Impact of Selected Cosmetic Ingredients on Common Microorganisms of Healthy Human Skin

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Formulations
2.3. Microorganisms
2.4. Malassezia spp. Growth Conditions
2.5. Bacterial Growth Conditions
2.6. Determination of Malassezia spp. and Bacterial Growth
2.7. HPTLC
3. Results
3.1. Malassezia spp. Growth
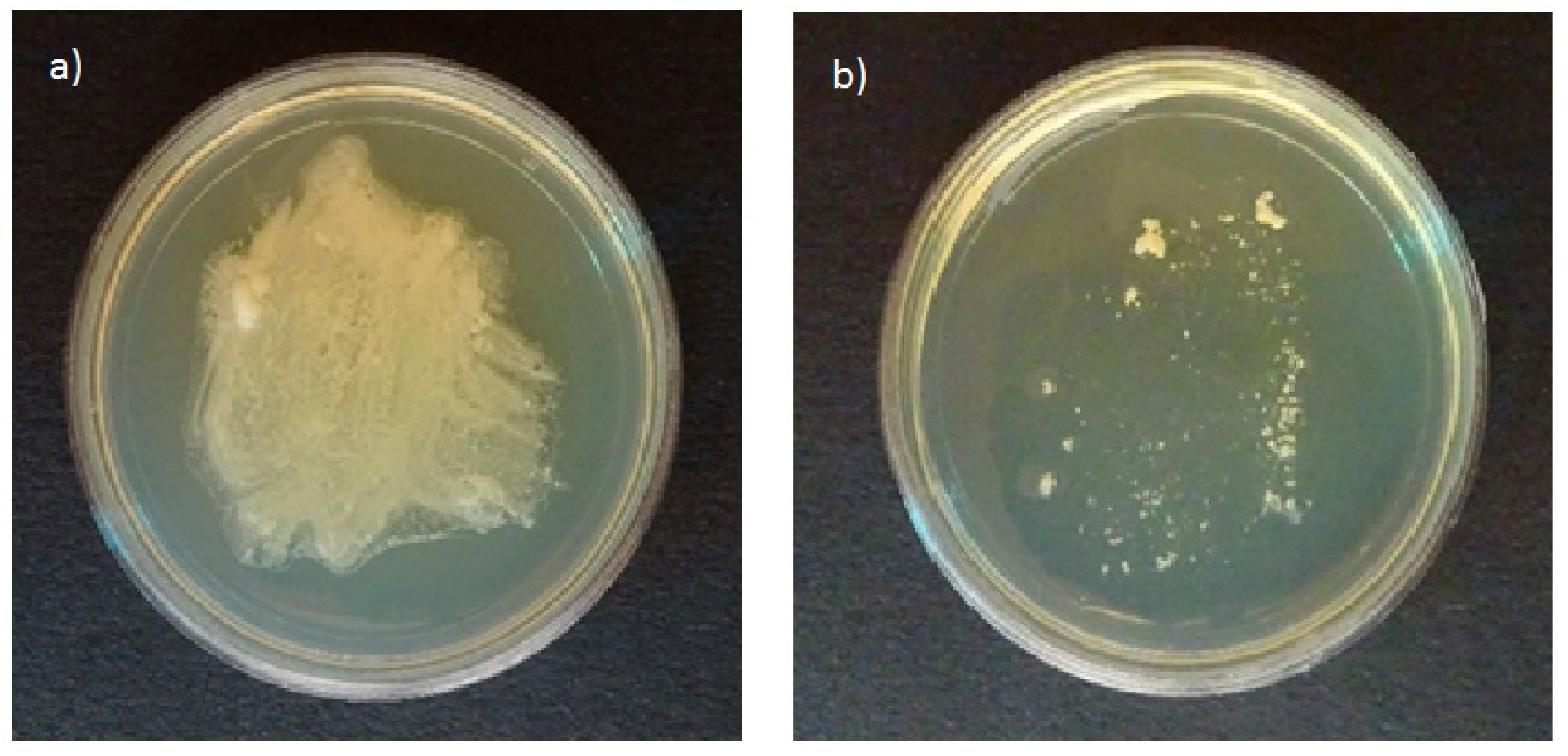
3.2. Bacterial Growth
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cundell, A.M. Microbial Ecology of the Human Skin. Microb. Ecol. 2018, 76, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Grice, E.A.; Dawson, T.L., Jr. Host-microbe interactions: Malassezia and human skin. Curr. Opin. Microbiol. 2017, 40, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.K. Seborrheic Dermatitis and Malassezia species: How Are They Related? J. Clin. Aesthet. Dermatol. 2009, 2, 14–17. [Google Scholar] [PubMed]
- Harada, K.; Saito, M.; Sugita, T.; Tsuboi, R. Malassezia species and their associated skin diseases. J. Dermatol. 2015, 42, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Jaeger, K.E.; Ransac, S.; Dijkstra, B.W.; Colson, C.; Vanheuvel, M.; Misset, O. Bacterial Lipases. FEMS Microbiol. Rev. 1994, 15, 29–63. [Google Scholar] [CrossRef] [PubMed]
- Mayser, P.; Gaitanis, G. Physiology and Biochemistry. In Malassezia and the Skin; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Pablo, G.; Hammons, A.; Bradley, S.; Fulton, J.E. Characteristics of the Extracellular Lipases from Corynebacterium Acnes and Staphylococcus Epidermis. J. Investig. Dermatol. 1974, 63, 231–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porro, M.N.; Passi, S.; Caprill, F.; Nazzaro, P.; Morpurgo, G. Growth requirements and lipid metabolism of Pityrosporum orbiculare. J. Investig. Dermatol. 1976, 66, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Gribbon, E.M.; Cunliffe, W.J.; Holland, K.T. Interaction of Propionibacterium acnes with skin lipids in vitro. J. Gen. Microbiol. 1993, 139, 1745–1751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rollof, J.; Braconier, J.H.; Soderstrom, C.; Nilsson-Ehle, P. Interference of Staphylococcus aureus lipase with human granulocyte function. Eur. J. Clin. Microbiol. Infect. Dis. 1988, 7, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, J.; Pommier, M.T.; Cremieux, A.; Michel, G. Influence of Tween 80 on the mycolic acid composition of three cutaneous corynebacteria. J. Gen. Microbiol. 1988, 134, 2457–2461. [Google Scholar] [CrossRef]
- Holland, K.T.; Bojar, R.A. Cosmetics: What is Their Influence on the Skin Microflora? Am. J. Clin. Dermatol. 2002, 3, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Mayser, P.; Haze, P.; Papavassilis, C.; Pickel, M.; Gruender, K.; Gueho, E. Differentiation of Malassezia species: Selectivity of cremophor EL, castor oil and ricinoleic acid for M. furfur. Br. J. Dermatol. 1997, 137, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Koyama, T.; Kanbe, T.; Kikuchi, A.; Tomita, Y. Effects of topical vehicles on growth of the lipophilic Malassezia species. J. Dermatol. Sci. 2002, 29, 166–170. [Google Scholar] [CrossRef]
- Nordstrom, K.M.; Labows, J.N.; McGinley, K.J.; Leyden, J.J. Characterization of wax esters, triglycerides, and free fatty acids of follicular casts. J. Invest. Dermatol. 1986, 86, 700–705. [Google Scholar] [CrossRef] [PubMed]
- Guého, E.; Midgley, G.; Guillot, J. The genus Malassezia with description of four new species. Antonie Van Leeuwenhoek 1996, 69, 337–355. [Google Scholar] [CrossRef] [PubMed]
- Mayser, P.; Fuhrer, D.; Schmidt, R.; Grunder, K. Hydrolysis of Fatty-Acid Esters by Malassezia-Furfur—Different Utilization Depending on Alcohol Moiety. Acta Derm. Venereol. 1995, 75, 105–109. [Google Scholar] [PubMed]
- Gannesen, A.V.; Borrel, V.; Lefeuvre, L.; Netrusov, A.I.; Plakunov, V.K.; Feuilloley, M.G.J. Effect of two cosmetic compounds on the growth, biofilm formation activity, and surface properties of acneic strains of Cutibacterium acnes and Staphylococcus aureus. Microbiologyopen 2019, 8, e00659. [Google Scholar] [CrossRef] [PubMed]
- Wallen-Russell, C. The Role of Every-Day Cosmetics in Altering the Skin Microbiome: A Study Using Biodiversity. Cosmetics 2018, 6, 2. [Google Scholar] [CrossRef]
- Lee, H.J.; Jeong, S.E.; Lee, S.; Kim, S.; Han, H.; Jeon, C.O. Effects of cosmetics on the skin microbiome of facial cheeks with different hydration levels. Microbiologyopen 2018, 7, e00557. [Google Scholar] [CrossRef]
- Jeong, J.-J.; Kim, D.-H. Effects of Cosmetics and Their Preservatives on the Growth and Composition of Human Skin Microbiota. J. Soc. Cosmet. Sci. Korea 2015, 41, 127–134. [Google Scholar]
- Lalitha, C.; Prasada Rao, P.V.V. Impact of superficial blends on skin micro biota. Int. J. Curr. Pharm. Res. 2013, 5, 61–65. [Google Scholar]
- Govi-Verlag Pharmazeutischer Verlag GmbH. Standardisierte Rezepturen (NRF/SR); Govi-Verlag Pharmazeutischer Verlag GmbH: Eschborn, Germany, 2011. [Google Scholar]
- Belsito, D.V.; Hill, R.A.; Klaassen, C.D.; Liebler, D.C.; Marks, J.G., Jr.; Shank, R.C.; Slaga, T.J.; Snyder, P.W. Plant-Derived Fatty Acid Oils as Used in Cosmetics. Final Rep. 2011, 1–100. Available online: https://purelyprofessional.dk/wp-content/uploads/inci/persea-gratissima-oil.pdf (accessed on 23 July 2019).
- Downing, D.T.; Stewart, M.E.; Wertz, P.W.; Colton, S.W.; Abraham, W.; Strauss, J.S. Skin lipids: An update. J. Investig. Dermatol. 1987, 88, 2s–6s. [Google Scholar] [CrossRef] [PubMed]
- Wilde, P.F.; Stewart, P.S. A study of the fatty acid metabolism of the yeast Pityrosporum ovale. Biochem. J. 1968, 108, 225–231. [Google Scholar] [CrossRef] [Green Version]
- Gordon James, A.; Abraham, K.H.; Cox, D.S.; Moore, A.E.; Pople, J.E. Metabolic analysis of the cutaneous fungi Malassezia globosa and M. restricta for insights on scalp condition and dandruff. Int. J. Cosmet. Sci. 2013, 35, 169–175. [Google Scholar] [CrossRef]
- Mayser, P.; Pickel, M.; Haze, P.; Erdmann, F.; Papavassilis, C.; Schmidt, R. Different utilization of neutral lipids by Malassezia furfur and Malassezia sympodialis. Med. Mycol. 1998, 36, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Connerth, M.; Czabany, T.; Wagner, A.; Zellnig, G.; Leitner, E.; Steyrer, E.; Daum, G. Oleate inhibits steryl ester synthesis and causes liposensitivity in yeast. J. Biol. Chem. 2010, 285, 26832–26841. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.B.; Alimova, Y.; Myers, T.M.; Ebersole, J.L. Short- and medium-chain fatty acids exhibit antimicrobial activity for oral microorganisms. Arch. Oral Biol. 2011, 56, 650–654. [Google Scholar] [CrossRef] [Green Version]
- Mayser, P. Medium chain fatty acid ethyl esters—Activation of antimicrobial effects by Malassezia enzymes. Mycoses 2015, 58, 215–219. [Google Scholar] [CrossRef]
- Papavassilis, C.; Mach, K.K.; Mayser, P.A. Medium-chain triglycerides inhibit growth of Malassezia: Implications for prevention of systemic infection. Crit. Care Med. 1999, 27, 1781–1786. [Google Scholar] [CrossRef]
- Vieira, O.V.; Hartmann, D.O.; Cardoso, C.M.P.; Oberdoerfer, D.; Baptista, M.; Santos, M.A.S.; Almeida, L.; Ramalho-Santos, J.; Vaz, W.L.C. Surfactants as microbicides and contraceptive agents: A systematic in vitro study. PLoS ONE 2008, 3, e2913. [Google Scholar] [CrossRef]
- Kwaszewska, A.; Sobiś-Glinkowska, M.; Szewczyk, E.M. Cohabitation—Relationships of corynebacteria and staphylococci on human skin. Folia Microbiol. 2014, 59, 495–502. [Google Scholar] [CrossRef]
- Sierra, G. A simple method for the detection of lipolytic activity of micro-organisms and some observations on the influence of the contact between cells and fatty substrates. Antonie Van Leeuwenhoek 1957, 23, 15–22. [Google Scholar] [CrossRef]
- Asada, Y. Lipolytic activity of resident flora of the skin: Some observations on lipase activity of Corynebacterium acnes and Staphylococcus epidermis compared with Staphylococcus aureus. Skin Res. 1968, 10, 585–593. [Google Scholar]
- Smith, R.F.; Willett, N.P. Lipolytic Activity of Human Cutaneous Bacteria. Microbiology 1968, 52, 441–445. [Google Scholar] [CrossRef] [Green Version]
- Freinkel, R.K.; Shen, Y. The Origin of Free Fatty Acids in Sebum II: Assay of the Lipases of the Cutaneous Bacteria and Effects of pH*. J. Investig. Dermatol. 1969, 53, 422–427. [Google Scholar] [CrossRef]
- Kabara, J.J.; Swieczkowski, D.M.; Conley, A.J.; Truant, J.P. Fatty acids and derivatives as antimicrobial agents. Antimicrob. Agents Chemother. 1972, 2, 23–28. [Google Scholar] [CrossRef]
- Galbraith, H.; Miller, T.B.; Paton, A.M.; Thompson, J.K. Antibacterial activity of long chain fatty acids and the reversal with calcium, magnesium, ergocalciferol and cholesterol. J. Appl. Bacteriol. 1971, 34, 803–813. [Google Scholar] [CrossRef]
- Smith, R.F. Fatty acid requirements of human cutaneous lipophilic corynebacteria. J. Gen. Microbiol. 1970, 60, 259–263. [Google Scholar] [CrossRef]
- Wille, J.J.; Kydonieus, A. Palmitoleic Acid Isomer (C16:1Δ6) in Human Skin Sebum Is Effective against Gram-Positive Bacteria. Skin Pharmacol. Physiol. 2003, 16, 176–187. [Google Scholar] [CrossRef]
- Ushijima, T.; Takahashi, M.; Ozaki, Y. Acetic, propionic, and oleic acid as the possible factors influencing the predominant residence of some species of Propionibacterium and coagulase-negative Staphylococcus on normal human skin. Can. J. Microbiol. 1984, 30, 647–652. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Kao, M.C.; Fang, J.-Y.; Zouboulis, C.C.; Zhang, L.; Gallo, R.L.; Huang, C.-M. Antimicrobial property of lauric acid against Propionibacterium acnes: Its therapeutic potential for inflammatory acne vulgaris. J. Investig. Dermatol. 2009, 129, 2480–2488. [Google Scholar] [CrossRef]
- Chao-Hsuan, C.; Wang, Y.; Nakatsuji, T.; Liu, Y.T.; Zouboulis, C.; Gallo, R.; Zhang, L.; Hsieh, M.F.; Huang, C.M. An innate bactericidal oleic acid effective against skin infection of methicillin-resistant Staphylococcus aureus: A therapy concordant with evolutionary medicine. J. Microbiol. Biotechnol. 2011, 21, 391–399. [Google Scholar]



{kind=link}
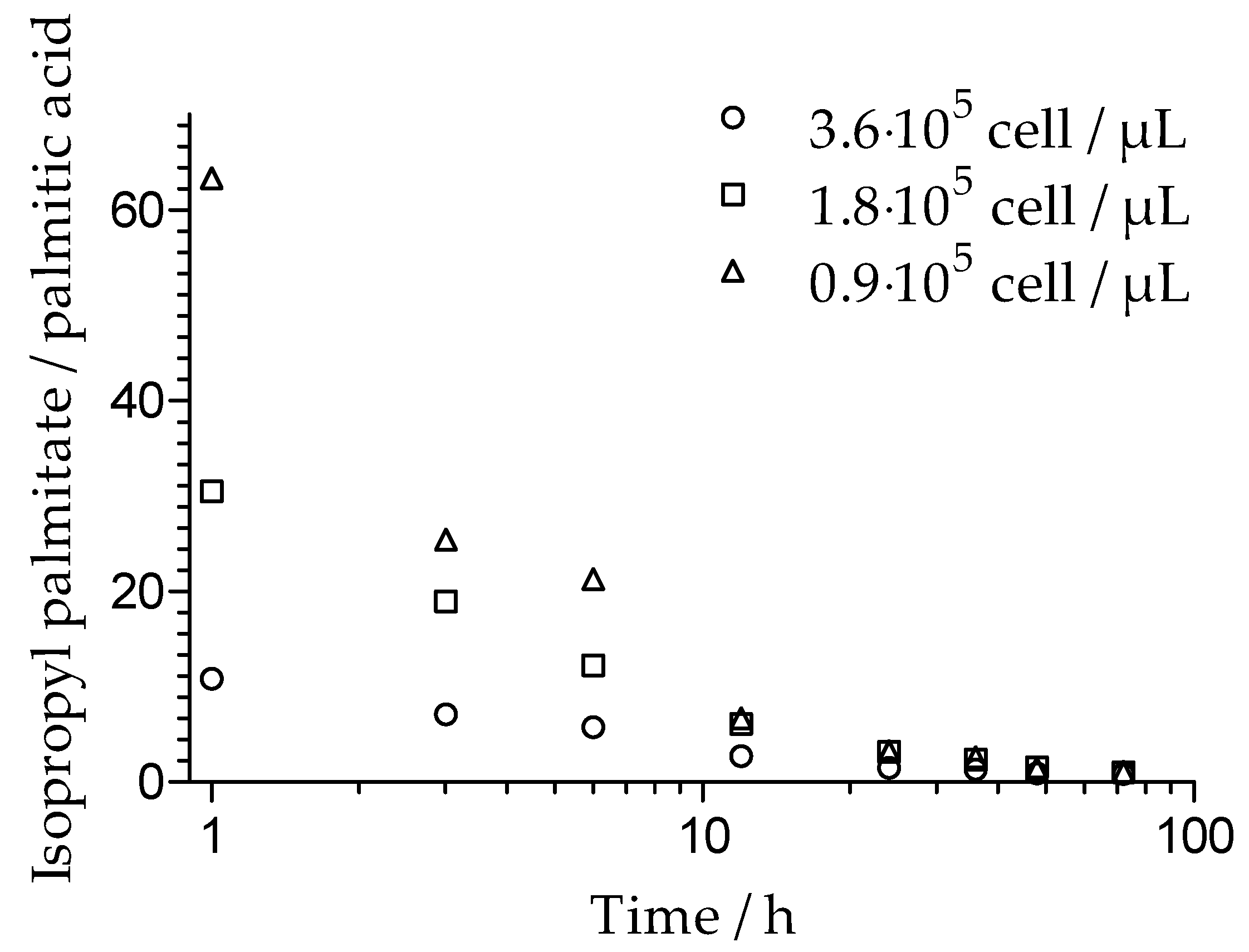{kind=link}
| Ingredient Group | International Nomenclature of Cosmetic Ingredients (INCI) | Producer |
|---|---|---|
| Native oils and waxes | Vitis vinifera Grape Seed Oil | Caelo |
| Olea europaea Fruit Oil | Caelo | |
| Persea gratissima Oil | Caelo | |
| Prunus amygdalus dulcis Oil | Caelo | |
| Oenothera biennis Oil | Caelo | |
| Triticum vulgare germ Oil | Caelo | |
| Brassica campestris oleifera Oil | Caelo | |
| Arachis hypogaea Oil | Caelo | |
| Glycine soja Oil | Caelo | |
| Sesamum indicum Seed Oil | Caelo | |
| Simmondsia chinensis (Jojoba) Seed Oil | Caelo | |
| Linum usitatissimum Seed Oil | Caelo | |
| Ricinus communis (Castor) Seed Oil | Caelo | |
| Cera alba (Bee wax) | Caelo | |
| Copernicia cerifera (Carnauba) Wax | Kahlwax | |
| Butyrospermum parkii (Shea) Butter | BTC Europe GmbH | |
| Fatty acid esters | Isopropyl myristate | Caelo |
| Isopropyl palmitate | Caelo | |
| Caprylic/capric triglyceride | Caelo | |
| Decyl oleate, coco-caprylate/caprate | BASF Personal Care and Nutrition GmbH | |
| Cetearyl isononanoate | BASF Personal Care and Nutrition GmbH | |
| Ethyl oleate | Croda | |
| Oleyl oleate | Fagron | |
| Decyl oleate | Caelo | |
| Cetearyl ethylhexanoate | Evonik | |
| Cetyl palmitate | Fagron | |
| Glyceryl stearate | Fagron | |
| PEG-40 stearate | Evonik | |
| Glyceryl oleate | Evonik | |
| Tween 80 | Caelo | |
| Span 80 | Fluka | |
| Macrogol 40 glycerolhydroxystearat | Fagron | |
| PEG-7 glyceryl cocoate | Evonik | |
| Sucrose stearate | SE Pharma | |
| Sucrose palmitate | SE Pharma | |
| PEG-40 hydrogenated castor oil | Evonik | |
| Methyl glucose isostearate | Evonik | |
| Polyglyceryl-3 caprate | Evonik | |
| Polyglyceryl-3 oleate | Evonik | |
| Tween 40 | TCI Deutschland GmbH | |
| Glyceryl stearate citrate | Evonik | |
| Polyglyceryl-3 dicitrate/stearate | Evonik | |
| Diisostearoyl polyglyceryl-3 dimer dilinoleate | Evonik | |
| Polyglyceryl-3 polyricinoleate | IOI OLEO GmbH | |
| Polyglyceryl-4 isostearate | Evonik | |
| Lanolin | Caelo | |
| Fatty acid alcohols | Octyldodecanol | Caelo |
| Cetearyl alcohol | Caelo | |
| Fatty alcohol ethers | Ceteth-20 | Croda |
| Oleth-20 | Acros organics | |
| Laureth 20 | Croda | |
| Steareth-20 | Croda | |
| Cetearyl glucoside | Evonik | |
| Fatty acids | Capric acid | Sigma-Aldrich |
| Palmitic acid | Carl Roth | |
| Oleic acid | Fagron | |
| Stearic acid | Cealo | |
| Other | Paraffin liquidum | Fagron |
| Dimethicone | Evonik | |
| Squalene | AlfaCesar | |
| Cyclopentasiloxane | Evonik | |
| Xanthan | Fagron | |
| Hydroxyethylcellulose | Fagron | |
| Cellulose gum (Carmellose natrium) | Fagron | |
| Carbomer | Caelo | |
| Acrylamide/sodium acryloyldimethyl taurate copolymer/isohexadecane and Polysorbate 80 | Seppic | |
| Hydroxyethyl acrylate/sodium acryloyldimethyl taurate copolymer | Seppic | |
| Cetyl PEG/PPG-10/1 dimethicone | Evonik | |
| Poloxamer 407 | Fagron | |
| Acrylates/C10-30 alkyl acrylate crosspolymer | Lubrizol | |
| Palmitamidopropyltrimonium chloride | Evonik | |
| PEG 300 | Caelo | |
| PEG 550 | Sigma-Aldrich |
| Oil-in-Water Emulsion | Water-in-Oil Emulsion |
|---|---|
| Water | Water |
| Pentylene glycol | Pentylene glycol |
| Magnesium sulfate | Magnesium sulfate |
| Glycerin | Glycerin |
| Sodium lactate | Sodium lactate |
| Hydroxyethyl acrylate/sodium acryloyl-dimethyl taurate copolymer | Cetyl PEG/PPG-10/1 dimethicone |
| Acrylates/C10-30 alkyl acrylate crosspolymer | Acrylates/C10-30 alkyl acrylate crosspolymer |
| Octyldodecanol | Octyldodecanol |
| Caprylic/capric triglyceride | Caprylic/capric triglyceride |
| Species | Strain No. |
|---|---|
| Malassezia furfur | CBS 1878 |
| Malassezia furfur | CBS 7019 |
| Malassezia sympodialis | CBS 7222 |
| Malassezia sympodialis | CBS 7977 |
| Malassezia sympodialis | ATCC 42132 |
| Malassezia sympodialis | DSM 6171 |
| Malassezia globosa | CBS 7966 |
| Malassezia globosa | CBS 7705 |
| Malassezia restricta | CBS 7877 |
| Corynebacterium minutissimum | DSM 20651 |
| Staphylococcus epidermidis | DSM 28764 |
| Staphylococcus galinarum | DSM 20610 |
| Substance | Concentration |
|---|---|
| Agar | 20 g/L |
| Lactose | 10 g/L |
| NaCl | 5 g/L |
| Substance | Band | M. Furfur | M. Globosa | M. Sympodialis |
|---|---|---|---|---|
| CBS 1878 | CBS 7705 | CBS 7222 | ||
| Oleic acid | 1 | 0.27 | 0.27 | - |
| 2 | 0.56 | 0.56 | - | |
| Oleyl oleate | 1 | 0.17 | 0.17 | - |
| 2 | 0.27 | 0.27 | - | |
| 3 | 0.56 | - | ||
| 4 | 0.65 | 0.65 | - | |
| Caprylic/capric triglyceride | 1 | 0.25 | 0.25 | 0.25 |
| 2 | 0.36 | 0.36 | 0.36 | |
| Isopropyl palmitate | 1 | 0.28 | 0.28 | 0.28 |
| 2 | 0.65 | 0.65 | 0.65 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dobler, D.; Schmidts, T.; Wildenhain, S.; Seewald, I.; Merzhäuser, M.; Runkel, F. Impact of Selected Cosmetic Ingredients on Common Microorganisms of Healthy Human Skin. Cosmetics 2019, 6, 45. https://doi.org/10.3390/cosmetics6030045
Dobler D, Schmidts T, Wildenhain S, Seewald I, Merzhäuser M, Runkel F. Impact of Selected Cosmetic Ingredients on Common Microorganisms of Healthy Human Skin. Cosmetics. 2019; 6(3):45. https://doi.org/10.3390/cosmetics6030045
Chicago/Turabian StyleDobler, Dorota, Thomas Schmidts, Sören Wildenhain, Ilona Seewald, Michael Merzhäuser, and Frank Runkel. 2019. "Impact of Selected Cosmetic Ingredients on Common Microorganisms of Healthy Human Skin" Cosmetics 6, no. 3: 45. https://doi.org/10.3390/cosmetics6030045
APA StyleDobler, D., Schmidts, T., Wildenhain, S., Seewald, I., Merzhäuser, M., & Runkel, F. (2019). Impact of Selected Cosmetic Ingredients on Common Microorganisms of Healthy Human Skin. Cosmetics, 6(3), 45. https://doi.org/10.3390/cosmetics6030045