

Somatic Cell Reprogramming for Nervous System Diseases: Techniques, Mechanisms, Potential Applications, and Challenges


Abstract
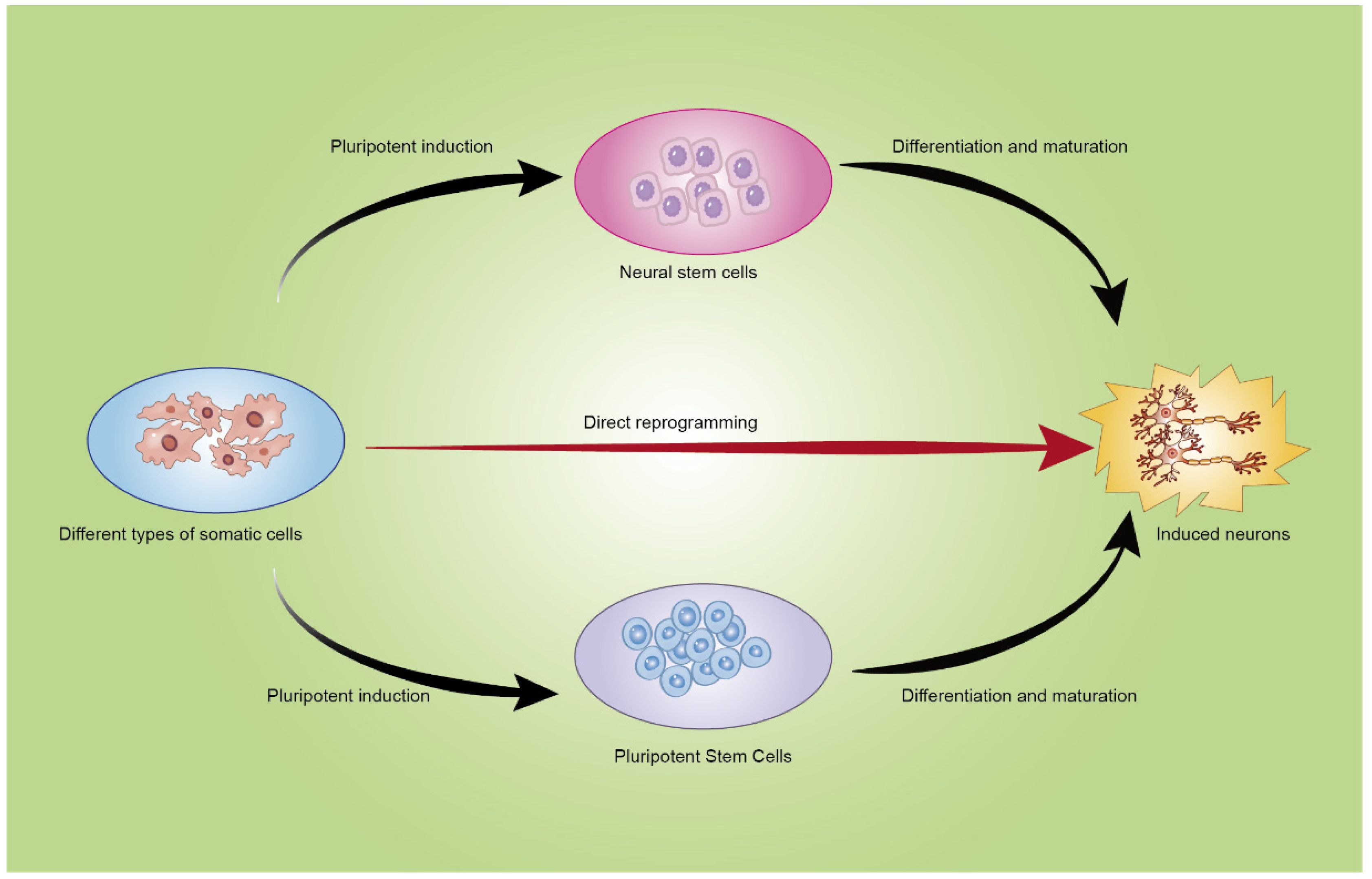
:1. Introduction
2. Cell Types That Have Been Researched for Transdifferentiation into Neurons
3. Mature Somatic Cells Reprogram into Neurons through Different Pathways
4. Transcription Factors, Small Molecules, and miRNAs That Induce Transdifferentiation of Somatic Cells into Neurons

{kind=link}
| Transcription Factor | Family Affiliation | Role in Neurogenesis, Differentiation, or Reprogramming | References |
|---|---|---|---|
| ASCL1 | bHLH family | Determination of neuronal subtypes during neural development. | [70] |
| ATOH1 | bHLH family | Unknown. | [71] |
| BCL11B | COUP TF1-interacting protein 2 (also known as Ctip2) and zinc finger-containing transcriptional repressors | Central to differentiation of medium spiny neurons and development of the striatum. | [72] |
| BCL2 | Anti-apoptotic factor | Promotes DNA damage, genetic instability, and cell proliferation. | [33,73,74] |
| BRCA1 | Tumor suppressor protein | Involved in BMP-2-mediated reactivation of Sox2. | [51] |
| BRN2a | Brain-specific homeobox/POU domain protein 2 | Associated with neuroendocrine function. | [6,75,76] |
| CEND1 | Neurogenic protein | Pathways involved in neuronal differentiation by CEND1 through activation by NEUROGENIN 1 and 2. | [77] |
| EBF1 | Zinc finger | Acting downstream of Ngn, EBF-1 can promote ectopic neurogenesis. | [78,79] |
| FEZF2 | Zinc finger transcriptional repressor | Fezf2 manipulates the origin, specific differentiation, and synaptic connectivity of corticospinal motor neurons by regulating neural progenitor cell lineage-directed differentiation signals. | [80] |
| FOXA2 | Forkhead | Expressed in the ventral hindbrain’s serotonergic progenitor regions and in midbrain dopaminergic neurons. | [28,81] |
| FOXG1 | Forkhead | Involved in the primitive (anterior) neuroectoderm during development of embryonic stem cells. | [82,83] |
| GATA3 | Zinc finger | Associated with noradrenergic phenotype and development of the sympathetic nervous system. | [84,85] |
| GATA4 | GATA | GATA4 can drive embryonic Sertoli-like cell differentiation. | [86] |
| HAND2 | bHLH family | Required for the acquisition of noradrenergic phenotype. | [84,85,87] |
| ISL2 | LIM homeodomain-containing | Vital to the development and differentiation of visceral motor neurons in the spinal cord. | [78,88] |
| KLF4 | Zinc finger | Directly represses p53. | [13] |
| LIN28 | RNA binding protein | Lin-28 can shuttle between the nucleus and cytoplasm and regulate other genes that control the cell cycle. | [89] |
| LMO2 | Key hematopoietic transcriptional regulator | Creates a regulatory complex that mediates transcription of multiple genes in hematopoietic progenitor cells; it is associated with the transcriptional control of stem/progenitor cells. | [90,91] |
| c-Myc | Myc | Alters expression of many proteins to enhance proliferation and transformation. | [13] |
| MYT1L | Neural zinc finger | Exemplifies a class of neural sequence-specific transcription factors that actively recruit histone deacetylases to selected genes during central nervous system development. | [92,93] |
| NANOG | Divergent homeodomain protein | NANOG sustains the identity of embryonic stem cells (ESCs). | [94] |
| NEUROD1 | bHLH family | Neural differentiation factor essential for late-stage neurogenesis and important in the development of the central nervous system, as well as in the auditory and vestibular systems. | [35,95,96] |
| NEUROD2 | bHLH family | Essential for the maturation and survival of neurons in the central nervous system. | [97,98,99] |
| NEUROG2 | bHLH family | NEUROG2 is a key contributor to early neurogenesis. | [67] |
| NURR1 | Nr4a2 (ligand-independent nuclear receptors) | Essential for the differentiation, maturation, and maintenance of midbrain dopaminergic neurons. | [100,101] |
| OCT/4 | POU5F1, a member of the POU class of homeodomain proteins | Central to the transcriptional regulatory hierarchy that specifies embryonic stem cell identity during early development. | [102] |
| OLIG2 | Basic helix–loop–helix | Mediates self-renewal in the expansion of neurosphere cultures and promotes the generation of neurons and oligodendrocytes under differentiation conditions. | [103] |
| PAX6 | Paired-box family | Critically important in multiple cell types and at several stages of forebrain development. | [104] |
| PHOX2A | Paired homeodomain | Selectively expressed and required for the specification of ventral motor neurons in the hindbrain and in the oculomotor nucleus, located laterally to dopaminergic neurons in the ventral midbrain. | [105] |
| Phox2B | Paired homeodomain | Same as Phox2a. | [105] |
| PTF1A | Basic helix–loop–helix | Mostly expressed in post-mitotic cells, and it specifies terminal cell fate in neural tissues. | [106] |
| SOX2 | HMG-box | Central to the transcriptional regulatory hierarchy that specifies embryonic stem cell identity during early development. | [102] |
| SOX4 | SoxC | Controls the survival of neural precursors and their differentiated progeny, in redundancy with SOX11. | [107] |
| SOX11 | SoxC | Same as SOX4. | [107] |
| SV40LT | SV40 large T gene | Unknown. | [108] |
| TLX3 | Tlx-class homeobox genes | Tlx3 functions as a post-mitotic selection gene in the embryonic spinal cord, determining the fate of dorsal glutamatergic neuronal cells. | [78,109] |
| ZEB1 | Zinc finger E-box-binding transcription factor | During individual development, Zeb1 plays a crucial role in the nervous system. It is upregulated in growing neurons throughout the central nervous system and is required for the survival of spinal cord neural stem cells. | [110] |
| Small Molecule | Description | References |
|---|---|---|
| 616452 | Repsox, an ALK5 inhibitor. | [129] |
| 17-allylaminogeldanamycin | GSK3 inhibitor. | [129] |
| A83-01 | ALK4/5/7 inhibitor. | [38,130,131] |
| All-trans retinoic acid | Physiologically active metabolite of vitamin A. | [53,132,133] |
| AM580 | Retinoic acid agonist, stable benzoic derivative of retinoic acid. | [129,134] |
| Apicidin | HDAC inhibitor. | [51] |
| Azacytidine | Nucleic acid synthesis inhibitor. | [135] |
| Blebbistatin | NMII inhibitor. | [136] |
| Bradykinin | Bradykinin plays a role in neural fate determination and facilitates neurogenesis and migration. | [137] |
| CH55 | Synthetic stable analog of retinoic acid. | [138] |
| CHIR99021 | GSK3 inhibitor. | [129] |
| CpdE | Notch signaling pathway inhibitor. | [139] |
| DAPT | Inhibits γ-secretase and Notch signaling. | [53,140] |
| DMH1 | BMP type I receptor inhibitor. | [139] |
| Dorsomorphin | Inhibitor of AMP-activated protein kinase and bone morphogenetic protein type 1 receptor. | [67] |
| DZNep | Histone methylation inhibitor. | [141] |
| EPZ004777 | Dot1l inhibitor. | [138,142] |
| Forskolin | cAMP agonist. | [129] |
| GSK3β inhibitor | Glycogen synthase kinase-3β boosts the production of neuroprotective and neurotrophic factors in the context of spinal cord injury. | [143] |
| GO6983 | PKC inhibitor. | [54,144] |
| Hh-Ag1.5 | Unknown. | [131] |
| I-BET 151 | BET family bromodomain inhibitor. | [145] |
| Insulin–transferrin–selenium | Insulin, transferring, and sodium selenium compound. | [135] |
| Isoxazole | Isoxazole is able to upregulate proneural marker genes and exhibit regulation of stem cells. | [41] |
| ISX9 | Induces neuronal differentiation through myocyte enhancer factor 2 (Mef2), which is a vital pathway for neural differentiation and maturation. | [128] |
| Kenpaullone | GSK-3β inhibitor. | [146] |
| LDN193189 | Inhibitor of bone morphogenetic protein type I receptors ALK2 and ALK3, used to suppress specification of mesoderm and endoderm. | [147] |
| LIF | Leukemia inhibitory factor. | [148,149] |
| LM-22A4 | Growth factor. | [150] |
| Mercaptoethanol | Unknown. | [148] |
| MS-275 | Benzamide. | [51] |
| Niclosamide | Wnt signaling inhibitor. | [34] |
| Noggin | SMAD inhibitor. | [151] |
| NT3 | Unknown. | [150] |
| P7C3-A20 | May stimulate NAMPT-relevant pathways to exert neurogenesis. | [24] |
| Parnate | Lysine-specific demethylase 1 inhibitor. | [24] |
| PD0325901 | Mitogen-activated protein kinase inhibitor. | [24] |
| PS48 | PDK1 activator. | [152] |
| Purmorphamine | Activator of the Shh signaling pathway. | [153] |
| Quercetin | PI3K signaling inhibitor. | [34] |
| QVD-OPH | Caspase inhibitor. | [144] |
| Repsox | Transforming growth factor-β inhibitor. | [154] |
| Retinoic acid | Induces neurogenesis and neuronal differentiation by activating retinoic acid receptors. | [38] |
| RG108 | DNA methyltransferase inhibitor that is less toxic to cells than parnate. | [24] |
| Ruxolitinib | Selective JAK1/2 inhibitor. | [155] |
| SB203580 | P38 MARK inhibitor. | [145,155] |
| SB43152 | Unknown. | [153] |
| SB431542 | Inhibits TGF-β type I receptors ALK4, ALK5, and ALK7. | [30,147] |
| SB4352 | Transforming growth factor-beta inhibitor | [36] |
| SMER28 | SMER28 shows neurotrophic and neuroprotective effects by inducing neurite growth and protecting against excitotoxin-induced axonal degeneration. | [156,157] |
| Smoothened agonist | Alone or in concert with other molecules, smoothened agonist stimulates proliferation of primary neuronal precursor cells. | [158,159] |
| Sodium butyrate | HDAC inhibitor, causes hyperacetylation of histones. | [38] |
| Sonic hedgehog (SHH) | Required for the development of dopaminergic neurons in multiple locations along the anterior neural tube. | [160,161] |
| SP600125 | 1,9-pyrazoloanthrone, JNK inhibitor. | [162] |
| SP600625 | JNK inhibitor. | [144] |
| TD114-2 | GSK3-beta inhibitor, preferred over CHIR99021. | [138] |
| Transforming growth factor beta 3 | Required for the induction, differentiation, and survival of midbrain dopaminergic neurons. | [163] |
| Thiazovivin | Unknown. | [164] |
| Tranylcypromine | Lysine-specific histone demethylase LSD1 inhibitor. | [141] |
| Trichostatin A | Histone deacetylase inhibitor. | [165] |
| TTNPB | Agonist of retinoic acid receptors, which play an important role in neural differentiation. | [164] |
| Valproic acid | Inhibits histone deacetylase activity. | [166] |
| Vitamin C | Prevents cell death. | [144] |
| Y-27632 | Rho-associated protein kinase inhibitor. | [162] |
5. Molecular Mechanisms of Somatic Cell Transdifferentiation into Neurons
6. Somatic Cell Transdifferentiation Provides New Possibilities for Research and Treatment of Neurological Disease
7. Limitations of Neural Reprogramming
8. Discussion
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Prabhakaran, D.; Anand, S.; Watkins, D.; Gaziano, T.; Wu, Y.; Mbanya, J.C.; Nugent, R. Cardiovascular, respiratory, and related disorders: Key messages from Disease Control Priorities, 3rd edition. Lancet 2018, 391, 1224–1236. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Sharma, R.; Zhang, Z.; Liaw, K.; Kambhampati, S.P.; Porterfield, J.E.; Lin, K.C.; DeRidder, L.B.; Kannan, S.; Kannan, R.M. Dense hydroxyl polyethylene glycol dendrimer targets activated glia in multiple CNS disorders. Sci. Adv. 2020, 6, eaay8514. [Google Scholar] [CrossRef] [PubMed]
- Collaborators, G.D.a.I. Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar] [CrossRef]
- Tasoglu, S.; Gurkan, U.A.; Wang, S.; Demirci, U. Manipulating biological agents and cells in micro-scale volumes for applications in medicine. Chem. Soc. Rev. 2013, 42, 5788–5808. [Google Scholar] [CrossRef]
- Grade, S.; Götz, M. Neuronal replacement therapy: Previous achievements and challenges ahead. NPJ Regen. Med. 2017, 2, 29. [Google Scholar] [CrossRef]
- Torper, O.; Pfisterer, U.; Wolf, D.A.; Pereira, M.; Lau, S.; Jakobsson, J.; Björklund, A.; Grealish, S.; Parmar, M. Generation of induced neurons via direct conversion in vivo. Proc. Natl. Acad. Sci. USA 2013, 110, 7038–7043. [Google Scholar] [CrossRef]
- Denoth-Lippuner, A.; Jessberger, S. Formation and integration of new neurons in the adult hippocampus. Nat. Rev. Neurosci. 2021, 22, 223–236. [Google Scholar] [CrossRef]
- Faiz, M.; Sachewsky, N.; Gascón, S.; Bang, K.W.; Morshead, C.M.; Nagy, A. Adult Neural Stem Cells from the Subventricular Zone Give Rise to Reactive Astrocytes in the Cortex after Stroke. Cell Stem Cell 2015, 17, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, D.; DeWitt, N. In Vivo Cellular Reprogramming: The Next Generation. Cell 2016, 166, 1386–1396. [Google Scholar] [CrossRef]
- Gurdon, J.B.; Elsdale, T.R.; Fischberg, M. Sexually mature individuals of Xenopus laevis from the transplantation of single somatic nuclei. Nature 1958, 182, 64–65. [Google Scholar] [CrossRef]
- Wilmut, I.; Schnieke, A.E.; McWhir, J.; Kind, A.J.; Campbell, K.H. Viable offspring derived from fetal and adult mammalian cells. Nature 1997, 385, 810–813. [Google Scholar] [CrossRef]
- Wakayama, T.; Perry, A.C.; Zuccotti, M.; Johnson, K.R.; Yanagimachi, R. Full-term development of mice from enucleated oocytes injected with cumulus cell nuclei. Nature 1998, 394, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef]
- de Leeuw, V.C.; van Oostrom, C.T.M.; Imholz, S.; Piersma, A.H.; Hessel, E.V.S.; Dollé, M.E.T. Going Back and Forth: Episomal Vector Reprogramming of Peripheral Blood Mononuclear Cells to Induced Pluripotent Stem Cells and Subsequent Differentiation into Cardiomyocytes and Neuron-Astrocyte Co-cultures. Cell. Reprogram. 2020, 22, 300–310. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Habekost, M.; Jorgensen, A.L.; Qvist, P.; Denham, M. MicroRNAs and Ascl1 facilitate direct conversion of porcine fibroblasts into induced neurons. Stem Cell Res. 2020, 48, 101984. [Google Scholar] [CrossRef]
- Onorati, M.; Camnasio, S.; Binetti, M.; Jung, C.B.; Moretti, A.; Cattaneo, E. Neuropotent self-renewing neural stem (NS) cells derived from mouse induced pluripotent stem (iPS) cells. Mol. Cell. Neurosci. 2010, 43, 287–295. [Google Scholar] [CrossRef]
- Wu, Z.; Parry, M.; Hou, X.Y.; Liu, M.H.; Wang, H.; Cain, R.; Pei, Z.F.; Chen, Y.C.; Guo, Z.Y.; Abhijeet, S.; et al. Gene therapy conversion of striatal astrocytes into GABAergic neurons in mouse models of Huntington’s disease. Nat. Commun. 2020, 11, 1105. [Google Scholar] [CrossRef] [PubMed]
- Rivetti di Val Cervo, P.; Romanov, R.A.; Spigolon, G.; Masini, D.; Martín-Montañez, E.; Toledo, E.M.; La Manno, G.; Feyder, M.; Pifl, C.; Ng, Y.H.; et al. Induction of functional dopamine neurons from human astrocytes in vitro and mouse astrocytes in a Parkinson’s disease model. Nat. Biotechnol. 2017, 35, 444–452. [Google Scholar] [CrossRef]
- Chen, Y.C.; Ma, N.X.; Pei, Z.F.; Wu, Z.; Do-Monte, F.H.; Keefe, S.; Yellin, E.; Chen, M.S.; Yin, J.C.; Lee, G.; et al. A NeuroD1 AAV-Based Gene Therapy for Functional Brain Repair after Ischemic Injury through In Vivo Astrocyte-to-Neuron Conversion. Mol. Ther. 2020, 28, 217–234. [Google Scholar] [CrossRef]
- Zhou, H.; Su, J.; Hu, X.; Zhou, C.; Li, H.; Chen, Z.; Xiao, Q.; Wang, B.; Wu, W.; Sun, Y.; et al. Glia-to-Neuron Conversion by CRISPR-CasRx Alleviates Symptoms of Neurological Disease in Mice. Cell 2020, 181, 590–603.e516. [Google Scholar] [CrossRef]
- Zaret, K.S. Pioneer Transcription Factors Initiating Gene Network Changes. Annu. Rev. Genet. 2020, 54, 367–385. [Google Scholar] [CrossRef] [PubMed]
- Löhle, M.; Hermann, A.; Glass, H.; Kempe, A.; Schwarz, S.C.; Kim, J.B.; Poulet, C.; Ravens, U.; Schwarz, J.; Schöler, H.R.; et al. Differentiation efficiency of induced pluripotent stem cells depends on the number of reprogramming factors. Stem. Cells 2012, 30, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chen, R.; Wu, X.; Zhao, Y.; Fan, Y.; Xiao, Z.; Han, J.; Sun, L.; Wang, X.; Dai, J. Rapid and Efficient Conversion of Human Fibroblasts into Functional Neurons by Small Molecules. Stem Cell Rep. 2019, 13, 862–876. [Google Scholar] [CrossRef]
- Zhou, C.; Gu, H.; Fan, R.; Wang, B.; Lou, J. MicroRNA 302/367 Cluster Effectively Facilitates Direct Reprogramming from Human Fibroblasts into Functional Neurons. Stem Cells Dev. 2015, 24, 2746–2755. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi-Kasman, M.; Shojaei, A.; Gol, M.; Moghadamnia, A.A.; Baharvand, H.; Javan, M. miR-302/367-induced neurons reduce behavioral impairment in an experimental model of Alzheimer’s disease. Mol. Cell. Neurosci. 2018, 86, 50–57. [Google Scholar] [CrossRef]
- Liang, X.G.; Tan, C.; Wang, C.K.; Tao, R.R.; Huang, Y.J.; Ma, K.F.; Fukunaga, K.; Huang, M.Z.; Han, F. Myt1l induced direct reprogramming of pericytes into cholinergic neurons. CNS Neurosci. Ther. 2018, 24, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Niu, W.; Zang, T.; Wang, L.L.; Zou, Y.; Zhang, C.L. Phenotypic Reprogramming of Striatal Neurons into Dopaminergic Neuron-like Cells in the Adult Mouse Brain. Stem Cell Rep. 2018, 11, 1156–1170. [Google Scholar] [CrossRef]
- Tanabe, K.; Ang, C.E.; Chanda, S.; Olmos, V.H.; Haag, D.; Levinson, D.F.; Sudhof, T.C.; Wernig, M. Transdifferentiation of human adult peripheral blood T cells into neurons. Proc. Natl. Acad. Sci. USA 2018, 115, 6470–6475. [Google Scholar] [CrossRef]
- Bossolasco, P.; Sassone, F.; Gumina, V.; Peverelli, S.; Garzo, M.; Silani, V. Motor neuron differentiation of iPSCs obtained from peripheral blood of a mutant TARDBP ALS patient. Stem Cell Res. 2018, 30, 61–68. [Google Scholar] [CrossRef]
- Marro, S.; Pang, Z.P.; Yang, N.; Tsai, M.C.; Qu, K.; Chang, H.Y.; Sudhof, T.C.; Wernig, M. Direct lineage conversion of terminally differentiated hepatocytes to functional neurons. Cell Stem Cell 2011, 9, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Robicsek, O.; Karry, R.; Petit, I.; Salman-Kesner, N.; Müller, F.J.; Klein, E.; Aberdam, D.; Ben-Shachar, D. Abnormal neuronal differentiation and mitochondrial dysfunction in hair follicle-derived induced pluripotent stem cells of schizophrenia patients. Mol. Psychiatry 2013, 18, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Gascon, S.; Murenu, E.; Masserdotti, G.; Ortega, F.; Russo, G.L.; Petrik, D.; Deshpande, A.; Heinrich, C.; Karow, M.; Robertson, S.P.; et al. Identification and Successful Negotiation of a Metabolic Checkpoint in Direct Neuronal Reprogramming. Cell Stem Cell 2016, 18, 396–409. [Google Scholar] [CrossRef]
- Radhakrishnan, S.; Martin, C.A.; Dhayanithy, G.; Reddy, M.S.; Rela, M.; Kalkura, S.N.; Sellathamby, S. Hypoxic Preconditioning Induces Neuronal Differentiation of Infrapatellar Fat Pad Stem Cells through Epigenetic Alteration. ACS Chem. Neurosci. 2021, 12, 704–718. [Google Scholar] [CrossRef] [PubMed]
- Puls, B.; Ding, Y.; Zhang, F.; Pan, M.; Lei, Z.; Pei, Z.; Jiang, M.; Bai, Y.; Forsyth, C.; Metzger, M.; et al. Regeneration of Functional Neurons After Spinal Cord Injury via in situ NeuroD1-Mediated Astrocyte-to-Neuron Conversion. Front. Cell Dev. Biol. 2020, 8, 591883. [Google Scholar] [CrossRef]
- El Ayachi, I.; Zhang, J.; Zou, X.Y.; Li, D.; Yu, Z.; Wei, W.; O’Connell, K.M.S.; Huang, G.T. Human dental stem cell derived transgene-free iPSCs generate functional neurons via embryoid body-mediated and direct induction methods. J. Tissue Eng. Regen. Med. 2018, 12, e1836–e1851. [Google Scholar] [CrossRef]
- Castaño, J.; Menendez, P.; Bruzos-Cidon, C.; Straccia, M.; Sousa, A.; Zabaleta, L.; Vazquez, N.; Zubiarrain, A.; Sonntag, K.C.; Ugedo, L.; et al. Fast and efficient neural conversion of human hematopoietic cells. Stem Cell Rep. 2014, 3, 1118–1131. [Google Scholar] [CrossRef]
- Xu, G.; Wu, F.; Gu, X.; Zhang, J.; You, K.; Chen, Y.; Getachew, A.; Zhuang, Y.; Zhong, X.; Lin, Z.; et al. Direct Conversion of Human Urine Cells to Neurons by Small Molecules. Sci. Rep. 2019, 9, 16707. [Google Scholar] [CrossRef]
- Sun, X.; Tan, Z.; Huang, X.; Cheng, X.; Yuan, Y.; Qin, S.; Wang, D.; Hu, X.; Gu, Y.; Qian, W.J.; et al. Direct neuronal reprogramming of olfactory ensheathing cells for CNS repair. Cell Death Dis. 2019, 10, 646. [Google Scholar] [CrossRef]
- Yang, H.; Hao, D.; Liu, C.; Huang, D.; Chen, B.; Fan, H.; Liu, C.; Zhang, L.; Zhang, Q.; An, J.; et al. Generation of functional dopaminergic neurons from human spermatogonial stem cells to rescue parkinsonian phenotypes. Stem Cell Res. Ther. 2019, 10, 195. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, P.; Hsu, T.; Aguilar, H.R.; Frantz, D.E.; Schneider, J.W.; Bachoo, R.M.; Hsieh, J. Small-molecule blocks malignant astrocyte proliferation and induces neuronal gene expression. Differ. Res. Biol. Divers. 2011, 81, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, T.; Irie, T.; Katsurabayashi, S.; Hayashi, Y.; Nagai, T.; Hamazaki, N.; Adefuin, A.M.D.; Miura, F.; Ito, T.; Kimura, H.; et al. Pioneer Factor NeuroD1 Rearranges Transcriptional and Epigenetic Profiles to Execute Microglia-Neuron Conversion. Neuron 2019, 101, 472–485.e477. [Google Scholar] [CrossRef]
- Rouaux, C.; Arlotta, P. Direct lineage reprogramming of post-mitotic callosal neurons into corticofugal neurons in vivo. Nat. Cell Biol. 2013, 15, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Noda, T.; Meas, S.J.; Nogami, J.; Amemiya, Y.; Uchi, R.; Ohkawa, Y.; Nishimura, K.; Dabdoub, A. Direct Reprogramming of Spiral Ganglion Non-neuronal Cells into Neurons: Toward Ameliorating Sensorineural Hearing Loss by Gene Therapy. Front. Cell Dev. Biol. 2018, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Chuang, W.; Sharma, A.; Shukla, P.; Li, G.; Mall, M.; Rajarajan, K.; Abilez, O.J.; Hamaguchi, R.; Wu, J.C.; Wernig, M.; et al. Partial Reprogramming of Pluripotent Stem Cell-Derived Cardiomyocytes into Neurons. Sci. Rep. 2017, 7, 44840. [Google Scholar] [CrossRef]
- Weinberg, M.S.; Criswell, H.E.; Powell, S.K.; Bhatt, A.P.; McCown, T.J. Viral Vector Reprogramming of Adult Resident Striatal Oligodendrocytes into Functional Neurons. Mol. Ther. 2017, 25, 928–934. [Google Scholar] [CrossRef]
- Torper, O.; Ottosson, D.R.; Pereira, M.; Lau, S.; Cardoso, T.; Grealish, S.; Parmar, M. In Vivo Reprogramming of Striatal NG2 Glia into Functional Neurons that Integrate into Local Host Circuitry. Cell Rep. 2015, 12, 474–481. [Google Scholar] [CrossRef]
- Cheng, F.; Lu, X.C.; Hao, H.Y.; Dai, X.L.; Da Qian, T.; Huang, B.S.; Tang, L.J.; Yu, W.; Li, L.X. Neurogenin 2 Converts Mesenchymal Stem Cells into a Neural Precursor Fate and Improves Functional Recovery after Experimental Stroke. Cell. Physiol. Biochem. 2014, 33, 847–858. [Google Scholar] [CrossRef]
- Pollak, J.; Wilken, M.S.; Ueki, Y.; Cox, K.E.; Sullivan, J.M.; Taylor, R.J.; Levine, E.M.; Reh, T.A. ASCL1 reprograms mouse Muller glia into neurogenic retinal progenitors. Development 2013, 140, 2619–2631. [Google Scholar] [CrossRef]
- Giorgetti, A.; Marchetto, M.C.; Li, M.; Yu, D.; Fazzina, R.; Mu, Y.; Adamo, A.; Paramonov, I.; Cardoso, J.C.; Monasterio, M.B.; et al. Cord blood-derived neuronal cells by ectopic expression of Sox2 and c-Myc. Proc. Natl. Acad. Sci. USA 2012, 109, 12556–12561. [Google Scholar] [CrossRef]
- Lyssiotis, C.A.; Walker, J.; Wu, C.; Kondo, T.; Schultz, P.G.; Wu, X. Inhibition of histone deacetylase activity induces developmental plasticity in oligodendrocyte precursor cells. Proc. Natl. Acad. Sci. USA 2007, 104, 14982–14987. [Google Scholar] [CrossRef]
- Grinnell, K.L.; Yang, B.; Eckert, R.L.; Bickenbach, J.R. De-differentiation of mouse interfollicular keratinocytes by the embryonic transcription factor Oct-4. J. Investig. Dermatol. 2007, 127, 372–380. [Google Scholar] [CrossRef]
- Long, Q.; Luo, Q.; Wang, K.; Bates, A.; Shetty, A.K. Mash1-dependent Notch Signaling Pathway Regulates GABAergic Neuron-Like Differentiation from Bone Marrow-Derived Mesenchymal Stem Cells. Aging Dis. 2017, 8, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Qiu, B.; Guan, W.; Wang, Q.; Wang, M.; Li, W.; Gao, L.; Shen, L.; Huang, Y.; Xie, G.; et al. Direct Conversion of Normal and Alzheimer’s Disease Human Fibroblasts into Neuronal Cells by Small Molecules. Cell Stem Cell 2015, 17, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Dewald, L.E.; Rodriguez, J.P.; Levine, J.M. The RE1 binding protein REST regulates oligodendrocyte differentiation. J. Neurosci. 2011, 31, 3470–3483. [Google Scholar] [CrossRef] [PubMed]
- Boshans, L.L.; Soh, H.; Wood, W.M.; Nolan, T.M.; Mandoiu, I.I.; Yanagawa, Y.; Tzingounis, A.V.; Nishiyama, A. Direct reprogramming of oligodendrocyte precursor cells into GABAergic inhibitory neurons by a single homeodomain transcription factor Dlx2. Sci. Rep. 2021, 11, 3552. [Google Scholar] [CrossRef]
- Kondo, T.; Raff, M. Oligodendrocyte precursor cells reprogrammed to become multipotential CNS stem cells. Science 2000, 289, 1754–1757. [Google Scholar] [CrossRef]
- Griffiths, B.B.; Bhutani, A.; Stary, C.M. Adult neurogenesis from reprogrammed astrocytes. Neural Regen. Res. 2020, 15, 973–979. [Google Scholar] [CrossRef]
- Peng, Z.; Lu, H.; Yang, Q.; Xie, Q. Astrocyte Reprogramming in Stroke: Opportunities and Challenges. Front. Aging Neurosci. 2022, 14, 885707. [Google Scholar] [CrossRef]
- Chen, C.; Zhong, X.; Smith, D.K.; Tai, W.; Yang, J.; Zou, Y.; Wang, L.L.; Sun, J.; Qin, S.; Zhang, C.L. Astrocyte-Specific Deletion of Sox2 Promotes Functional Recovery After Traumatic Brain Injury. Cereb. Cortex 2019, 29, 54–69. [Google Scholar] [CrossRef]
- Aravantinou-Fatorou, K.; Vejdani, S.; Thomaidou, D. Cend1 and Neurog2 efficiently reprogram human cortical astrocytes to neural precursor cells and induced-neurons. Int. J. Dev. Biol. 2021, 66, 199–209. [Google Scholar] [CrossRef]
- Liu, M.-H.; Li, W.; Zheng, J.-J.; Xu, Y.-G.; He, Q.; Chen, G. Differential neuronal reprogramming induced by NeuroD1 from astrocytes in grey matter versus white matter. Neural Regen. Res. 2020, 15, 342–351. [Google Scholar] [CrossRef]
- Fujii, Y.; Arima, M.; Shimokawa, S.; Murakami, Y.; Sonoda, K.-h. The direct reprogramming of retinal astrocytes into neurons with small-molecule compounds. Investig. Ophthalmol. Vis. Sci. 2019, 60, 2. [Google Scholar]
- Bocchi, R.; Masserdotti, G.; Götz, M. Direct neuronal reprogramming: Fast forward from new concepts toward therapeutic approaches. Neuron 2022, 110, 366–393. [Google Scholar] [CrossRef]
- VandenBosch, L.S.; Wohl, S.G.; Wilken, M.S.; Hooper, M.; Finkbeiner, C.; Cox, K.; Chipman, L.; Reh, T.A. Developmental changes in the accessible chromatin, transcriptome and Ascl1-binding correlate with the loss in Müller Glial regenerative potential. Sci. Rep. 2020, 10, 13615. [Google Scholar] [CrossRef] [PubMed]
- Ali, F.R.; Cheng, K.; Kirwan, P.; Metcalfe, S.; Livesey, F.J.; Barker, R.A.; Philpott, A. The phosphorylation status of Ascl1 is a key determinant of neuronal differentiation and maturation in vivo and in vitro. Development 2014, 141, 2216–2224. [Google Scholar] [CrossRef]
- Smith, D.K.; Yang, J.; Liu, M.L.; Zhang, C.L. Small Molecules Modulate Chromatin Accessibility to Promote NEUROG2-Mediated Fibroblast-to-Neuron Reprogramming. Stem Cell Rep. 2016, 7, 955–969. [Google Scholar] [CrossRef]
- Walther, C.; Gruss, P. Pax-6, a murine paired box gene, is expressed in the developing CNS. Development 1991, 113, 1435–1449. [Google Scholar] [CrossRef] [PubMed]
- Georgala, P.A.; Carr, C.B.; Price, D.J. The role of Pax6 in forebrain development. Dev. Neurobiol. 2011, 71, 690–709. [Google Scholar] [CrossRef] [PubMed]
- Chouchane, M.; Costa, M.R. Instructing neuronal identity during CNS development and astroglial-lineage reprogramming: Roles of NEUROG2 and ASCL1. Brain Res. 2019, 1705, 66–74. [Google Scholar] [CrossRef]
- Todd, L.; Hooper, M.J.; Haugan, A.K.; Finkbeiner, C.; Jorstad, N.; Radulovich, N.; Wong, C.K.; Donaldson, P.C.; Jenkins, W.; Chen, Q.; et al. Efficient stimulation of retinal regeneration from Muller glia in adult mice using combinations of proneural bHLH transcription factors. Cell Rep. 2021, 37, 109857. [Google Scholar] [CrossRef]
- Victor, M.B.; Richner, M.; Hermanstyne, T.O.; Ransdell, J.L.; Sobieski, C.; Deng, P.Y.; Klyachko, V.A.; Nerbonne, J.M.; Yoo, A.S. Generation of human striatal neurons by microRNA-dependent direct conversion of fibroblasts. Neuron 2014, 84, 311–323. [Google Scholar] [CrossRef]
- Jin, Z.; May, W.S.; Gao, F.; Flagg, T.; Deng, X. Bcl2 suppresses DNA repair by enhancing c-Myc transcriptional activity. J. Biol. Chem. 2006, 281, 14446–14456. [Google Scholar] [CrossRef] [PubMed]
- Gresita, A.; Glavan, D.; Udristoiu, I.; Catalin, B.; Hermann, D.M.; Popa-Wagner, A. Very Low Efficiency of Direct Reprogramming of Astrocytes into Neurons in the Brains of Young and Aged Mice After Cerebral Ischemia. Front. Aging Neurosci. 2019, 11, 334. [Google Scholar] [CrossRef]
- Mattiassi, S.; Rizwan, M.; Grigsby, C.L.; Zaw, A.M.; Leong, K.W.; Yim, E.K.F. Enhanced efficiency of nonviral direct neuronal reprogramming on topographical patterns. Biomater. Sci. 2021, 9, 5175–5191. [Google Scholar] [CrossRef]
- Michaud, J.L.; Rosenquist, T.; May, N.R.; Fan, C.M. Development of neuroendocrine lineages requires the bHLH-PAS transcription factor SIM1. Genes Dev. 1998, 12, 3264–3275. [Google Scholar] [CrossRef]
- Aravantinou-Fatorou, K.; Thomaidou, D. In Vitro Direct Reprogramming of Mouse and Human Astrocytes to Induced Neurons. Methods Mol. Biol. 2020, 2155, 41–61. [Google Scholar] [CrossRef]
- Wainger, B.J.; Buttermore, E.D.; Oliveira, J.T.; Mellin, C.; Lee, S.; Saber, W.A.; Wang, A.J.; Ichida, J.K.; Chiu, I.M.; Barrett, L.; et al. Modeling pain in vitro using nociceptor neurons reprogrammed from fibroblasts. Nat. Neurosci. 2015, 18, 17–24. [Google Scholar] [CrossRef]
- Dubois, L.; Bally-Cuif, L.; Crozatier, M.; Moreau, J.; Paquereau, L.; Vincent, A. XCoe2, a transcription factor of the Col/Olf-1/EBF family involved in the specification of primary neurons in Xenopus. Curr. Biol. 1998, 8, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Arlotta, P.; Molyneaux, B.J.; Chen, J.; Inoue, J.; Kominami, R.; Macklis, J.D. Neuronal subtype-specific genes that control corticospinal motor neuron development in vivo. Neuron 2005, 45, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Aydin, B.; Sierk, M.; Moreno-Estelles, M.; Tejavibulya, L.; Kumar, N.; Flames, N.; Mahony, S.; Mazzoni, E.O. Foxa2 and Pet1 Direct and Indirect Synergy Drive Serotonergic Neuronal Differentiation. Front. Neurosci. 2022, 16, 903881. [Google Scholar] [CrossRef] [PubMed]
- Lujan, E.; Chanda, S.; Ahlenius, H.; Sudhof, T.C.; Wernig, M. Direct conversion of mouse fibroblasts to self-renewing, tripotent neural precursor cells. Proc. Natl. Acad. Sci. USA 2012, 109, 2527–2532. [Google Scholar] [CrossRef] [PubMed]
- Raciti, M.; Granzotto, M.; Duc, M.D.; Fimiani, C.; Cellot, G.; Cherubini, E.; Mallamaci, A. Reprogramming fibroblasts to neural-precursor-like cells by structured overexpression of pallial patterning genes. Mol. Cell. Neurosci. 2013, 57, 42–53. [Google Scholar] [CrossRef]
- Li, S.; Shi, Y.; Yao, X.; Wang, X.; Shen, L.; Rao, Z.; Yuan, J.; Liu, Y.; Zhou, Z.; Zhang, Z.; et al. Conversion of Astrocytes and Fibroblasts into Functional Noradrenergic Neurons. Cell Rep. 2019, 28, 682–697.e687. [Google Scholar] [CrossRef]
- Apostolova, G.; Dechant, G. Development of neurotransmitter phenotypes in sympathetic neurons. Auton. Neurosci. 2009, 151, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Buganim, Y.; Itskovich, E.; Hu, Y.C.; Cheng, A.W.; Ganz, K.; Sarkar, S.; Fu, D.; Welstead, G.G.; Page, D.C.; Jaenisch, R. Direct reprogramming of fibroblasts into embryonic Sertoli-like cells by defined factors. Cell Stem Cell 2012, 11, 373–386. [Google Scholar] [CrossRef]
- Schmidt, M.; Lin, S.; Pape, M.; Ernsberger, U.; Stanke, M.; Kobayashi, K.; Howard, M.J.; Rohrer, H. The bHLH transcription factor Hand2 is essential for the maintenance of noradrenergic properties in differentiated sympathetic neurons. Dev. Biol. 2009, 329, 191–200. [Google Scholar] [CrossRef]
- Triplett, J.W.; Wei, W.; Gonzalez, C.; Sweeney, N.T.; Huberman, A.D.; Feller, M.B.; Feldheim, D.A. Dendritic and axonal targeting patterns of a genetically-specified class of retinal ganglion cells that participate in image-forming circuits. Neural Dev. 2014, 9, 2. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, R.; Fausett, B.V.; Goldman, D. Ascl1a regulates Müller glia dedifferentiation and retinal regeneration through a Lin-28-dependent, let-7 microRNA signalling pathway. Nat. Cell Biol. 2010, 12, 1101–1107. [Google Scholar] [CrossRef]
- Batta, K.; Florkowska, M.; Kouskoff, V.; Lacaud, G. Direct reprogramming of murine fibroblasts to hematopoietic progenitor cells. Cell Rep. 2014, 9, 1871–1884. [Google Scholar] [CrossRef]
- Wilson, N.K.; Foster, S.D.; Wang, X.; Knezevic, K.; Schütte, J.; Kaimakis, P.; Chilarska, P.M.; Kinston, S.; Ouwehand, W.H.; Dzierzak, E.; et al. Combinatorial transcriptional control in blood stem/progenitor cells: Genome-wide analysis of ten major transcriptional regulators. Cell Stem Cell 2010, 7, 532–544. [Google Scholar] [CrossRef]
- Baek, S.; Oh, J.; Song, J.; Choi, H.; Yoo, J.; Park, G.Y.; Han, J.; Chang, Y.; Park, H.; Kim, H.; et al. Generation of Integration-Free Induced Neurons Using Graphene Oxide-Polyethylenimine. Small 2017, 13, 201601993. [Google Scholar] [CrossRef] [PubMed]
- Romm, E.; Nielsen, J.A.; Kim, J.G.; Hudson, L.D. Myt1 family recruits histone deacetylase to regulate neural transcription. J. Neurochem. 2005, 93, 1444–1453. [Google Scholar] [CrossRef] [PubMed]
- Corti, S.; Nizzardo, M.; Simone, C.; Falcone, M.; Donadoni, C.; Salani, S.; Rizzo, F.; Nardini, M.; Riboldi, G.; Magri, F.; et al. Direct reprogramming of human astrocytes into neural stem cells and neurons. Exp. Cell Res. 2012, 318, 1528–1541. [Google Scholar] [CrossRef]
- Guo, Z.; Zhang, L.; Wu, Z.; Chen, Y.; Wang, F.; Chen, G. In vivo direct reprogramming of reactive glial cells into functional neurons after brain injury and in an Alzheimer’s disease model. Cell Stem Cell 2014, 14, 188–202. [Google Scholar] [CrossRef]
- Pennesi, M.E.; Cho, J.H.; Yang, Z.; Wu, S.H.; Zhang, J.; Wu, S.M.; Tsai, M.J. BETA2/NeuroD1 null mice: A new model for transcription factor-dependent photoreceptor degeneration. J. Neurosci. 2003, 23, 453–461. [Google Scholar] [CrossRef]
- Bavamian, S.; Mellios, N.; Lalonde, J.; Fass, D.M.; Wang, J.; Sheridan, S.D.; Madison, J.M.; Zhou, F.; Rueckert, E.H.; Barker, D.; et al. Dysregulation of miR-34a links neuronal development to genetic risk factors for bipolar disorder. Mol. Psychiatry 2015, 20, 573–584. [Google Scholar] [CrossRef]
- Yoo, A.S.; Sun, A.X.; Li, L.; Shcheglovitov, A.; Portmann, T.; Li, Y.; Lee-Messer, C.; Dolmetsch, R.E.; Tsien, R.W.; Crabtree, G.R. MicroRNA-mediated conversion of human fibroblasts to neurons. Nature 2011, 476, 228–231. [Google Scholar] [CrossRef]
- Olson, J.M.; Asakura, A.; Snider, L.; Hawkes, R.; Strand, A.; Stoeck, J.; Hallahan, A.; Pritchard, J.; Tapscott, S.J. NeuroD2 is necessary for development and survival of central nervous system neurons. Dev. Biol. 2001, 234, 174–187. [Google Scholar] [CrossRef]
- Rodríguez-Traver, E.; Solís, O.; Díaz-Guerra, E.; Ortiz, Ó.; Vergaño-Vera, E.; Méndez-Gómez, H.R.; García-Sanz, P.; Moratalla, R.; Vicario-Abejón, C. Role of Nurr1 in the Generation and Differentiation of Dopaminergic Neurons from Stem Cells. Neurotox. Res. 2016, 30, 14–31. [Google Scholar] [CrossRef] [PubMed]
- Perlmann, T.; Wallén-Mackenzie, A. Nurr1, an orphan nuclear receptor with essential functions in developing dopamine cells. Cell Tissue Res. 2004, 318, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Boyer, L.A.; Lee, T.I.; Cole, M.F.; Johnstone, S.E.; Levine, S.S.; Zucker, J.P.; Guenther, M.G.; Kumar, R.M.; Murray, H.L.; Jenner, R.G.; et al. Core transcriptional regulatory circuitry in human embryonic stem cells. Cell 2005, 122, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Hack, M.A.; Sugimori, M.; Lundberg, C.; Nakafuku, M.; Götz, M. Regionalization and fate specification in neurospheres: The role of Olig2 and Pax6. Mol. Cell. Neurosci. 2004, 25, 664–678. [Google Scholar] [CrossRef]
- Heins, N.; Malatesta, P.; Cecconi, F.; Nakafuku, M.; Tucker, K.L.; Hack, M.A.; Chapouton, P.; Barde, Y.A.; Gotz, M. Glial cells generate neurons: The role of the transcription factor Pax6. Nat. Neurosci. 2002, 5, 308–315. [Google Scholar] [CrossRef]
- Panman, L.; Andersson, E.; Alekseenko, Z.; Hedlund, E.; Kee, N.; Mong, J.; Uhde, C.W.; Deng, Q.; Sandberg, R.; Stanton, L.W.; et al. Transcription factor-induced lineage selection of stem-cell-derived neural progenitor cells. Cell Stem Cell 2011, 8, 663–675. [Google Scholar] [CrossRef]
- Jin, K.; Zou, M.; Xiao, D.; Xiang, M. Reprogramming Fibroblasts to Neural Stem Cells by Overexpression of the Transcription Factor Ptf1a. Methods Mol. Biol. 2020, 2117, 245–263. [Google Scholar] [CrossRef]
- Mu, L.F.; Berti, L.; Masserdotti, G.; Covic, M.; Michaelidis, T.M.; Doberauer, K.; Merz, K.; Rehfeld, F.; Haslinger, A.; Wegner, M.; et al. SoxC Transcription Factors Are Required for Neuronal Differentiation in Adult Hippocampal Neurogenesis. J. Neurosci. 2012, 32, 3067–3080. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, L.; Huang, W.; Su, H.; Xue, Y.; Su, Z.; Liao, B.; Wang, H.; Bao, X.; Qin, D.; et al. Generation of integration-free neural progenitor cells from cells in human urine. Nat. Methods 2013, 10, 84–89. [Google Scholar] [CrossRef]
- Cheng, L.; Arata, A.; Mizuguchi, R.; Qian, Y.; Karunaratne, A.; Gray, P.A.; Arata, S.; Shirasawa, S.; Bouchard, M.; Luo, P.; et al. Tlx3 and Tlx1 are post-mitotic selector genes determining glutamatergic over GABAergic cell fates. Nat. Neurosci. 2004, 7, 510–517. [Google Scholar] [CrossRef]
- Yan, L.; Li, Y.; Shi, Z.; Lu, X.; Ma, J.; Hu, B.; Jiao, J.; Wang, H. The zinc finger E-box-binding homeobox 1 (Zeb1) promotes the conversion of mouse fibroblasts into functional neurons. J. Biol. Chem. 2017, 292, 12959–12970. [Google Scholar] [CrossRef]
- Damberg, M. Transcription factor AP-2 and monoaminergic functions in the central nervous system. J. Neural Transm. 2005, 112, 1281–1296. [Google Scholar] [CrossRef]
- Bertrand, N.; Castro, D.S.; Guillemot, F. Proneural genes and the specification of neural cell types. Nat. Rev. Neurosci. 2002, 3, 517–530. [Google Scholar] [CrossRef]
- Liu, F.; Zhang, Y.; Chen, F.; Yuan, J.; Li, S.; Han, S.; Lu, D.; Geng, J.; Rao, Z.; Sun, L.; et al. Neurog2 directly converts astrocytes into functional neurons in midbrain and spinal cord. Cell Death Dis. 2021, 12, 225. [Google Scholar] [CrossRef]
- Mattugini, N.; Bocchi, R.; Scheuss, V.; Russo, G.L.; Torper, O.; Lao, C.L.; Gotz, M. Inducing Different Neuronal Subtypes from Astrocytes in the Injured Mouse Cerebral Cortex. Neuron 2019, 103, 1086–1095. [Google Scholar] [CrossRef]
- Kronenberg, G.; Gertz, K.; Cheung, G.; Buffo, A.; Kettenmann, H.; Götz, M.; Endres, M. Modulation of fate determinants Olig2 and Pax6 in resident glia evokes spiking neuroblasts in a model of mild brain ischemia. Stroke 2010, 41, 2944–2949. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Efe, J.A.; Zhu, S.; Talantova, M.; Yuan, X.; Wang, S.; Lipton, S.A.; Zhang, K.; Ding, S. Direct reprogramming of mouse fibroblasts to neural progenitors. Proc. Natl. Acad. Sci. USA 2011, 108, 7838–7843. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Wang, Y.; Sun, L.; Ma, K.; Zheng, J.C. Reprogrammed mouse astrocytes retain a “memory” of tissue origin and possess more tendencies for neuronal differentiation than reprogrammed mouse embryonic fibroblasts. Protein Cell 2011, 2, 128–140. [Google Scholar] [CrossRef]
- Blum, R.; Heinrich, C.; Sánchez, R.; Lepier, A.; Gundelfinger, E.D.; Berninger, B.; Götz, M. Neuronal network formation from reprogrammed early postnatal rat cortical glial cells. Cereb. Cortex 2011, 21, 413–424. [Google Scholar] [CrossRef]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Meza-Sosa, K.F.; Valle-Garcia, D.; Pedraza-Alva, G.; Perez-Martinez, L. Role of microRNAs in central nervous system development and pathology. J. Neurosci. Res. 2012, 90, 1–12. [Google Scholar] [CrossRef]
- Zhao, Y.; Srivastava, D. A developmental view of microRNA function. Trends Biochem. Sci. 2007, 32, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jin, P. Roles of small regulatory RNAs in determining neuronal identity. Nat. Rev. Neurosci. 2010, 11, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Visvanathan, J.; Lee, S.; Lee, B.; Lee, J.W.; Lee, S.K. The microRNA miR-124 antagonizes the anti-neural REST/SCP1 pathway during embryonic CNS development. Genes Dev. 2007, 21, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Ouyang, K.; Huang, J.; Zhou, Y.; Ouyang, H.; Li, H.; Wang, G.; Wu, Q.; Wei, C.; Bi, Y.; et al. Direct conversion of fibroblasts to neurons by reprogramming PTB-regulated microRNA circuits. Cell 2013, 152, 82–96. [Google Scholar] [CrossRef] [PubMed]
- Wohl, S.G.; Reh, T.A. miR-124-9-9* potentiates Ascl1-induced reprogramming of cultured Müller glia. Glia 2016, 64, 743–762. [Google Scholar] [CrossRef] [PubMed]
- Hou, P.; Li, Y.; Zhang, X.; Liu, C.; Guan, J.; Li, H.; Zhao, T.; Ye, J.; Yang, W.; Liu, K.; et al. Pluripotent stem cells induced from mouse somatic cells by small-molecule compounds. Science 2013, 341, 651–654. [Google Scholar] [CrossRef]
- Quist, E.; Trovato, F.; Avaliani, N.; Zetterdahl, O.G.; Gonzalez-Ramos, A.; Hansen, M.G.; Kokaia, M.; Canals, I.; Ahlenius, H. Transcription factor-based direct conversion of human fibroblasts to functional astrocytes. Stem Cell Rep. 2022, 17, 1620–1635. [Google Scholar] [CrossRef]
- Lee, C.; Robinson, M.; Willerth, S.M. Direct Reprogramming of Glioblastoma Cells into Neurons Using Small Molecules. ACS Chem. Neurosci. 2018, 9, 3175–3185. [Google Scholar] [CrossRef]
- Yang, Z.; Xu, X.; Gu, C.; Nielsen, A.V.; Chen, G.; Guo, F.; Tang, C.; Zhao, Y. Chemical Pretreatment Activated a Plastic State Amenable to Direct Lineage Reprogramming. Front. Cell Dev. Biol. 2022, 10, 865038. [Google Scholar] [CrossRef]
- Ruetz, T.; Pfisterer, U.; Di Stefano, B.; Ashmore, J.; Beniazza, M.; Tian, T.V.; Kaemena, D.F.; Tosti, L.; Tan, W.; Manning, J.R.; et al. Constitutively Active SMAD2/3 Are Broad-Scope Potentiators of Transcription-Factor-Mediated Cellular Reprogramming. Cell Stem Cell 2017, 21, 791–805.e799. [Google Scholar] [CrossRef]
- Liu, C.; Hu, X.; Li, Y.; Lu, W.; Li, W.; Cao, N.; Zhu, S.; Cheng, J.; Ding, S.; Zhang, M. Conversion of mouse fibroblasts into oligodendrocyte progenitor-like cells through a chemical approach. J. Mol. Cell Biol. 2019, 11, 489–495. [Google Scholar] [CrossRef]
- Nakano, R.; Kitanaka, T.; Namba, S.; Kitanaka, N.; Sato, M.; Shibukawa, Y.; Masuhiro, Y.; Kano, K.; Matsumoto, T.; Sugiya, H. All-trans retinoic acid induces reprogramming of canine dedifferentiated cells into neuron-like cells. PLoS ONE 2020, 15, e0229892. [Google Scholar] [CrossRef] [PubMed]
- Schweinfurth, N.; Hohmann, S.; Deuschle, M.; Lederbogen, F.; Schloss, P. Valproic acid and all trans retinoic acid differentially induce megakaryopoiesis and platelet-like particle formation from the megakaryoblastic cell line MEG-01. Platelets 2010, 21, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhao, T.; Guan, J.; Zhang, X.; Fu, Y.; Ye, J.; Zhu, J.; Meng, G.; Ge, J.; Yang, S.; et al. A XEN-like State Bridges Somatic Cells to Pluripotency during Chemical Reprogramming. Cell 2015, 163, 1678–1691. [Google Scholar] [CrossRef]
- Kogo, Y.; Seto, C.; Totani, Y.; Mochizuki, M.; Nakahara, T.; Oka, K.; Yoshioka, T.; Ito, E. Rapid differentiation of human dental pulp stem cells to neuron-like cells by high K(+) stimulation. Biophys. Physicobiol. 2020, 17, 132–139. [Google Scholar] [CrossRef] [PubMed]
- He, Z.Q.; Li, Y.H.; Feng, G.H.; Yuan, X.W.; Lu, Z.B.; Dai, M.; Hu, Y.P.; Zhang, Y.; Zhou, Q.; Li, W. Pharmacological Perturbation of Mechanical Contractility Enables Robust Transdifferentiation of Human Fibroblasts into Neurons. Adv. Sci. 2022, 9, e2104682. [Google Scholar] [CrossRef]
- Lee, T.H.; Liu, P.S.; Wang, S.J.; Tsai, M.M.; Shanmugam, V.; Hsieh, H.L. Bradykinin, as a Reprogramming Factor, Induces Transdifferentiation of Brain Astrocytes into Neuron-like Cells. Biomedicines 2021, 9, 923. [Google Scholar] [CrossRef]
- Li, X.; Liu, D.; Ma, Y.; Du, X.; Jing, J.; Wang, L.; Xie, B.; Sun, D.; Sun, S.; Jin, X.; et al. Direct Reprogramming of Fibroblasts via a Chemically Induced XEN-like State. Cell Stem Cell 2017, 21, 264–273.e267. [Google Scholar] [CrossRef]
- Solomon, E.; Davis-Anderson, K.; Hovde, B.; Micheva-Viteva, S.; Harris, J.F.; Twary, S.; Iyer, R. Global transcriptome profile of the developmental principles of in vitro iPSC-to-motor neuron differentiation. BMC Mol. Cell Biol. 2021, 22, 13. [Google Scholar] [CrossRef]
- Yin, J.C.; Zhang, L.; Ma, N.X.; Wang, Y.; Lee, G.; Hou, X.Y.; Lei, Z.F.; Zhang, F.Y.; Dong, F.P.; Wu, G.Y.; et al. Chemical Conversion of Human Fetal Astrocytes into Neurons through Modulation of Multiple Signaling Pathways. Stem Cell Rep. 2019, 12, 488–501. [Google Scholar] [CrossRef]
- Tian, E.; Sun, G.; Sun, G.; Chao, J.; Ye, P.; Warden, C.; Riggs, A.D.; Shi, Y. Small-Molecule-Based Lineage Reprogramming Creates Functional Astrocytes. Cell Rep. 2016, 16, 781–792. [Google Scholar] [CrossRef]
- Yang, C.; Chen, Z.; Yu, H.; Liu, X. Inhibition of Disruptor of Telomeric Silencing 1-Like Alleviated Renal Ischemia and Reperfusion Injury-Induced Fibrosis by Blocking PI3K/AKT-Mediated Oxidative Stress. Drug. Des. Devel. Ther. 2019, 13, 4375–4387. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Oh, J.; Huang, L.; Zeng, Z.; Duan, P.; Li, Z.; Yun, Y.; Kim, J.; Ha, Y.; Cao, K. The combination of forskolin and VPA increases gene expression efficiency to the hypoxia/neuron-specific system. Ann. Transl. Med. 2020, 8, 933. [Google Scholar] [CrossRef]
- Liu, D.; Rychkov, G.; Al-Hawwas, M.; Manaph, N.P.A.; Zhou, F.; Bobrovskaya, L.; Liao, H.; Zhou, X.F. Conversion of human urine-derived cells into neuron-like cells by small molecules. Mol. Biol. Rep. 2020, 47, 2713–2722. [Google Scholar] [CrossRef]
- Chang, J.H.; Tsai, P.H.; Wang, K.Y.; Wei, Y.T.; Chiou, S.H.; Mou, C.Y. Generation of Functional Dopaminergic Neurons from Reprogramming Fibroblasts by Nonviral-based Mesoporous Silica Nanoparticles. Sci. Rep. 2018, 8, 11. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Zhao, A.D.; Sun, M.L.; Ma, K.; Fu, X.B. Direct conversion of human fibroblasts into dopaminergic neuron-like cells using small molecules and protein factors. Mil. Med. Res. 2020, 7, 52. [Google Scholar] [CrossRef]
- Ma, N.X.; Yin, J.C.; Chen, G. Transcriptome Analysis of Small Molecule-Mediated Astrocyte-to-Neuron Reprogramming. Front. Cell Dev. Biol. 2019, 7, 82. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Gong, T.; Liu, H.; Shen, J.; Wu, B.; Jiang, Q.; Wang, Q.; Zhang, Y.; Duan, L.; Hu, J.; et al. Identification of microRNAs related with neural germ layer lineage-specific progenitors during reprogramming. J. Mol. Histol. 2022, 53, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Huang, Y.; Yu, C.; Liu, Q.; Qiu, C.; Wan, G. Cochlear Sox2(+) Glial Cells Are Potent Progenitors for Spiral Ganglion Neuron Reprogramming Induced by Small Molecules. Front. Cell Dev. Biol. 2021, 9, 728352. [Google Scholar] [CrossRef]
- Drouin-Ouellet, J.; Lau, S.; Brattås, P.L.; Rylander Ottosson, D.; Pircs, K.; Grassi, D.A.; Collins, L.M.; Vuono, R.; Andersson Sjöland, A.; Westergren-Thorsson, G.; et al. REST suppression mediates neural conversion of adult human fibroblasts via microRNA-dependent and -independent pathways. EMBO Mol. Med. 2017, 9, 1117–1131. [Google Scholar] [CrossRef]
- Seibler, P.; Graziotto, J.; Jeong, H.; Simunovic, F.; Klein, C.; Krainc, D. Mitochondrial Parkin Recruitment Is Impaired in Neurons Derived from Mutant PINK1 Induced Pluripotent Stem Cells. J. Neurosci. 2011, 31, 5970–5976. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Li, W.; Zhou, H.; Wei, W.; Ambasudhan, R.; Lin, T.; Kim, J.; Zhang, K.; Ding, S. Reprogramming of human primary somatic cells by OCT4 and chemical compounds. Cell Stem Cell 2010, 7, 651–655. [Google Scholar] [CrossRef]
- Zhao, A.D.; Qin, H.; Sun, M.L.; Ma, K.; Fu, X.B. Efficient and rapid conversion of human astrocytes and ALS mouse model spinal cord astrocytes into motor neuron-like cells by defined small molecules. Mil. Med. Res. 2020, 7, 42. [Google Scholar] [CrossRef]
- Wan, X.Y.; Xu, L.Y.; Li, B.; Sun, Q.H.; Ji, Q.L.; Huang, D.D.; Zhao, L.; Xiao, Y.T. Chemical conversion of human lung fibroblasts into neuronal cells. Int. J. Mol. Med. 2018, 41, 1463–1468. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Huang, Z.; Xu, J.; Hou, K.; Yu, Y.; Lv, S.; Chen, L.; Li, Y.; Quan, C.; Chi, G. MiR-124 and Small Molecules Synergistically Regulate the Generation of Neuronal Cells from Rat Cortical Reactive Astrocytes. Mol. Neurobiol. 2021, 58, 2447–2464. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Lin, Y.H.; Sun, Y.J.; Zhu, S.; Zheng, J.; Liu, K.; Cao, N.; Li, K.; Huang, Y.; Ding, S. Pharmacological Reprogramming of Fibroblasts into Neural Stem Cells by Signaling-Directed Transcriptional Activation. Cell Stem Cell 2016, 18, 653–667. [Google Scholar] [CrossRef]
- Kirchenwitz, M.; Stahnke, S.; Grunau, K.; Melcher, L.; van Ham, M.; Rottner, K.; Steffen, A.; Stradal, T.E.B. The autophagy inducer SMER28 attenuates microtubule dynamics mediating neuroprotection. Sci. Rep. 2022, 12, 17805. [Google Scholar] [CrossRef]
- Fernandes, G.S.; Singh, R.D.; Kim, K.K. Generation of a Pure Culture of Neuron-like Cells with a Glutamatergic Phenotype from Mouse Astrocytes. Biomedicines 2022, 10, 928. [Google Scholar] [CrossRef]
- Wang, J.; Lu, J.; Bond, M.C.; Chen, M.; Ren, X.R.; Lyerly, H.K.; Barak, L.S.; Chen, W. Identification of select glucocorticoids as Smoothened agonists: Potential utility for regenerative medicine. Proc. Natl. Acad. Sci. USA 2010, 107, 9323–9328. [Google Scholar] [CrossRef]
- Ye, W.; Shimamura, K.; Rubenstein, J.L.; Hynes, M.A.; Rosenthal, A. FGF and Shh signals control dopaminergic and serotonergic cell fate in the anterior neural plate. Cell 1998, 93, 755–766. [Google Scholar] [CrossRef]
- Rhee, Y.H.; Ko, J.Y.; Chang, M.Y.; Yi, S.H.; Kim, D.; Kim, C.H.; Shim, J.W.; Jo, A.Y.; Kim, B.W.; Lee, H.; et al. Protein-based human iPS cells efficiently generate functional dopamine neurons and can treat a rat model of Parkinson disease. J. Clin. Investig. 2011, 121, 2326–2335. [Google Scholar] [CrossRef]
- Sotthibundhu, A.; Nopparat, C.; Natphopsuk, S.; Phuthong, S.; Noisa, P.; Govitrapong, P. Combination of Melatonin and Small Molecules Improved Reprogramming Neural Cell Fates via Autophagy Activation. Neurochem. Res. 2022, 47, 2580–2590. [Google Scholar] [CrossRef]
- Roussa, E.; Oehlke, O.; Rahhal, B.; Heermann, S.; Heidrich, S.; Wiehle, M.; Krieglstein, K. Transforming growth factor beta cooperates with persephin for dopaminergic phenotype induction. Stem. Cells. 2008, 26, 1683–1694. [Google Scholar] [CrossRef]
- Zhang, L.; Yin, J.-C.; Yeh, H.; Ma, N.-X.; Lee, G.; Chen, X.A.; Wang, Y.; Lin, L.; Chen, L.; Jin, P.; et al. Small Molecules Efficiently Reprogram Human Astroglial Cells into Functional Neurons. Cell Stem Cell 2015, 17, 735–747. [Google Scholar] [CrossRef]
- Alari, V.; Scalmani, P.; Ajmone, P.F.; Perego, S.; Avignone, S.; Catusi, I.; Lonati, P.A.; Borghi, M.O.; Finelli, P.; Terragni, B.; et al. Histone Deacetylase Inhibitors Ameliorate Morphological Defects and Hypoexcitability of iPSC-Neurons from Rubinstein-Taybi Patients. Int. J. Mol. Sci. 2021, 22, 5777. [Google Scholar] [CrossRef]
- Kanai, H.; Sawa, A.; Chen, R.W.; Leeds, P.; Chuang, D.M. Valproic acid inhibits histone deacetylase activity and suppresses excitotoxicity-induced GAPDH nuclear accumulation and apoptotic death in neurons. Pharmacogenom. J. 2004, 4, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Klemm, S.L.; Shipony, Z.; Greenleaf, W.J. Chromatin accessibility and the regulatory epigenome. Nat. Rev. Genet. 2019, 20, 207–220. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Chen, J.; Zhang, Y.; Zhang, M.; Yang, X.; Li, Y.; Sun, H.; Lin, L.; Fan, K.; Liang, L.; et al. Sequential EMT-MET induces neuronal conversion through Sox2. Cell Discov. 2017, 3, 17017. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Cai, Y.; Wang, Y.; Nie, Y.; Zhang, C.; Xu, Y.; Zhang, X.; Lu, Y.; Wang, Z.; Poo, M.; et al. Cloning of Macaque Monkeys by Somatic Cell Nuclear Transfer. Cell 2018, 172, 881–887.e887. [Google Scholar] [CrossRef] [PubMed]
- Willadsen, S.M. Nuclear transplantation in sheep embryos. Nature 1986, 320, 63–65. [Google Scholar] [CrossRef]
- Luo, C.; Lee, Q.Y.; Wapinski, O.; Castanon, R.; Nery, J.R.; Mall, M.; Kareta, M.S.; Cullen, S.M.; Goodell, M.A.; Chang, H.Y.; et al. Global DNA methylation remodeling during direct reprogramming of fibroblasts to neurons. eLife 2019, 8, e40197. [Google Scholar] [CrossRef]
- Yang, D.W.; Moon, J.S.; Ko, H.M.; Shin, Y.K.; Fukumoto, S.; Kim, S.H.; Kim, M.S. Direct reprogramming of fibroblasts into diverse lineage cells by DNA demethylation followed by differentiating cultures. Korean J. Physiol. Pharmacol. 2020, 24, 463–472. [Google Scholar] [CrossRef]
- Chung, Y.G.; Matoba, S.; Liu, Y.; Eum, J.H.; Lu, F.; Jiang, W.; Lee, J.E.; Sepilian, V.; Cha, K.Y.; Lee, D.R.; et al. Histone Demethylase Expression Enhances Human Somatic Cell Nuclear Transfer Efficiency and Promotes Derivation of Pluripotent Stem Cells. Cell Stem Cell 2015, 17, 758–766. [Google Scholar] [CrossRef] [PubMed]
- Patel, T.; Hobert, O. Coordinated control of terminal differentiation and restriction of cellular plasticity. eLife 2017, 6, e24100. [Google Scholar] [CrossRef] [PubMed]
- Treutlein, B.; Lee, Q.Y.; Camp, J.G.; Mall, M.; Koh, W.; Shariati, S.A.; Sim, S.; Neff, N.F.; Skotheim, J.M.; Wernig, M.; et al. Dissecting direct reprogramming from fibroblast to neuron using single-cell RNA-seq. Nature 2016, 534, 391–395. [Google Scholar] [CrossRef]
- Ma, N.X.; Puls, B.; Chen, G. Transcriptomic analyses of NeuroD1-mediated astrocyte-to-neuron conversion. Dev. Neurobiol. 2022, 82, 375–391. [Google Scholar] [CrossRef]
- Wang, L.L.; Su, Z.; Tai, W.; Zou, Y.; Xu, X.M.; Zhang, C.L. The p53 Pathway Controls SOX2-Mediated Reprogramming in the Adult Mouse Spinal Cord. Cell Rep. 2016, 17, 891–903. [Google Scholar] [CrossRef]
- McKay, N.D.; Robinson, B.; Brodie, R.; Rooke-Allen, N. Glucose transport and metabolism in cultured human skin fibroblasts. Biochim. Biophys. Acta 1983, 762, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Tsacopoulos, M.; Magistretti, P.J. Metabolic coupling between glia and neurons. J. Neurosci. 1996, 16, 877–885. [Google Scholar] [CrossRef]
- Liu, H.; He, Z.; April, S.L.; Trefny, M.P.; Rougier, J.S.; Salemi, S.; Olariu, R.; Widmer, H.R.; Simon, H.U. Biochemical re-programming of human dermal stem cells to neurons by increasing mitochondrial membrane potential. Cell Death Differ. 2019, 26, 1048–1061. [Google Scholar] [CrossRef]
- Riechers, S.P.; Mojsilovic-Petrovic, J.; Belton, T.B.; Chakrabarty, R.P.; Garjani, M.; Medvedeva, V.; Dalton, C.; Wong, Y.C.; Chandel, N.S.; Dienel, G.; et al. Neurons undergo pathogenic metabolic reprogramming in models of familial ALS. Mol. Metab. 2022, 60, 101468. [Google Scholar] [CrossRef]
- Moutaoufik, M.T.; Malty, R.; Amin, S.; Zhang, Q.; Phanse, S.; Gagarinova, A.; Zilocchi, M.; Hoell, L.; Minic, Z.; Gagarinova, M.; et al. Rewiring of the Human Mitochondrial Interactome during Neuronal Reprogramming Reveals Regulators of the Respirasome and Neurogenesis. iScience 2019, 19, 1114–1132. [Google Scholar] [CrossRef]
- Hämäläinen, R.H.; Manninen, T.; Koivumäki, H.; Kislin, M.; Otonkoski, T.; Suomalainen, A. Tissue- and cell-type-specific manifestations of heteroplasmic mtDNA 3243A>G mutation in human induced pluripotent stem cell-derived disease model. Proc. Natl. Acad. Sci. USA 2013, 110, E3622–E3630. [Google Scholar] [CrossRef]
- Liu, Y.; Lopez-Santiago, L.F.; Yuan, Y.; Jones, J.M.; Zhang, H.; O’Malley, H.A.; Patino, G.A.; O’Brien, J.E.; Rusconi, R.; Gupta, A.; et al. Dravet syndrome patient-derived neurons suggest a novel epilepsy mechanism. Ann. Neurol. 2013, 74, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Chanda, S.; Marro, S.; Wernig, M.; Sudhof, T.C. Neurons generated by direct conversion of fibroblasts reproduce synaptic phenotype caused by autism-associated neuroligin-3 mutation. Proc. Natl. Acad. Sci. USA 2013, 110, 16622–16627. [Google Scholar] [CrossRef] [PubMed]
- Marchetto, M.C.; Belinson, H.; Tian, Y.; Freitas, B.C.; Fu, C.; Vadodaria, K.; Beltrao-Braga, P.; Trujillo, C.A.; Mendes, A.P.D.; Padmanabhan, K.; et al. Altered proliferation and networks in neural cells derived from idiopathic autistic individuals. Mol. Psychiatry 2017, 22, 820–835. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Liu, M.L.; Zang, T.; Zhang, C.L. Direct Reprogramming Rather than iPSC-Based Reprogramming Maintains Aging Hallmarks in Human Motor Neurons. Front. Mol. Neurosci. 2017, 10, 359. [Google Scholar] [CrossRef] [PubMed]
- Kumamaru, H.; Kadoya, K.; Adler, A.F.; Takashima, Y.; Graham, L.; Coppola, G.; Tuszynski, M.H. Generation and post-injury integration of human spinal cord neural stem cells. Nat. Methods 2018, 15, 723–731. [Google Scholar] [CrossRef]
- Kenvin, S.; Torregrosa-Muñumer, R.; Reidelbach, M.; Pennonen, J.; Turkia, J.J.; Rannila, E.; Kvist, J.; Sainio, M.T.; Huber, N.; Herukka, S.K.; et al. Threshold of heteroplasmic truncating MT-ATP6 mutation in reprogramming, Notch hyperactivation and motor neuron metabolism. Hum. Mol. Genet. 2022, 31, 958–974. [Google Scholar] [CrossRef]
- Doers, M.E.; Musser, M.T.; Nichol, R.; Berndt, E.R.; Baker, M.; Gomez, T.M.; Zhang, S.C.; Abbeduto, L.; Bhattacharyya, A. iPSC-derived forebrain neurons from FXS individuals show defects in initial neurite outgrowth. Stem Cells Dev. 2014, 23, 1777–1787. [Google Scholar] [CrossRef]
- Begentas, O.C.; Koc, D.; Kiris, E. Establishment of Human Induced Pluripotent Stem Cells from Multiple Sclerosis Patients. Methods Mol. Biol. 2022, 2549, 43–67. [Google Scholar] [CrossRef]
- Almeida, S.; Gascon, E.; Tran, H.; Chou, H.J.; Gendron, T.F.; DeGroot, S.; Tapper, A.R.; Sellier, C.; Charlet-Berguerand, N.; Karydas, A.; et al. Modeling key pathological features of frontotemporal dementia with C9ORF72 repeat expansion in iPSC-derived human neurons. Acta Neuropathol. 2013, 126, 385–399. [Google Scholar] [CrossRef] [PubMed]
- Camnasio, S.; Delli Carri, A.; Lombardo, A.; Grad, I.; Mariotti, C.; Castucci, A.; Rozell, B.; Lo Riso, P.; Castiglioni, V.; Zuccato, C.; et al. The first reported generation of several induced pluripotent stem cell lines from homozygous and heterozygous Huntington’s disease patients demonstrates mutation related enhanced lysosomal activity. Neurobiol. Dis. 2012, 46, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Brennand, K.J.; Simone, A.; Jou, J.; Gelboin-Burkhart, C.; Tran, N.; Sangar, S.; Li, Y.; Mu, Y.; Chen, G.; Yu, D.; et al. Modelling schizophrenia using human induced pluripotent stem cells. Nature 2011, 473, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.; Qin, S.; Yuan, Y.; Hu, X.; Huang, X.; Liu, H.; Pu, Y.; He, C.; Su, Z. NOTCH1 signaling regulates the latent neurogenic program in adult reactive astrocytes after spinal cord injury. Theranostics 2022, 12, 4548–4563. [Google Scholar] [CrossRef]
- Kim, K.Y.; Hysolli, E.; Park, I.H. Neuronal maturation defect in induced pluripotent stem cells from patients with Rett syndrome. Proc. Natl. Acad. Sci. USA 2011, 108, 14169–14174. [Google Scholar] [CrossRef]
- Petit, I.; Kesner, N.S.; Karry, R.; Robicsek, O.; Aberdam, E.; Müller, F.J.; Aberdam, D.; Ben-Shachar, D. Induced pluripotent stem cells from hair follicles as a cellular model for neurodevelopmental disorders. Stem Cell Res. 2012, 8, 134–140. [Google Scholar] [CrossRef]
- Gatto, N.; Dos Santos Souza, C.; Shaw, A.C.; Bell, S.M.; Myszczynska, M.A.; Powers, S.; Meyer, K.; Castelli, L.M.; Karyka, E.; Mortiboys, H.; et al. Directly converted astrocytes retain the ageing features of the donor fibroblasts and elucidate the astrocytic contribution to human CNS health and disease. Aging Cell 2021, 20, e13281. [Google Scholar] [CrossRef]
- Qin, H.; Zhao, A.; Ma, K.; Fu, X. Chemical conversion of human and mouse fibroblasts into motor neurons. Sci. China Life Sci. 2018, 61, 1151–1167. [Google Scholar] [CrossRef]
- Zhao, T.; Zhang, Z.N.; Rong, Z.; Xu, Y. Immunogenicity of induced pluripotent stem cells. Nature 2011, 474, 212–215. [Google Scholar] [CrossRef]
- Hu, K. Vectorology and factor delivery in induced pluripotent stem cell reprogramming. Stem Cells Dev. 2014, 23, 1301–1315. [Google Scholar] [CrossRef]
- Yu, J.; Hu, K.; Smuga-Otto, K.; Tian, S.; Stewart, R.; Slukvin, I.I.; Thomson, J.A. Human induced pluripotent stem cells free of vector and transgene sequences. Science 2009, 324, 797–801. [Google Scholar] [CrossRef]
- Rouhani, F.J.; Zou, X.; Danecek, P.; Badja, C.; Amarante, T.D.; Koh, G.; Wu, Q.; Memari, Y.; Durbin, R.; Martincorena, I.; et al. Substantial somatic genomic variation and selection for BCOR mutations in human induced pluripotent stem cells. Nat. Genet. 2022, 54, 1406–1416. [Google Scholar] [CrossRef]
- Tanaka, Y.; Hysolli, E.; Su, J.; Xiang, Y.; Kim, K.Y.; Zhong, M.; Li, Y.; Heydari, K.; Euskirchen, G.; Snyder, M.P.; et al. Transcriptome Signature and Regulation in Human Somatic Cell Reprogramming. Stem Cell Rep. 2015, 4, 1125–1139. [Google Scholar] [CrossRef]
- Dhanasekaran, R.; Deutzmann, A.; Mahauad-Fernandez, W.D.; Hansen, A.S.; Gouw, A.M.; Felsher, D.W. The MYC oncogene—The grand orchestrator of cancer growth and immune evasion. Nat. Rev. Clin. Oncol. 2022, 19, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, M.; Koyanagi, M.; Tanabe, K.; Takahashi, K.; Ichisaka, T.; Aoi, T.; Okita, K.; Mochiduki, Y.; Takizawa, N.; Yamanaka, S. Generation of induced pluripotent stem cells without Myc from mouse and human fibroblasts. Nat. Biotechnol. 2008, 26, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Assinck, P.; Duncan, G.J.; Hilton, B.J.; Plemel, J.R.; Tetzlaff, W. Cell transplantation therapy for spinal cord injury. Nat. Neurosci. 2017, 20, 637–647. [Google Scholar] [CrossRef]
- Rubio, A.; Luoni, M.; Giannelli, S.G.; Radice, I.; Iannielli, A.; Cancellieri, C.; Di Berardino, C.; Regalia, G.; Lazzari, G.; Menegon, A.; et al. Rapid and efficient CRISPR/Cas9 gene inactivation in human neurons during human pluripotent stem cell differentiation and direct reprogramming. Sci. Rep. 2016, 6, 37540. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.J.; Yang, F.H.; Li, W.; Wang, T.; Lin, Y.; Feng, J.; Chen, N.H.; Jiang, M.; Wang, J.H.; Hu, X.T.; et al. In vivo Neuroregeneration to Treat Ischemic Stroke Through NeuroD1 AAV-Based Gene Therapy in Adult Non-human Primates. Front. Cell Dev. Biol. 2020, 8, 590008. [Google Scholar] [CrossRef]

| Original Cell Type | Type of Reprogrammed Neuron | References |
|---|---|---|
| Fibroblasts | Neurons | [24] |
| Infrapatellar fat pad stem cells | Neurons | [34] |
| Astrocytes | Neurons | [26,35] |
| Dental stem cell | Neurons | [36] |
| Hematopoietic cells | Induced neuronal cells | [37] |
| Urine-derived cells | Induced neuronal cells | [38] |
| Olfactory ensheathing cells | Neuronal cells | [39] |
| Spermatogonial stem cells | Dopaminergic neurons | [40] |
| Glioma cells | Neurons | [41] |
| Microglia | Neurons | [42] |
| Striatal neurons/post-mitotic callosal neurons | Neurons | [43] |
| Peripheral blood T cells | Neurons | [29] |
| Peripheral blood mononuclear cells | Neurons | [30] |
| Spiral ganglion non-neuronal cells | Cochlear hair cells and cochlear nucleus neurons | [44] |
| Pericytes | Cholinergic neurons | [27] |
| Pluripotent stem cell-derived cardiomyocytes | Neurons | [45] |
| Oligodendrocytes | Functional neurons | [46] |
| NG2 cells | Neurons | [47] |
| Mesenchymal stem cells | Neural precursors | [48] |
| Hair follicle keratinocytes | Dopaminergic neurons | [32] |
| Müller glia | Neurogenic retinal progenitors | [49] |
| Adipose-derived stem cells | Neural stem cells and functional GABAergic neurons | [50] |
| Hepatocytes | Functional induced neuronal cells | [31] |
| Oligodendrocyte precursor cells | Neurons | [51] |
| Interfollicular keratinocytes | Neurons | [52] |
| Bone marrow-derived mesenchymal stem cells | GABAergic neurons | [53] |
| Disease | Applications/Results of Neuronal Reprogramming | Reference |
|---|---|---|
| Dravet syndrome | Fibroblasts derived from controls and patients were differentiated into neurons. Epilepsy-specific iPSC-derived neurons are helpful for modeling epilepsy-like hyperactivity. | [184] |
| MT-ATP6 | Skin fibroblast reprogramming and iPSCs can model disease caused by the MT-ATP6 mutation. | [189] |
| Alzheimer’s disease | Small molecules induce the reprogramming of patient fibroblasts into neurons for personalized modeling of neurological disease. | [54] |
| Fragile X syndrome | Fibroblasts from patients can be induced into iPSC lines to enable in vitro modeling of the human disease. | [190] |
| Multiple sclerosis | iPSCs from peripheral blood mononuclear cells can be used to model multiple sclerosis. | [191] |
| Glioblastoma multiforme | Isoxazole acts as a stem cell modulator to trigger neuronal gene expression and block tumor cell proliferation, which may guide research into reprogramming as an antitumor strategy. | [41] |
| Frontotemporal dementia, amyotrophic lateral sclerosis | Fibroblasts were isolated from patients’ skin to generate induced pluripotent stem cells to investigate the pathological mechanisms underlying frontotemporal dementia or amyotrophic lateral sclerosis. | [192] |
| Huntington’s disease | Stable HD-iPS cell lines have been established to investigate disease mechanisms. | [193] |
| Schizophrenia | The authors directly reprogrammed fibroblasts/hair follicle-derived cells from schizophrenia patients into iPSCs, which they differentiated into neurons. These neurons were then studied for disease pathology. | [32,194] |
| Spinal cord injury | NOTCH1 signaling regulates the latent neurogenic program in adult reactive astrocytes after spinal cord injury. | [195] |
| Amyotrophic lateral sclerosis | Peripheral blood cells from an ALS patient carrying the TARDBP p.A382T mutation were reprogrammed into iPSCs. | [30] |
| Stroke | Overexpression of Ascl1 can convert astrocytes from the subventricular zone into neurons in vivo after stroke. | [8] |
| Parkinson’s disease | Fibroblasts were taken from the pathology biopsies of Parkinson’s disease patients and encouraged to develop into dopaminergic neurons, which can be used for future studies into the mechanistic underpinnings of the disease. | [151] |
| Rett syndrome | Overexpressing reprogramming factors in Rett syndrome fibroblasts generated iPSCs, which differentiated into neurons with a neuronal maturation phenotype similar to that of the clinical syndrome. | [196] |
| Neurodevelopmental disorders | Human hair follicle-derived iPSCs can be differentiated into various neural lineages. This experimental system provides an in vitro model to study normal and pathological neural development without the need for skin biopsies. | [197] |
| Ageing | Directly converted astrocytes retain the ageing features of the donor fibroblasts and clarify the astrocytic contribution to human CNS health and disease. | [198] |
| Bipolar disorder | Human fibroblasts can be reprogrammed into induced neurons or iPSCs, then differentiated into neurons for mechanistic studies of the disease. | [97] |
| Pain | Transcription factors can transform mouse and human fibroblasts into noxious-stimulus-detecting (injury receptor) neurons, which displayed TrpV1-mediated sensitization to inflammation. | [78] |
| Demyelinating diseases | Exposing mouse embryonic fibroblasts to chemical conditions could induce their differentiation into OPC-like cells, which may serve as a therapeutic strategy for treating demyelinating diseases. | [131] |
| Mitochondrial DNA mutations | This work generated stem cells from patients carrying the most common human disease mutation in mitochondrial DNA, m.3243A>G (MELAS). | [183] |
| Autism spectrum disorders | Fibroblasts from patients can be reprogrammed into neurons with fewer excitatory synapses, a faulty neural network phenotype, or a synaptic phenotype comparable to that induced by autism-associated neuroligand protein-3 mutations, confirming the use of induced neuronal cells for disease modeling. | [185,186] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Huang, L.; Yang, Y.; Xu, W.; Qin, Q.; Qin, R.; Liang, X.; Lai, X.; Huang, X.; Xie, M.; et al. Somatic Cell Reprogramming for Nervous System Diseases: Techniques, Mechanisms, Potential Applications, and Challenges. Brain Sci. 2023, 13, 524. https://doi.org/10.3390/brainsci13030524
Chen J, Huang L, Yang Y, Xu W, Qin Q, Qin R, Liang X, Lai X, Huang X, Xie M, et al. Somatic Cell Reprogramming for Nervous System Diseases: Techniques, Mechanisms, Potential Applications, and Challenges. Brain Sciences. 2023; 13(3):524. https://doi.org/10.3390/brainsci13030524
Chicago/Turabian StyleChen, Jiafeng, Lijuan Huang, Yue Yang, Wei Xu, Qingchun Qin, Rongxing Qin, Xiaojun Liang, Xinyu Lai, Xiaoying Huang, Minshan Xie, and et al. 2023. "Somatic Cell Reprogramming for Nervous System Diseases: Techniques, Mechanisms, Potential Applications, and Challenges" Brain Sciences 13, no. 3: 524. https://doi.org/10.3390/brainsci13030524
APA StyleChen, J., Huang, L., Yang, Y., Xu, W., Qin, Q., Qin, R., Liang, X., Lai, X., Huang, X., Xie, M., & Chen, L. (2023). Somatic Cell Reprogramming for Nervous System Diseases: Techniques, Mechanisms, Potential Applications, and Challenges. Brain Sciences, 13(3), 524. https://doi.org/10.3390/brainsci13030524