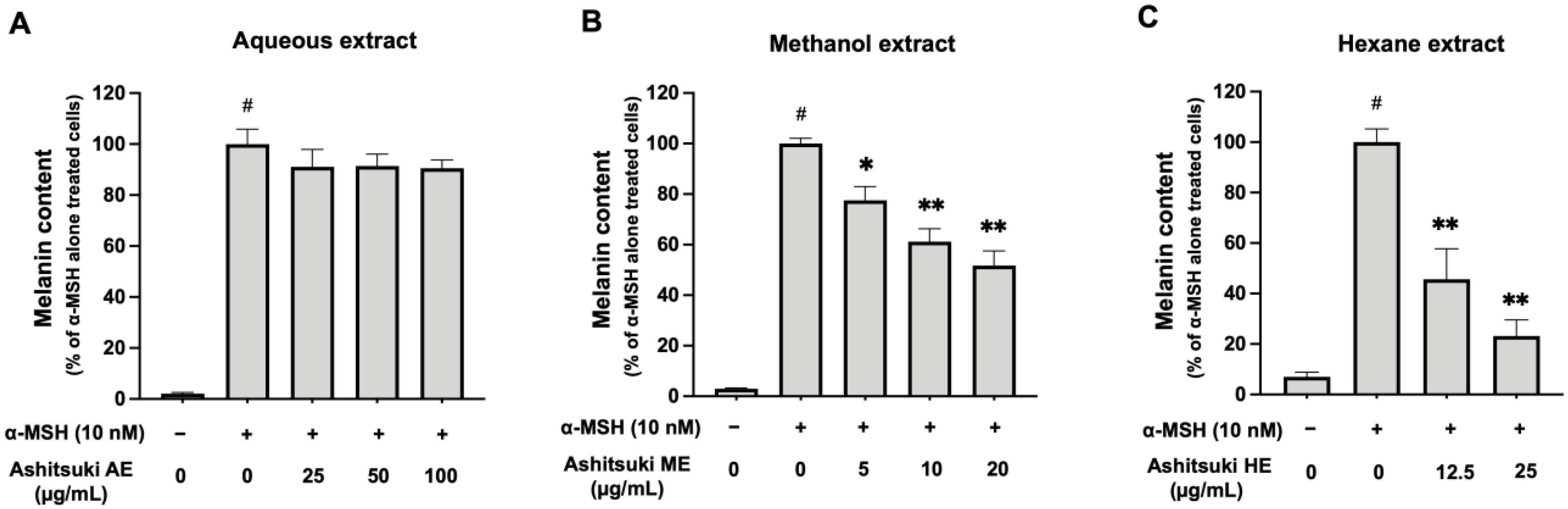
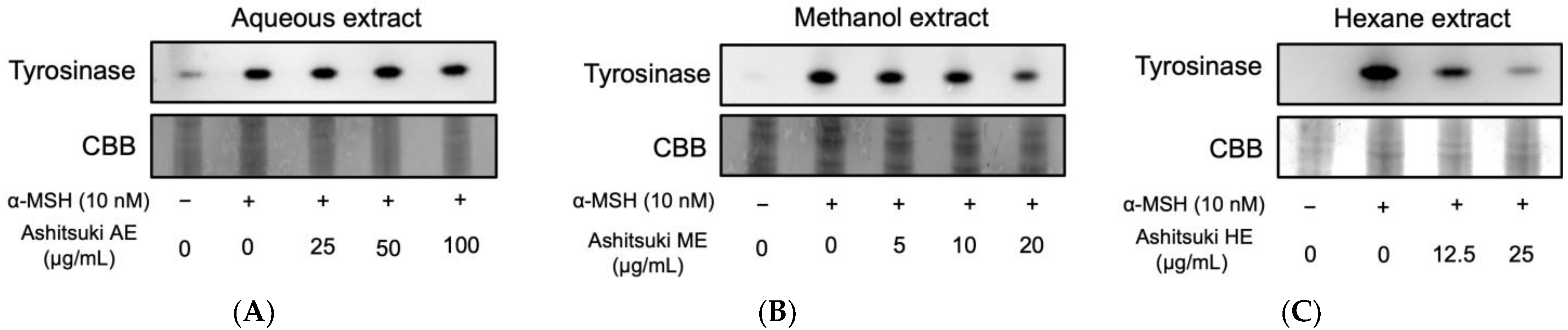
3.4. Melanin Production and Intracellular Tyrosinase Activity
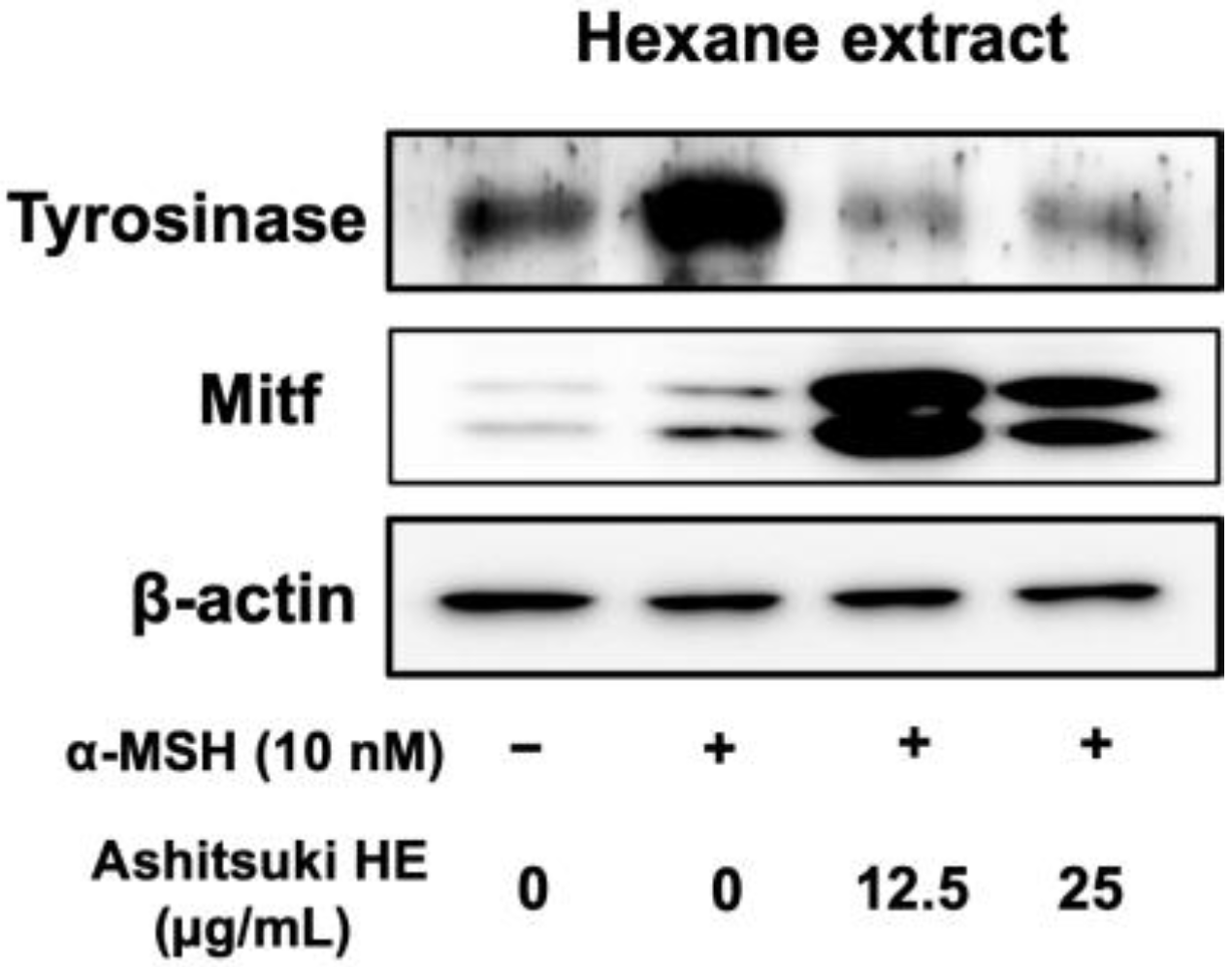
3.5. Effect of N. verrucosum HE on Tyrosinase and Mitf Protein Levels
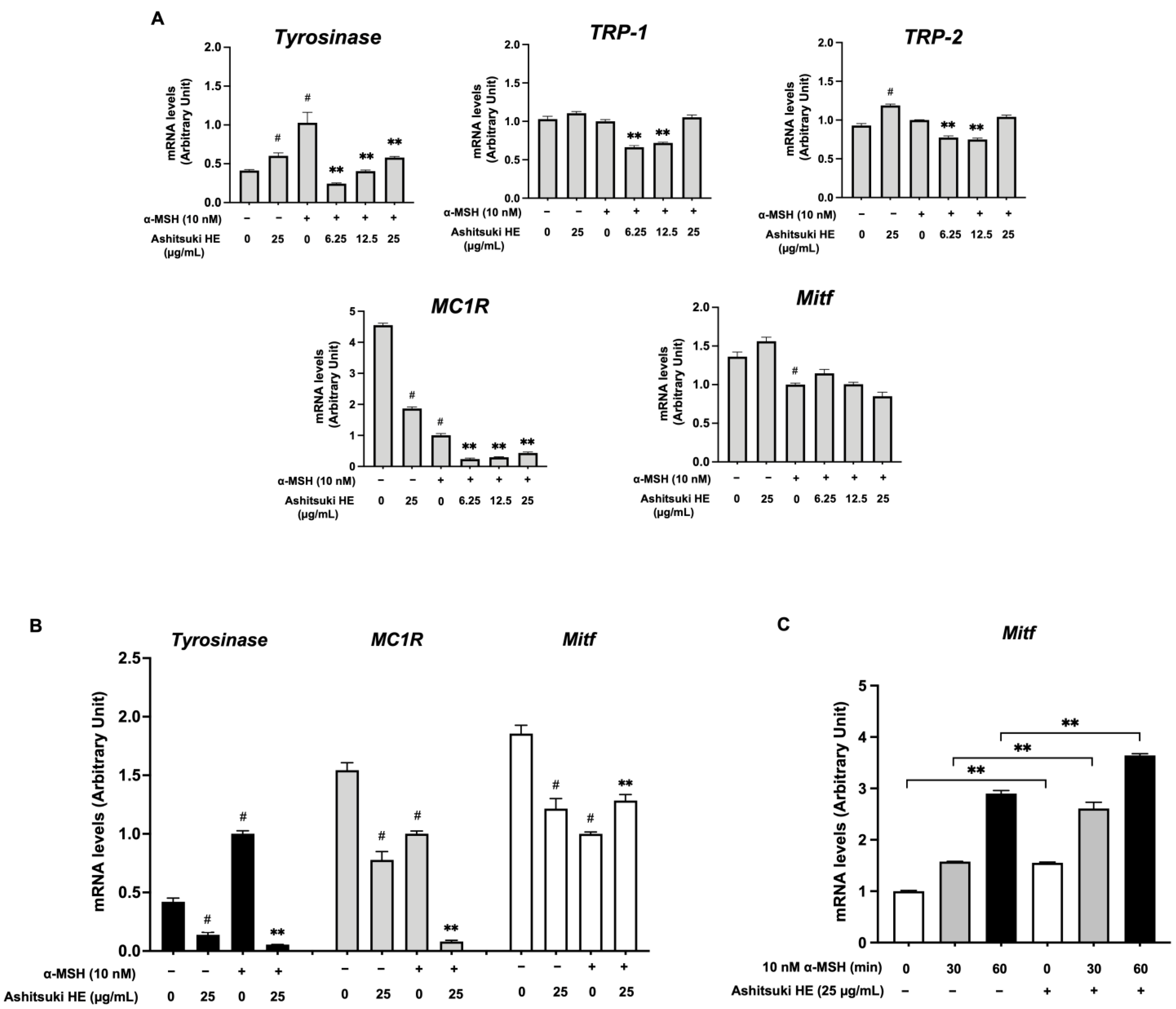
3.6. Influence of N. verrucosum HE on Melanogenic Genes Transcription
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oku, N.; Yonejima, K.; Sugawa, T.; Igarashi, Y. Identification of the n-1 fatty acid as an antibacterial constituent from the edible freshwater cyanobacterium Nostoc verrucosum. Biosci. Biotechnol. Biochem. 2014, 78, 1147–1150. [Google Scholar] [CrossRef] [PubMed]
- Johnson, H.E.; King, S.R.; Banack, S.A.; Webster, C.; Callanaupa, W.J.; Cox, P.A. Cyanobacteria (Nostoc commune) used as a dietary item in the Peruvian highlands produce the neurotoxic amino acid BMAA. J. Ethnopharmacol. 2008, 118, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Inoue-Sakamoto, K.; Nazifi, E.; Tsuji, C.; Asano, T.; Nishiuchi, T.; Matsuo, S.; Ishihara, K.; Kanesaki, Y.; Yoshikawa, H.; Sakamoto, T. Characterization of mycosporine-like amino acids in the cyanobacterium Nostoc verrucosum. J. Gen. Appl. Microbiol. 2018, 64, 201–211. [Google Scholar] [CrossRef]
- Korteerakul, C.; Honda, M.; Ngoennet, S.; Hibino, T.; Waditee-Sirisattha, R.; Kageyama, H. Antioxidative and antiglycative properties of mycosporine-like amino acids—Aontaining aqueous extracts derived from edible terrestrial cyanobacteria. J. Nutr. Sci. Vitaminol. 2020, 66, 339–346. [Google Scholar] [CrossRef]
- Maranduca, M.A.; Branisteanu, D.; Serban, D.N.; Branisteanu, D.C.; Stoleriu, G.; Manolache, N.; Serban, I.L. Synthesis and physiological implications of melanic pigments. Oncol. Lett. 2019, 17, 4183–4187. [Google Scholar] [CrossRef]
- Matoba, Y.; Kumagai, T.; Yamamoto, A.; Yoshitsu, H.; Sugiyama, M. Crystallographic evidence that the dinuclear copper center of tyrosinase is flexible during catalysis. J. Biol. Chem. 2006, 281, 8981–8990. [Google Scholar] [CrossRef]
- Tsukamoto, K.; Jackson, I.J.; Urabe, K.; Montague, P.M.; Hearing, V.J. A second tyrosinase-related protein, TRP-2, is a melanogenic enzyme termed DOPAchrome tautomerase. EMBO J. 1992, 11, 519–526. [Google Scholar] [CrossRef]
- Kameyama, K.; Takemura, T.; Hamada, Y.; Sakai, C.; Kondoh, S.; Nishiyama, S.; Urabe, K.; Hearing, V.J. Pigment production in murine melanoma cells is regulated by tyrosinase, tyrosinase-related protein 1 (TRP-1), DOPAchrome tautomerase (TRP2), and a melanogenic inhibitor. J. Investig. Dermatol. 1993, 100, 126–131. [Google Scholar] [CrossRef]
- Streingrimsson, E.; Copeland, N.G.; Jenkins, N.A. Melanocytes and the microphthalmia transcription factor network. Annu. Rev. Genet. 2004, 38, 365–411. [Google Scholar] [CrossRef]
- Sato, K.; Yamaguchi, Y.; Sakaki, S.; Takenaka, H. Pleurochrysis carterae hot-water extract inhibits melanogenesis in murine melanoma cells. Cosmetics 2019, 6, 60. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Ito, Y.; Sato, K. Salicylamide enhances melanin synthesis in B16F1 melanoma cells. Biomol. Ther. 2021, 29, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Takahashi, H.; Toriyama, M. Depigmenting mechanism of NSAIDs on B16F1 melanoma cells. Arch. Dermatol. Res. 2011, 303, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Takei, M.; Iyota, R.; Muraoka, Y.; Nagashijma, M.; Yoshimura, Y. Indomethacin inhibits melanogenesis via down-regulation of Mitf mRNA transcription. Biosci. Biotechnol. Biochem. 2017, 81, 2307–2313. [Google Scholar] [CrossRef] [PubMed]
- Khuanekkaphan, M.; Khobjai, W.; Noysang, C.; Wisidsri, N.; Thungmungmee, S. Bioactivities of Karanda (Carissa carandas Linn.) fruit extracts for novel cosmeceutical applicationsof Completion. J. Adv. Pharm. Technol. Res. 2021, 12, 162–168. [Google Scholar] [CrossRef]
- Worrapan Poomanee, W.; Khunkitti, W.; Chaiyana, W.; Intasai, N.; Lin, W.-C.; Lue, S.-L.; Leelapornpisid, P. Multifunctional biological properties and phytochemical constituents of Mangifera indica L. seed kernel extract for preventing skin aging. Toxicol. Res. 2021, 37, 459–472. [Google Scholar] [CrossRef]
- Shin, K.K.; Park, S.H.; Lim, H.Y.; Lorza, L.R.; Qomaladewia, N.P.; You, L.; Aziz, N.; Kim, S.A.; Lee, J.S.; Choung, E.S.; et al. In Vitro Anti-Photoaging and Skin Protective Effects of Licania macrocarpa Cuatrec Methanol Extract. Plants 2022, 11, 1383. [Google Scholar] [CrossRef]
- Swope, V.B.; Jameson, J.A.; McFarland, K.L.; Supp, D.M.; Miller, W.E.; McGraw, D.W.; Patel, M.A.; Nix, M.A.; Millhauser, G.L.; Babcock, G.F.; et al. Defining MC1R regulation in human melanocytes by its agonist α-melanocortin and antagonists agouti signaling protein and β-defensin 3. J. Investig. Dermatol. 2012, 132, 2255–2262. [Google Scholar] [CrossRef]
- Hasegawa, K.; Furuya, R.; Mizuno, H.; Umishio, K.; Suetsugu, M.; Sato, K. Inhibitory effect of Elephantopus mollis H.B. and K. extract on melanogenesis in B16 murine melanoma cells by downregulating microphthalmia-associated transcription factor expression. Biosci. Biotechnol. Biochem. 2010, 74, 1908–1912. [Google Scholar] [CrossRef]
- Seong, I.; Min, H.J.; Lee, J.-H.; Yeo, C.-Y.; Kang, D.M.; Oh, E.-S.; Hwang, E.S.; Kim, J. Sox10 controls migration of B16F10 melanoma cells through multiple regulatory target genes. PLoS ONE 2012, 7, e31477. [Google Scholar] [CrossRef]
- Park, J.; Jeong, D.; Jang, B.; Oh, E.-S. The melanocortin-1 receptor reversely regulates the melanin synthesis and migration of melanoma cells via dimerization-induced conformational changes. Biochem. Biophys. Res. Commun. 2019, 518, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Chuliá-Peris, L.; Carreres-Rey, C.; Gabasa, M.; Alcaraz, J.; Carretero, J.; Pereda, J. Matrix metalloproteinases and their inhibitors in pulmonary fibrosis: EMMPRIN/CD147 comes into play. Int. J. Mol. Sci. 2022, 23, 6894. [Google Scholar] [CrossRef] [PubMed]
- Parrado, C.; Mascaraque, M.; Gilaberte, Y.; Juarranz, A.; Gonzalez, S. Fernblock (Polypodium leucotomos Extract): Molecular mechanisms and pleiotropic effects in light-related skin conditions, photoaging and skin cancers, a review. Int. J. Mol. Sci. 2016, 17, 1026. [Google Scholar] [CrossRef]
- Zhou, J.; Wu, L.; Xu, P.; Li, Y.; Ji, Z.; Kang, X. Filamin A is a potential driver of breast cancer metastasis via regulation of MMP-1. Front. Oncol. 2022, 12, 836126. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, A.; Cheng, M.; Bhujwalla, Z.M.; Krishnamachary, B.; Jiang, L.; Glunde, K. Breast cancer cell adhesome and degradome interact to drive metastasis. NPJ Breast Cancer 2015, 1, 15017. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Kato, Y.; Erzinger, S.A.; Kiriakova, G.M.; Qian, Y.; Palmieri, D.; Steeg, P.S.; Price, J.E. The role of MMP-1 in breast cancer growth and metastasis to the brain in a xenograft model. BMC Cancer 2012, 12, 583. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Kim, J.Y.; Heo, J.; Cho, D.-H.; Kim, H.-S.; An, I.-S.; An, S.; Bae, S. The inhibition of melanogenesis via the PKA and ERK signaling pathways by Chlamydomonas reinhardtii extract in B16F1 melanoma cells and artificial human skin equivalents. J. Microbiol. Biotechnol. 2018, 28, 2121–2132. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kwon, Y.M.; Kim, K.W.; Kim, J.Y.H. Exploring the potential of Nannochloropsis sp. extract for cosmeceutical applications. Mar. Drugs 2021, 19, 690. [Google Scholar] [CrossRef]
- Oliva, A.K.; Bejaoui, M.; Hirano, A.; Arimura, T.; Linh, T.N.; Uchiage, E.; Nukaga, S.; Tominaga, K.; Nozaki, H.; Isoda, H. Elucidation of the potential hair growth-promoting effect of Botryococcus terribilis, its novel compound methylated-meijicoccenem and C32 botryococcene on cultured hair follicle dermal papilla cells using DNA microarray gene expression analysis. Biomedicines 2022, 10, 1186. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sato, K.; Hiraga, Y.; Yamaguchi, Y.; Sakaki, S.; Takenaka, H. Anti-Melanogenic and Anti-Oxidative Effects of Nostoc verrucosum (ashitsuki) Extracts. Cosmetics 2023, 10, 30. https://doi.org/10.3390/cosmetics10010030
Sato K, Hiraga Y, Yamaguchi Y, Sakaki S, Takenaka H. Anti-Melanogenic and Anti-Oxidative Effects of Nostoc verrucosum (ashitsuki) Extracts. Cosmetics. 2023; 10(1):30. https://doi.org/10.3390/cosmetics10010030
Chicago/Turabian StyleSato, Kazuomi, Yosuke Hiraga, Yuji Yamaguchi, Setsuko Sakaki, and Hiroyuki Takenaka. 2023. "Anti-Melanogenic and Anti-Oxidative Effects of Nostoc verrucosum (ashitsuki) Extracts" Cosmetics 10, no. 1: 30. https://doi.org/10.3390/cosmetics10010030
APA StyleSato, K., Hiraga, Y., Yamaguchi, Y., Sakaki, S., & Takenaka, H. (2023). Anti-Melanogenic and Anti-Oxidative Effects of Nostoc verrucosum (ashitsuki) Extracts. Cosmetics, 10(1), 30. https://doi.org/10.3390/cosmetics10010030