Natural Products as New Treatment Options for Trichomoniasis: A Molecular Docking Investigation




Abstract
:1. Introduction
1.1. Trichomonas vaginalis Protein Targets
1.2. Homology Modeling
1.3. Molecular Docking
2. Materials and Methods
2.1. Homology Modeling
2.2. Molecular Docking
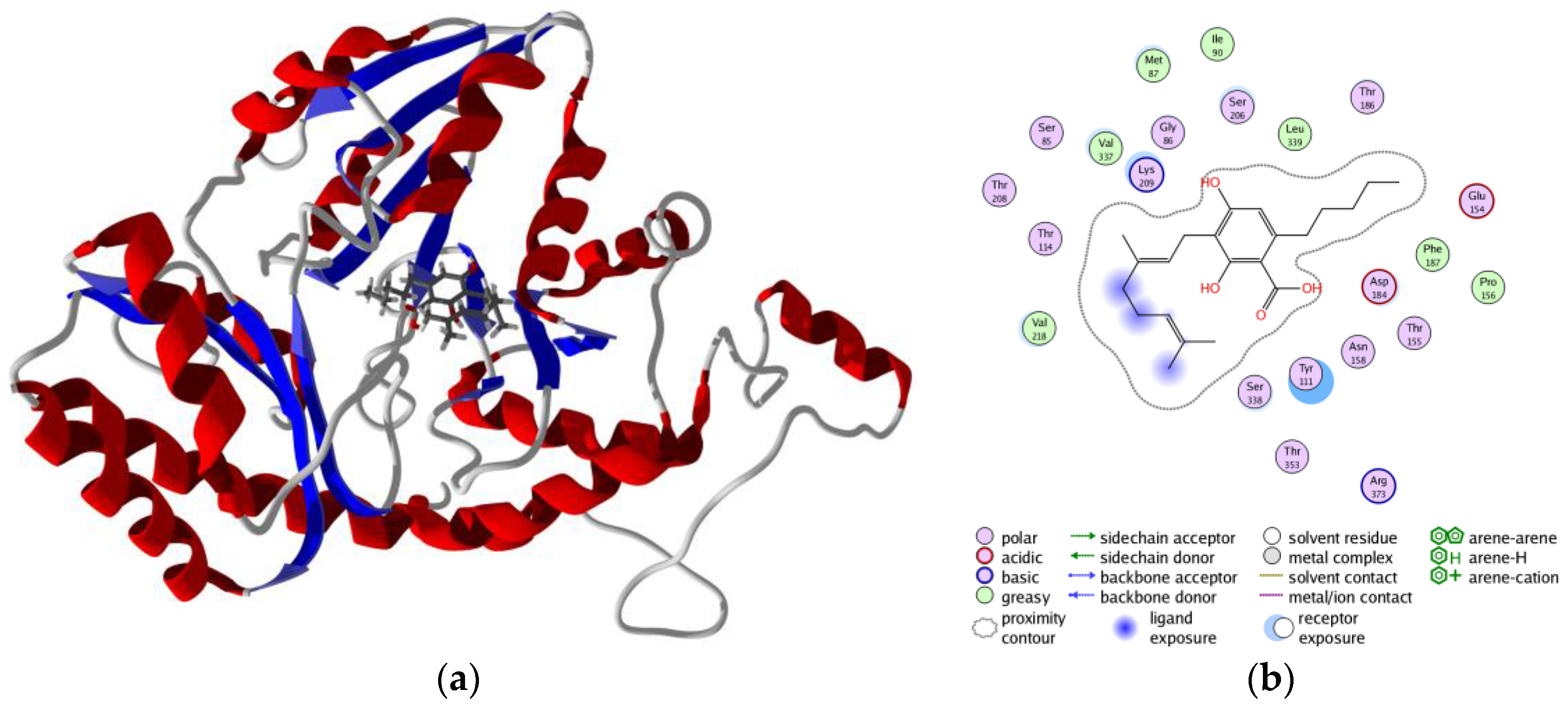
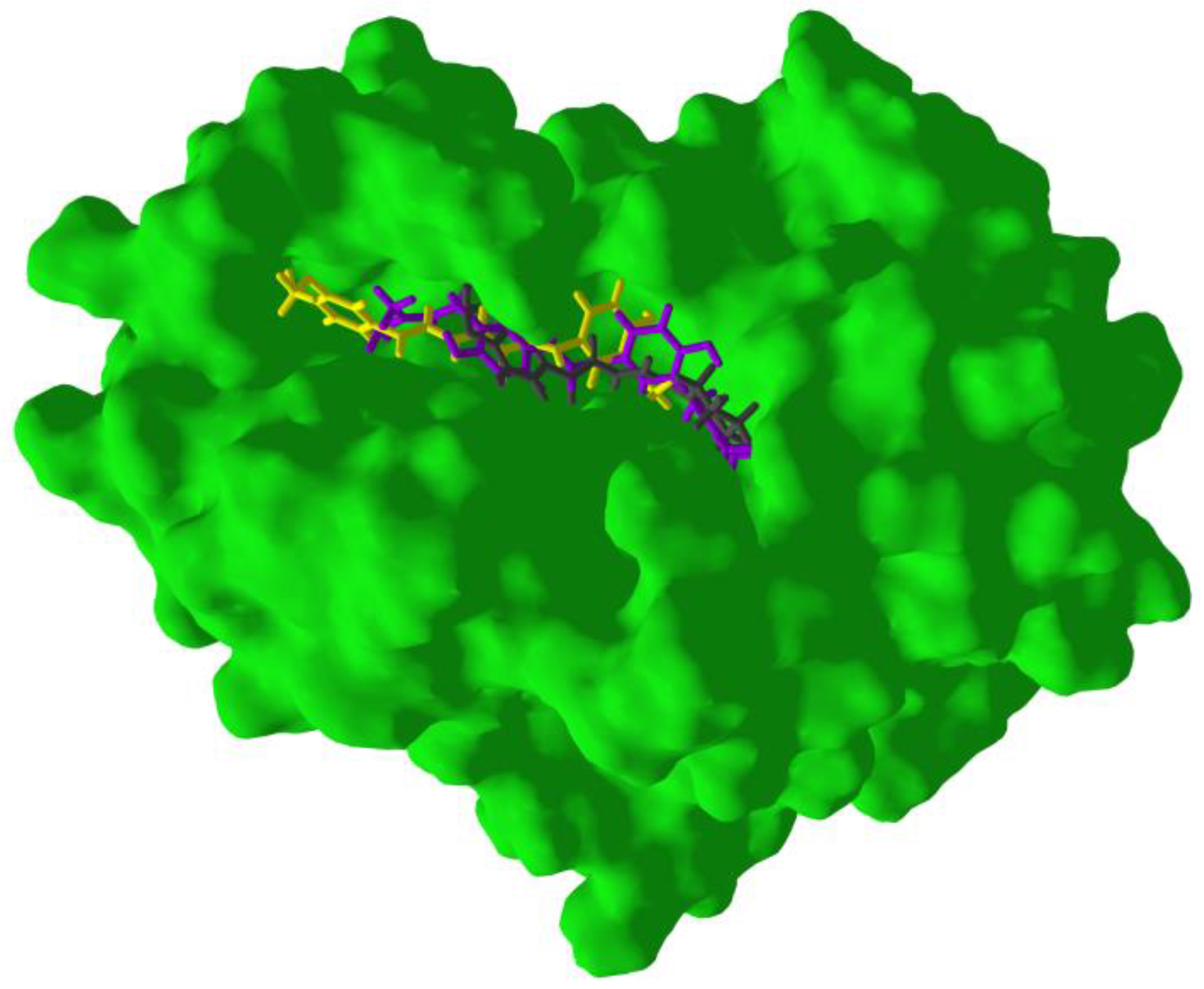
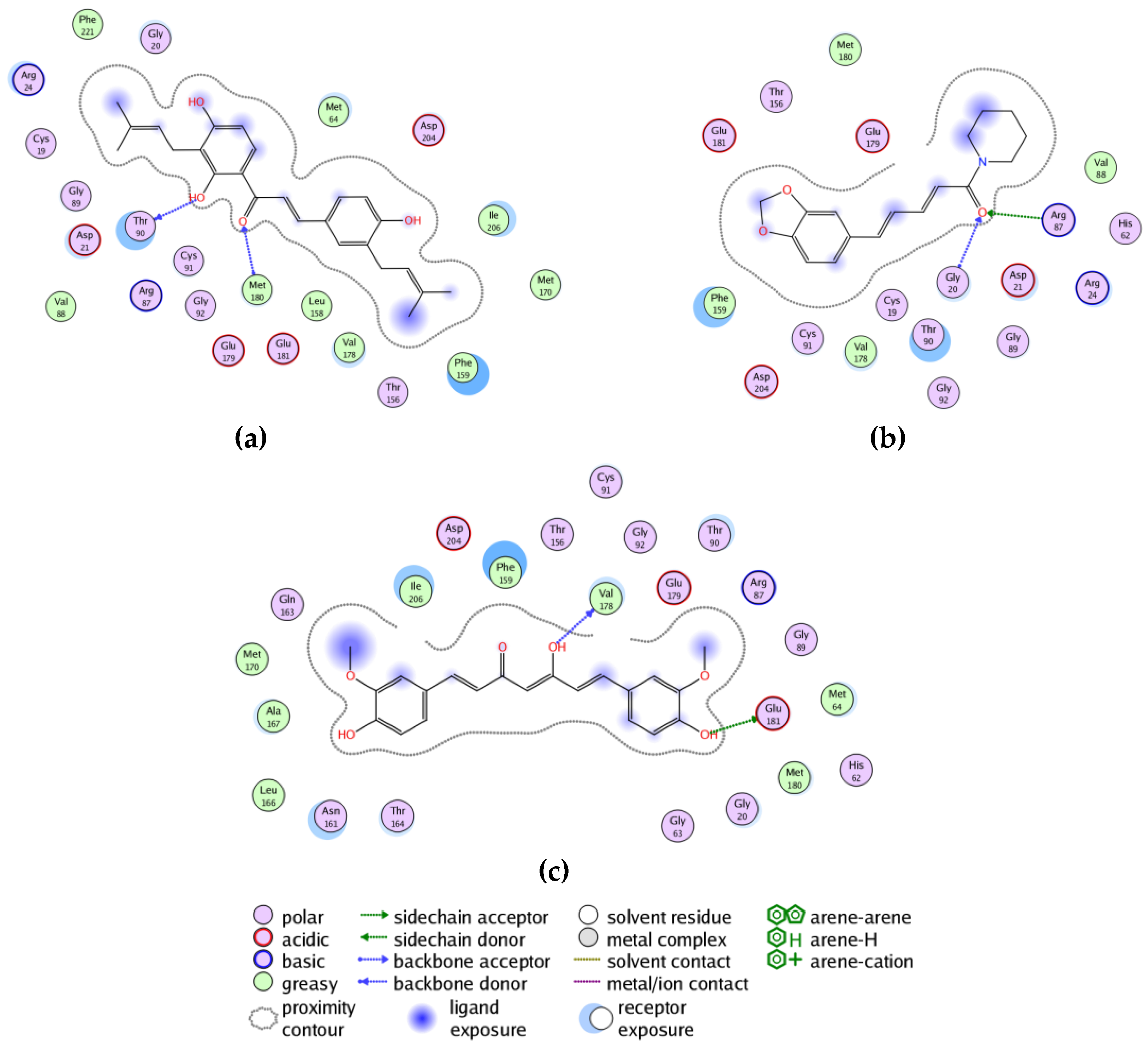
3. Results and Discussion
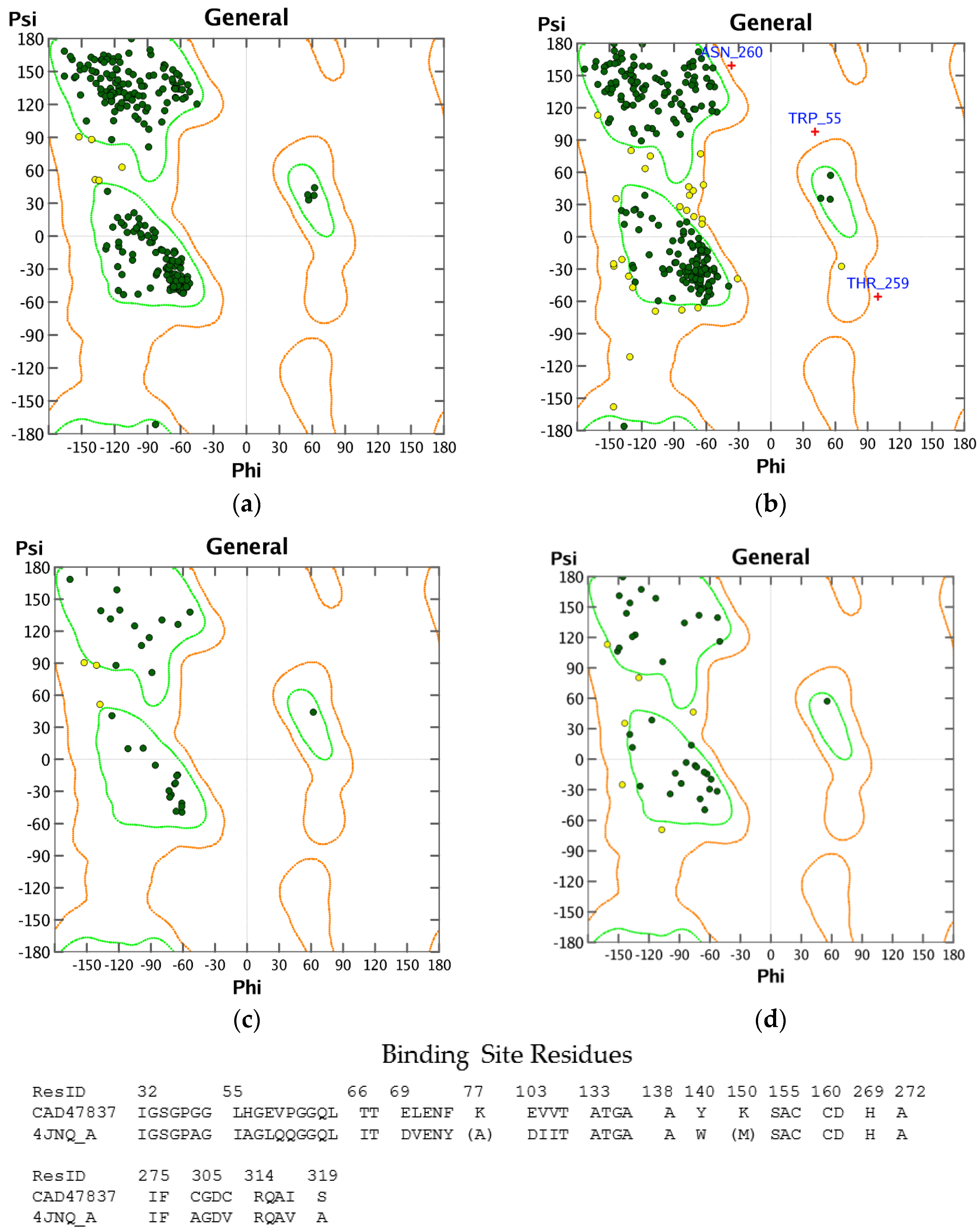
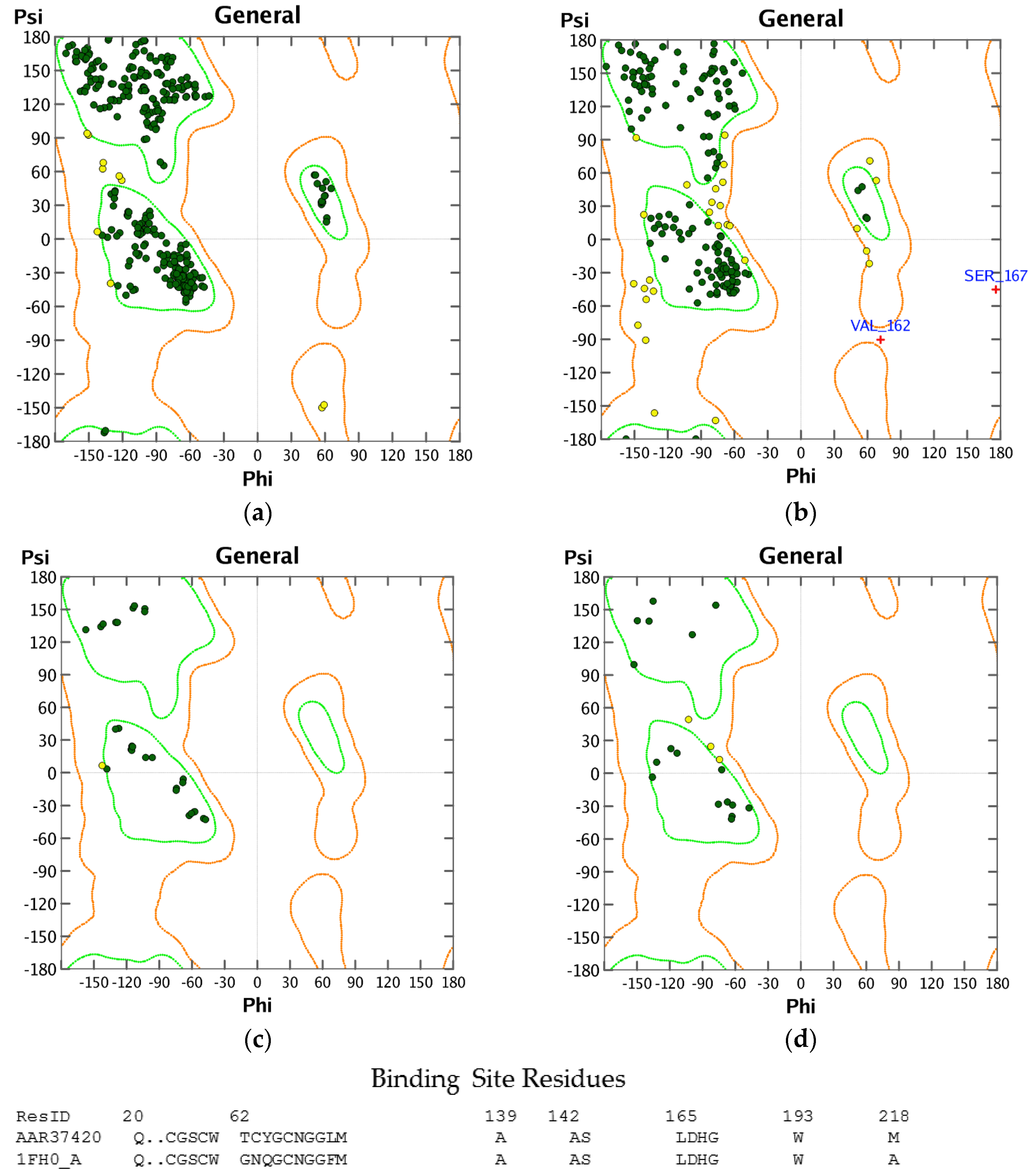
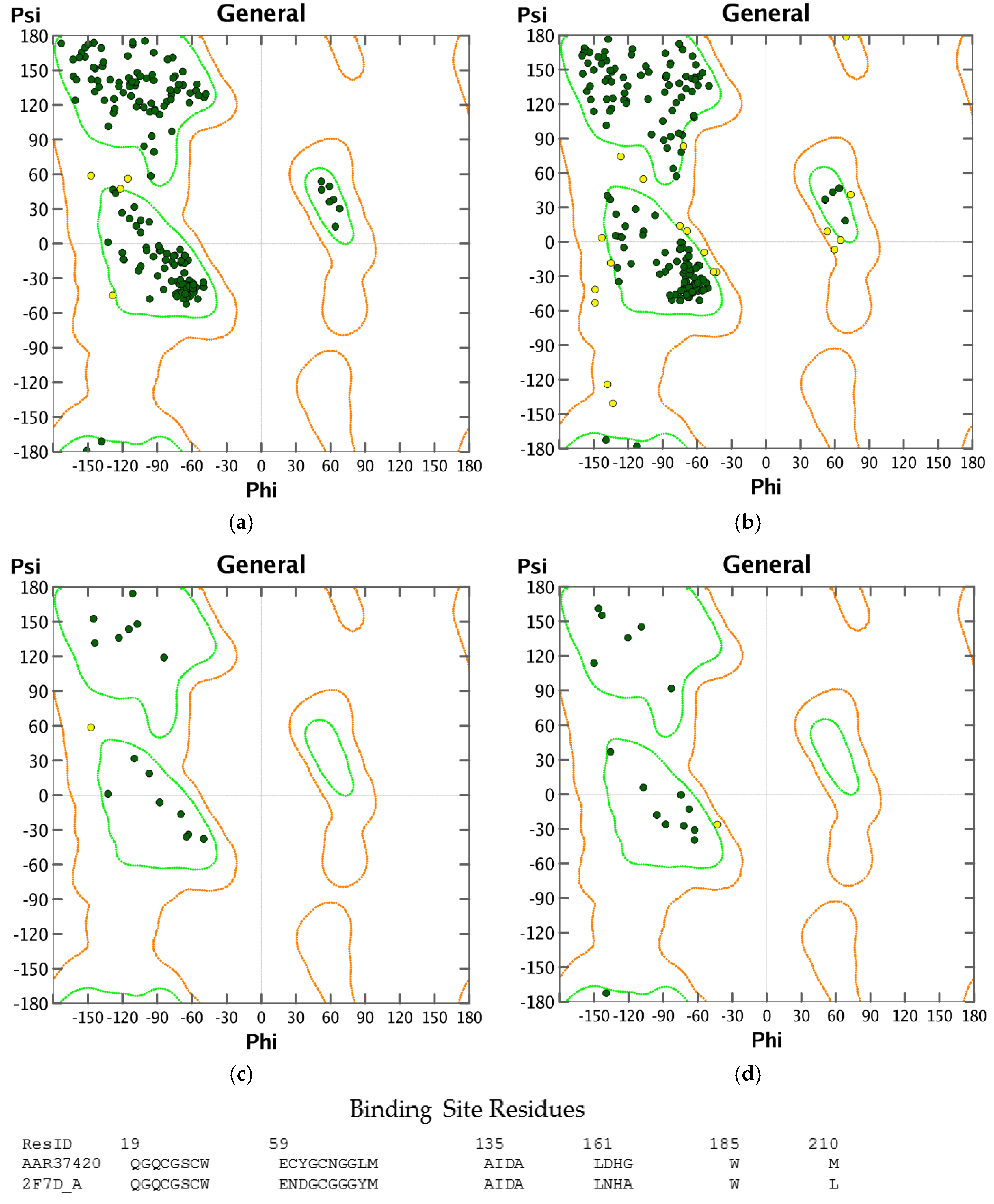
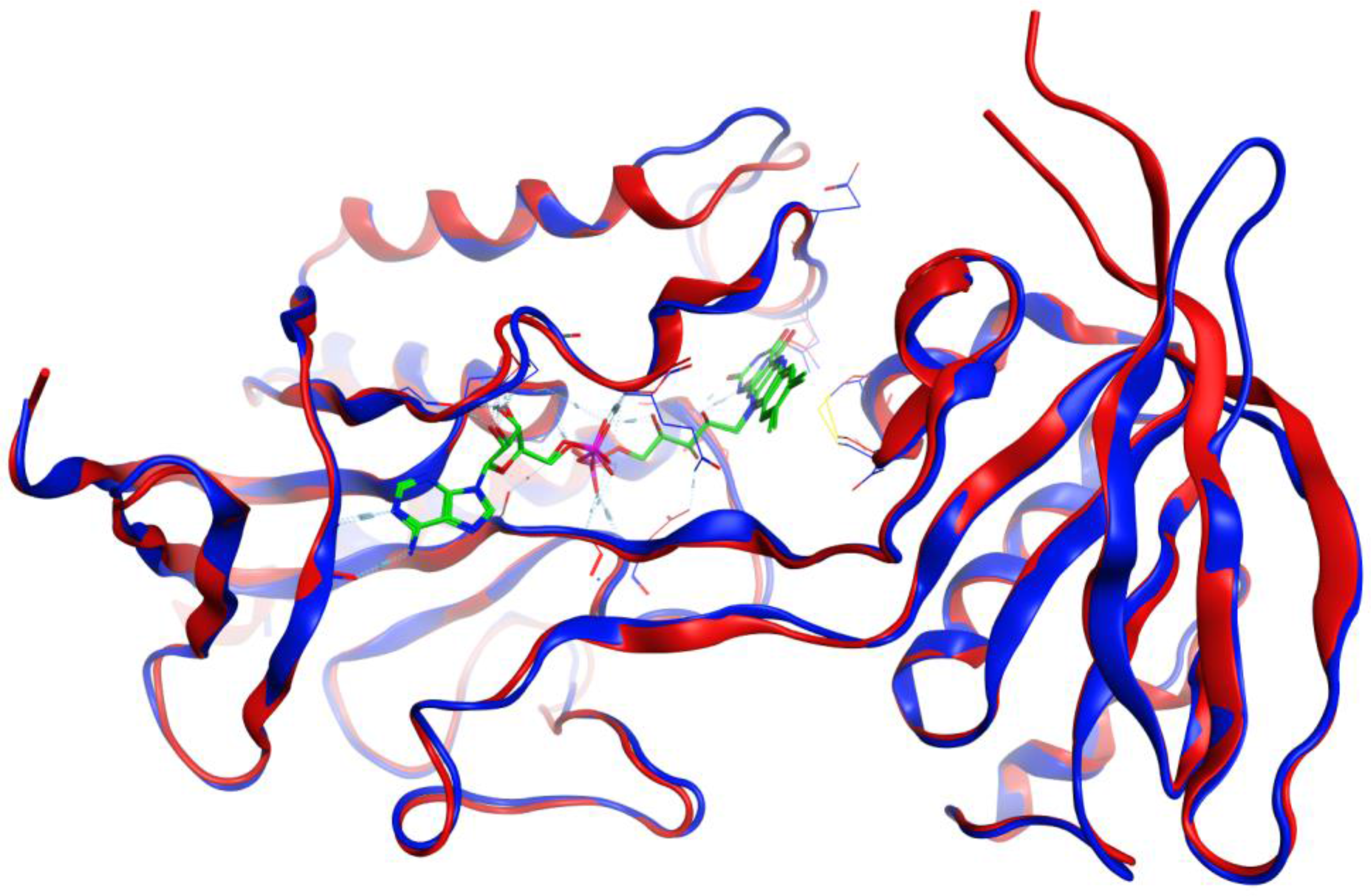
3.1. Homology Modeling
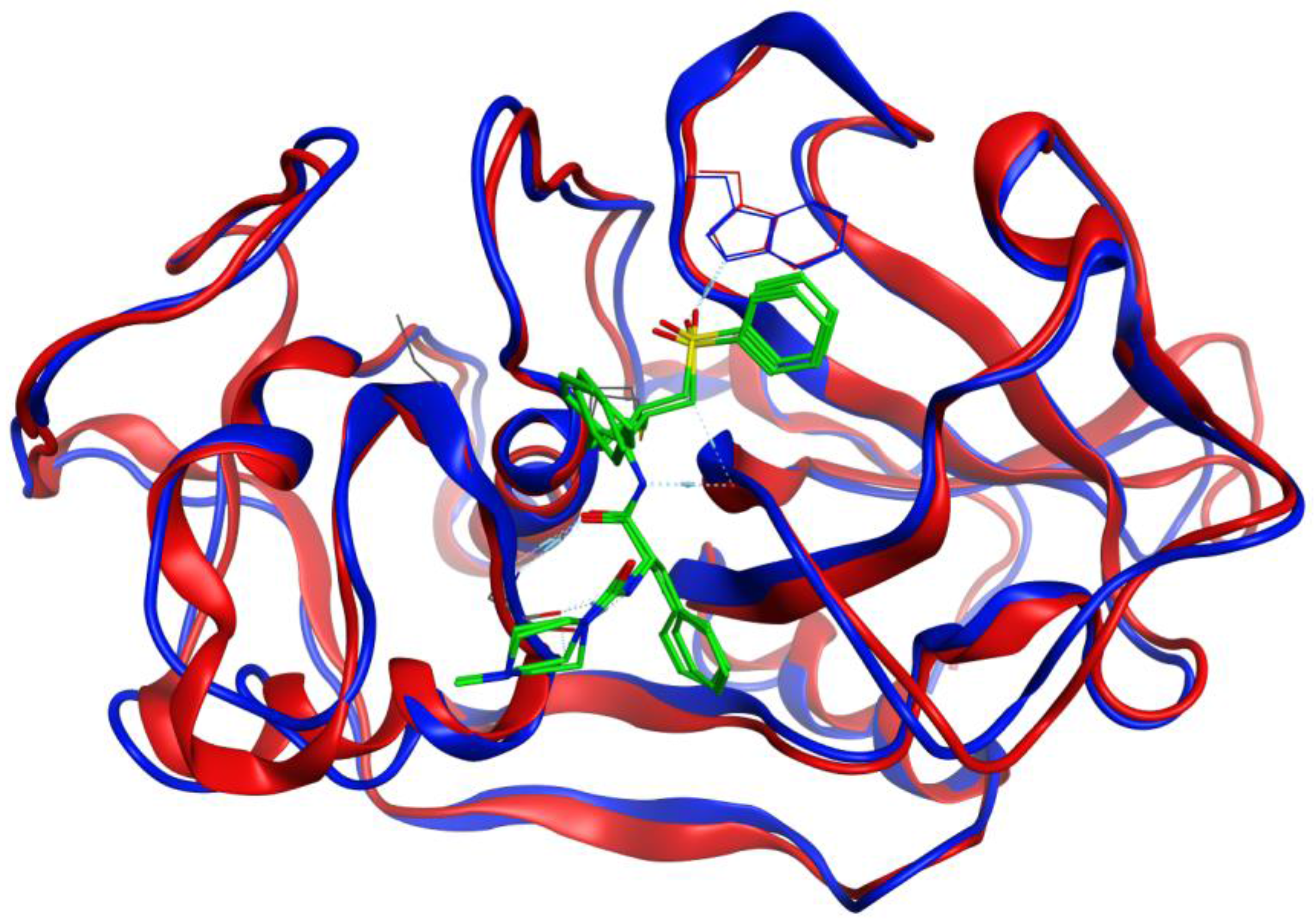
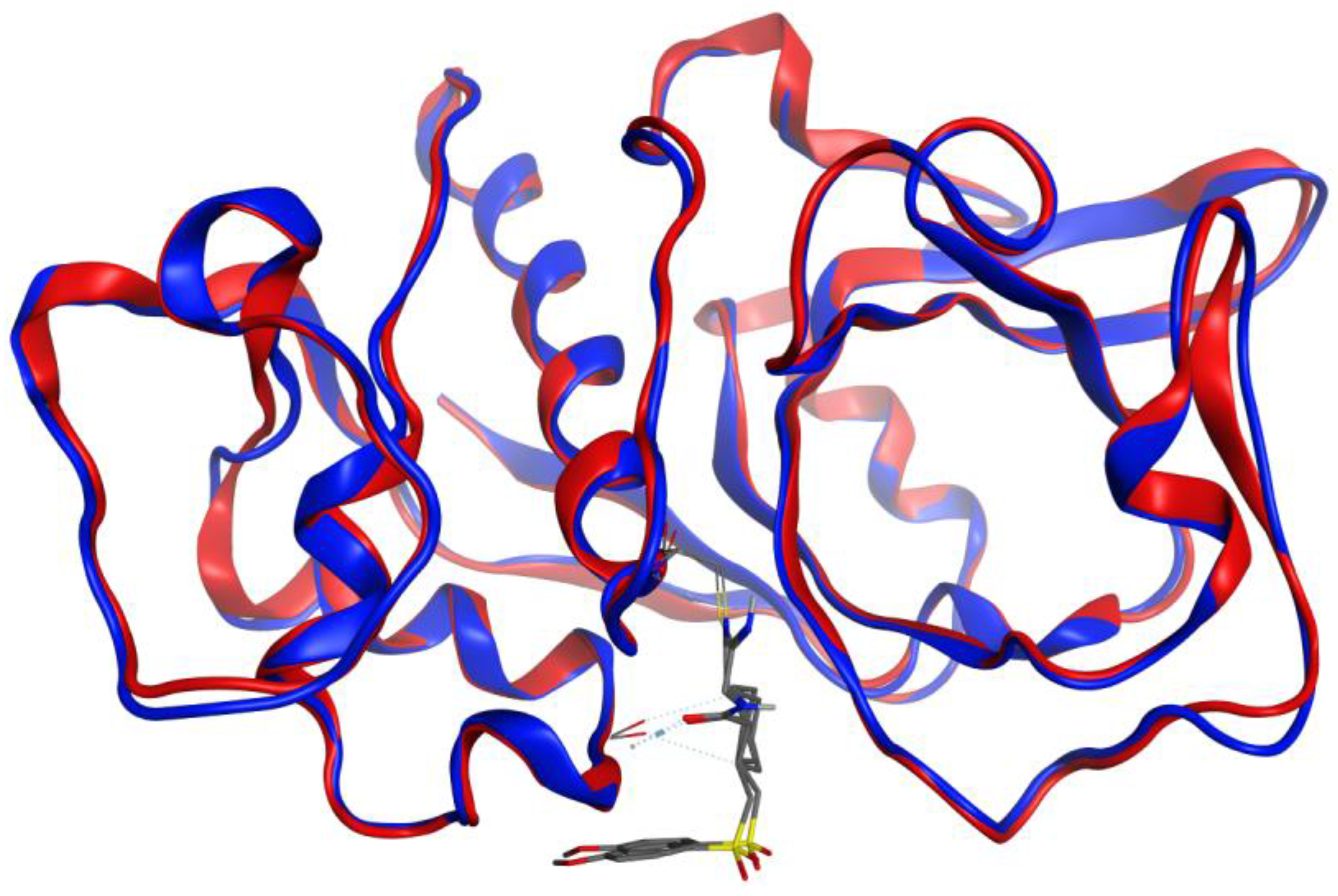
3.2. Molecular Docking Validation
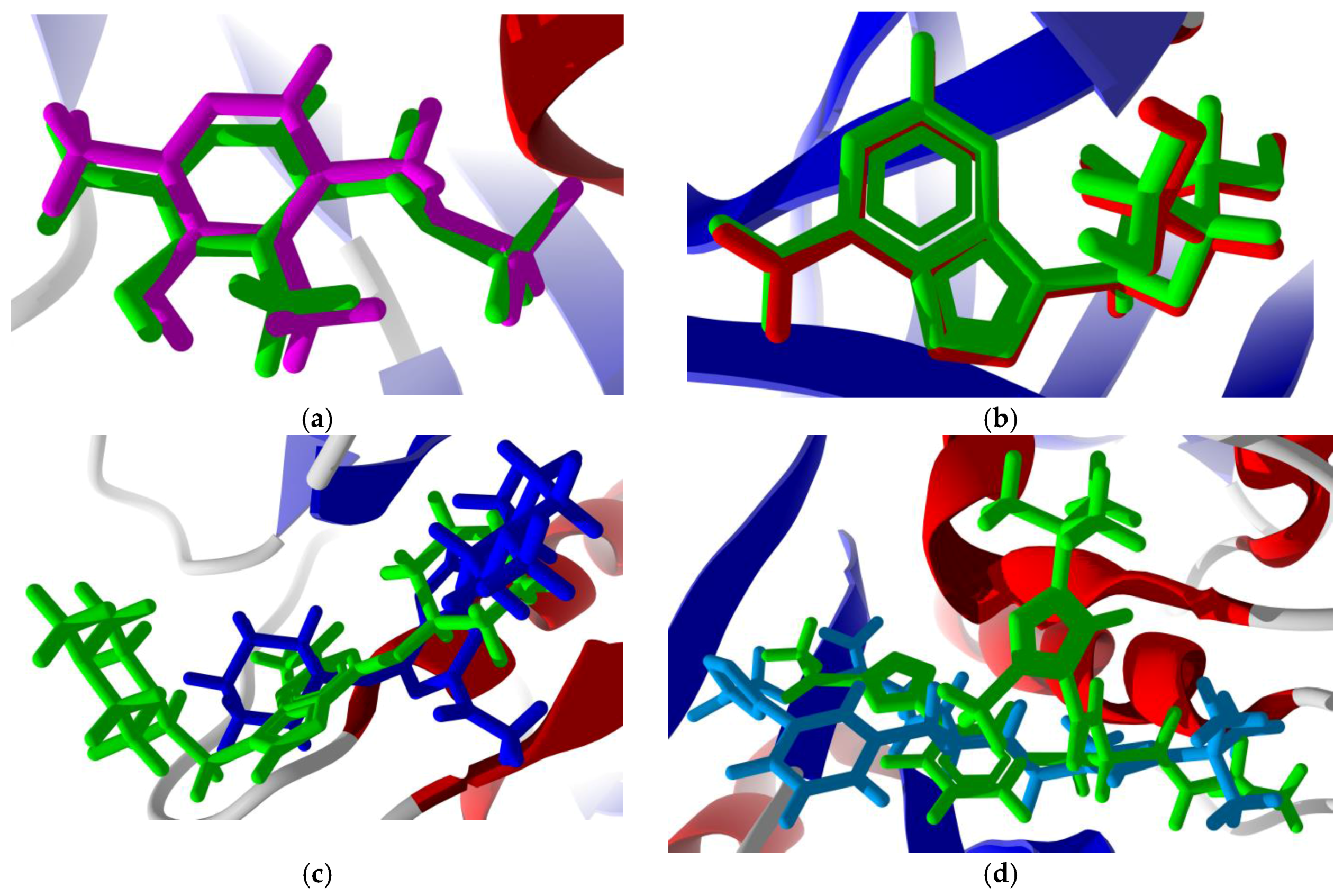
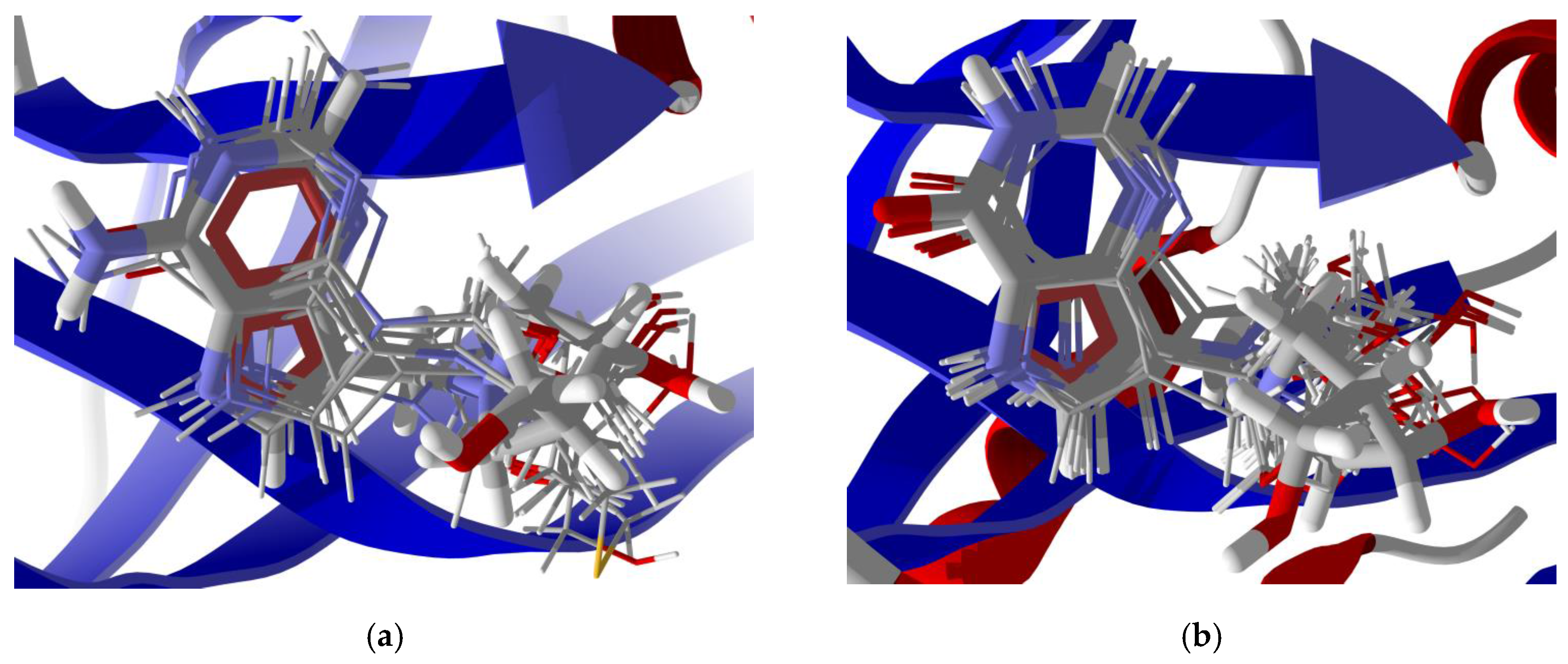
3.3. Molecular Docking of Phytochemicals
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Centers for Disease Control and Prevention. Trichomoniasis—CDC Fact Sheet. Available online: http://www.cdc.gov/std/trichomonas/STDFact-Trichomoniasis.htm (accessed on 22 October 2016).
- Dunne, R.L.; Dunn, L.A.; Upcroft, P.; O’Donoghue, P.J.; Upcroft, J.A. Drug resistance in the sexually transmitted protozoan Trichomonas vaginalis. Cell Res. 2003, 13, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Hirt, R.P.; de Miguel, N.; Nakjang, S.; Dessi, D.; Liu, Y.C.; Diaz, N.; Rappelli, P.; Acosta-Serrano, A.; Fiori, P.L.; Mottram, J.C. Trichomonas vaginalis pathobiology: New insights from the genome sequence. Adv. Parasitol. 2011, 77, 87–140. [Google Scholar] [PubMed]
- Carlton, J.M.; Hirt, R.P.; Silva, J.C.; Delcher, A.L.; Schatz, M.; Zhao, Q.; Wortman, J.R.; Bidwell, S.L.; Alsmark, U.C.; Besteiro, S.; et al. Draft genome sequence of the sexually transmitted pathogen Trichomonas vaginalis. Science 2007, 315, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Mande, S.C.; Mainfroid, V.; Kalk, K.H.; Goraj, K.; Martial, J.A.; Hol, W.G.J. Crystal structure of recombinant human triosephosphate isomerase at 2.8 Å resolution. Triosephosphate isomerase-related human genetic disorders and comparison with the trypanosomal enzyme. Protein Sci. 1994, 3, 810–821. [Google Scholar] [CrossRef] [PubMed]
- Ogungbe, I.V.; Setzer, W.N. The potential of secondary metabolites from plants as drugs or leads against protozoan neglected diseases—Part III: In-silico molecular docking investigations. Molecules 2016, 21, 1389. [Google Scholar] [CrossRef] [PubMed]
- Deck, L.M.; Royer, R.E.; Chamblee, B.B.; Hernandez, V.M.; Malone, R.R.; Torres, J.E.; Hunsaker, L.A.; Piper, R.C.; Makler, M.T.; Vander Jagt, D.L. Selective inhibitors of human lactate dehydrogenases and lactate dehydrogenase from the malarial parasite Plasmodium falciparum. J. Med. Chem. 1998, 41, 3879–3887. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Fixer, A.; ter Kuile, B.; Šali, A.; Müller, M. Convergent evolution of Trichomonas vaginalis lactate dehydrogenase from malate dehydrogenase. Proc. Natl. Acad. Sci. USA 1999, 96, 6285–6290. [Google Scholar] [CrossRef] [PubMed]
- Sato, D.; Nozaki, T. Methionone gamma-lyase: The unique reaction mechanism, physiological roles, and therapeutic applications against infectious diseases and cancers. IUBMB Life 2009, 61, 1019–1028. [Google Scholar] [CrossRef] [PubMed]
- Meyer, Y.; Buchanan, B.B.; Vignols, F.; Reichheld, J.P. Thioredoxins and glutaredoxins: Unifying elements in redox biology. Ann. Rev. Genet. 2009, 43, 335–367. [Google Scholar] [CrossRef] [PubMed]
- Hirt, R.P.; Müller, S.; Embley, T.M.; Coombs, G.H. The diversity and evolution of thioredoxin reductase: New perspectives. Trends Parasitol. 2002, 18, 302–308. [Google Scholar] [CrossRef]
- Huang, K.Y.; Chien, K.Y.; Lin, Y.C.; Hsu, W.M.; Fong, I.K.; Huang, P.J.; Yueh, Y.M.; Gan, R.R.C.; Tang, P. A proteome reference map of Trichomonas vaginalis. Parasitol. Res. 2009, 104, 927–933. [Google Scholar] [CrossRef] [PubMed]
- Leitsch, D.; Kolarich, D.; Binder, M.; Stadlmann, J.; Altmann, F.; Duchêne, M. Trichomonas vaginalis: Metronidazole and other nitroimidazole drugs are reduced by the flavin enzyme thioredoxin reductase and disrupt the cellular redox system. Implications for nitroimidazole toxicity and resistance. Mol. Microbiol. 2009, 72, 518–536. [Google Scholar] [CrossRef] [PubMed]
- Ealick, S.E.; Babu, Y.S.; Bugg, C.E.; Erion, M.D.; Guida, W.C.; Montgomery, J.A.; Secrist, J.A. Application of crystallographic and modeling methods in the design of purine nucleoside phosphorylase inhibitors. Proc. Natl. Acad. Sci. USA 1991, 88, 11540–11544. [Google Scholar] [CrossRef] [PubMed]
- Datta, A.K.; Datta, R.; Sen, B. Antiparasitic chemotherapy: Tinkering with the purine salvage pathway. Adv. Exp. Med. Biol. 2008, 625, 116–132. [Google Scholar] [PubMed]
- Saxena, A.; Sangwan, R.S.; Mishra, S. Fundamentals of homology modeling steps and comparison among important bioinformatics tools: An overview. Sci. Int. 2013, 1, 237–252. [Google Scholar]
- Cavasotto, C.N.; Phatak, S.S. Homology modeling in drug discovery: Current trends and applications. Drug Disc. Today 2009, 14, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Yuriev, E.; Agostino, M.; Ramsland, P.A. Challenges and advances in computational docking: 2009 in review. J. Mol. Recog. 2011, 24, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Yuriev, E.; Ramsland, P.A. Latest developments in molecular docking: 2010–2011 in review. J. Mol. Recog. 2013, 26, 215–239. [Google Scholar] [CrossRef] [PubMed]
- Yuriev, E.; Holien, J.; Ramsland, P.A. Improvements, trends, and new ideas in molecular docking: 2012–2013 in review. J. Mol. Recog. 2015, 28, 581–604. [Google Scholar] [CrossRef] [PubMed]
- Gerber, P.R.; Müller, K. MAB, a generally applicable molecular force field for structure modelling in medicinal chemistry. J. Comput. Aided Mol. Des. 1995, 9, 251–268. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cieplak, P.; Kollman, P.A. How well does a restrained electrostatic potential (RESP) model perform in calculating conformational energies of organic and biological molecules? J. Comp. Chem. 2000, 21, 1049–1074. [Google Scholar] [CrossRef]
- Case, D.A.; Darden, T.A.; Cheatham, T.E.; Simmerling, C.L.; Wang, J.; Duke, R.E.; Luo, R.; Crowley, M.; Walker, R.C.; Zhang, W.; et al. AMBER 10. University of California: San Francisco, CA, USA, 2008. [Google Scholar]
- Edwards, T.E.; Arakaki, T.L.; Seattle Structural Genomics Center for Infectious Disease. Crystal structure of a thioredoxin reductase from Brucella melitensis. To be published. [CrossRef]
- Crane, S.N.; Black, W.C.; Palmer, J.T.; Davis, D.E.; Setti, E.; Robichaud, J.; Paquet, J.; Oballa, R.M.; Bayly, C.I.; McKay, D.J.; et al. β-Substituted cyclohexanecarboxamide: A nonpeptidic framework for the design of potent inhibitors of cathepsin K. J. Med. Chem. 2006, 49, 1066–1079. [Google Scholar] [CrossRef] [PubMed]
- Somoza, J.R.; Zhan, H.; Bowman, K.K.; Yu, L.; Mortara, K.D.; Palmer, J.T.; Clark, J.M.; McGrath, M.E. Crystal structure of human cathepsin V. Biochemistry 2000, 39, 12543–12551. [Google Scholar] [CrossRef] [PubMed]
- Goodall, G.; Mottram, J.C.; Coombs, G.H.; Lapthorn, A.J. The structure and proposed catalytic mechanism of methionine gamma-lyase. To be published. [CrossRef]
- Zang, Y.; Wang, W.H.; Wu, S.W.; Ealick, S.E.; Wang, C.C. Identification of a subversive substrate of Trichomonas vaginalis purine nucleoside phosphorylase and the crystal structure of the enzyme-substrate complex. J. Biol. Chem. 2005, 280, 22318–22325. [Google Scholar] [CrossRef] [PubMed]
- Rinaldo-Matthis, A.; Wing, C.; Ghanem, M.; Deng, H.; Wu, P.; Gupta, A.; Tyler, P.C.; Evans, G.B.; Furneaux, R.H.; Almo, S.C.; et al. Inhibition and structure of Trichomonas vaginalis purine nucleoside phosphorylase with picomolar transition state analogues. Biochemistry 2007, 46, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Salgado-Lugo, H.; Lara-Gonzalez, S.; Brieba, L.G. Crystal structure of Trichomonas vaginalis triosephosphate isomerase TVAG_096350 gene. To be published. [CrossRef]
- Lara-Gonzalez, S.; Benitez-Cardoza, C.G.; Brieba, L.G. Engineering mutants with altered dimer-monomer equilibrium revale the existence of stable monomeric triosephosphate isomerases. To be published. [CrossRef]
- Steindel, P.A.; Chen, E.H.; Wirth, J.D.; Theobald, D.L. Gradual neofunctionalization in the convergent evolution of trichomonad lactate and malate dehydrogenases. Protein Sci. 2016, 25, 1319. [Google Scholar] [CrossRef] [PubMed]
- Rinaldo-Matthis, A.; Murkin, A.S.; Ramagopal, U.A.; Clinch, K.; Mee, S.P.; Evans, G.B.; Tyler, P.C.; Furneaux, R.H.; Almo, S.C.; Schramm, V.L. l-Enantiomers of transition state analogue inhibitors bound to human purine nucleoside phosphorylase. J. Am. Chem. Soc. 2008, 130, 842–844. [Google Scholar] [CrossRef] [PubMed]
- Caceres, R.A.; Timmers, L.F.; Pauli, I.; Gava, L.M.; Ducati, R.H.; Basso, L.A.; Santos, D.S.; de Azevedo, W.F. Crystal structure and molecular dynamics studies of human purine nucleoside phosphorylase complexed with 7-deazaguanine. J. Struct. Biol. 2010, 169, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Almazan, C.; Arreola, R.; Rodriguez-Larrea, D.; Aguirre-Lopez, B.; De Gomez-Puyou, M.T.; Perez-Montfort, R.; Costas, M.; Gomez-Puyou, A.; Lorres-Larios, A. Structural basis of human triosephosphate isomerase deficiency: Mutation E104D is related to alterations of a conserved water network at the dimer interface. J. Biol. Chem. 2008, 283, 23254–23263. [Google Scholar] [CrossRef] [PubMed]
- Roland, B.P.; Amrich, C.G.; Kammerer, C.J.; Stuchul, K.A.; Larsen, S.B.; Rode, S.; Aslam, A.A.; Heroux, A.; Wetzel, R.; VanDemark, A.P.; et al. Triosephosphate isomerase I170V alters catalytic site, enhances stability and induces pathology in a Drosophila model of TPI deficiency. Biochim. Biophys. Acta 2015, 1852, 61–69. [Google Scholar] [CrossRef] [PubMed]
- McGrath, M.E.; Klaus, J.L.; Barnes, M.G.; Bromme, D. Crystal structure of human cathepsin K complexed with a potent inhibitor. Nat. Struct. Biol. 1997, 4, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Altmann, E.; Cowan-Jacob, S.W.; Missbach, M. Novel purine nitrile derived inhibitors of the cysteine protease cathepsin K. J. Med. Chem. 2004, 47, 5833–5836. [Google Scholar] [CrossRef] [PubMed]
- Bethel, P.A.; Gerhardt, S.; Jones, E.V.; Kenny, P.W.; Karoutchi, G.I.; Morley, A.D.; Oldham, K.; Rankine, N.; Augustin, M.; Krapp, S.; et al. Design of selective cathepsin inhibitors. Bioorg. Med. Chem. Lett. 2009, 19, 4622–4625. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, R.T.; Sivaraman, J. Structural basis for reversible and irreversible inhibition of human cathepsin L by their respective dipeptidyl glyoxal and diazomethylketone inhibitors. J. Struct. Biol. 2011, 173, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, R.; Christensen, M.H. MolDock: A new technique for high-accuracy molecular docking. J. Med. Chem. 2006, 49, 3315–3321. [Google Scholar] [CrossRef] [PubMed]
- Dictionary of Natural Products on DVD, version 25:1; CRC Press: Boca Raton, FL, USA, 2016.
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 2012, 64, 4–17. [Google Scholar] [CrossRef]
- Halgren, T.A. Merck molecular force field. I. Basis, form, scope, parameterization, and performance of MMFF 94. J. Comput. Chem. 1996, 17, 490–519. [Google Scholar] [CrossRef]
- Pan, Y.; Huang, N.; Cho, S.; MacKerell, A.D. Consideration of molecular weight during compound selection in virtual target-based database screening. J. Chem. Inf. Comput. Sci. 2003, 43, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.M.; Shen, T.W. A pharmacophore-based evolutionary approach for screening selective estrogen receptor modulators. Proteins Struct. Funct. Bioinform. 2005, 59, 205–220. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.; Nagarsekar, A.; Xia, G.; Hayashi, J.; MacKerell, A.D. Identification of non-phosphate-containing small molecular weight inhibitors of the tyrosine kinase p56 Lck SH2 domain via in silico screening against the pY + 3 binding site. J. Med. Chem. 2004, 47, 3502–3511. [Google Scholar] [CrossRef] [PubMed]
- Hancock, C.N.; Macias, A.; Lee, E.K.; Yu, S.Y.; MacKerell, A.D.; Shapiro, P. Identification of novel extracellular signal-regulated kinase docking domain inhibitors. J. Med. Chem. 2005, 48, 4586–4595. [Google Scholar] [CrossRef] [PubMed]
- Abad-Zapatero, C.; Metz, J.T. Ligand efficiency indices as guideposts for drug discovery. Drug Discov. Today 2005, 10, 464–469. [Google Scholar] [CrossRef]
- Carta, G.; Knox, A.J.; Lloyd, D.G. Unbiasing scoring functions: A new normalization and rescoring strategy. J. Chem. Inf. Model. 2007, 47, 1564–1571. [Google Scholar] [CrossRef] [PubMed]
- Setzer, M.S.; Sharifi-Rad, J.; Setzer, W.N. The search for herbal antibiotics: An in-silico investigation of antibacterial phytochemicals. Antibiotics 2016, 5, 30. [Google Scholar] [CrossRef] [PubMed]
- Edwards, A.A.; Mason, J.M.; Clinch, K.; Tyler, P.C.; Evans, G.B.; Schramm, V.L. Altered enthalpy-entropy compensation in picomolar transition state analogues of human purine nucleoside phosphorylase. Biochemistry 2009, 48, 5226–5238. [Google Scholar] [CrossRef] [PubMed]
- Munagala, N.; Wang, C.C. The purine nucleoside phosphorylase from Trichomonas vaginalis is a homolog of the bacterial enzyme. Biochemistry 2002, 41, 10382–10389. [Google Scholar] [CrossRef] [PubMed]
- Leung-Toung, R.; Zhao, Y.; Li, W.; Tam, T.F.; Karimian, K.; Spino, M. Thiol proteases: Inhibitors and potential therapeutic targets. Curr. Med. Chem. 2006, 13, 547–581. [Google Scholar] [CrossRef] [PubMed]








 | ||
 | ||
 | ||
 HsCatK inhibitor 43 [54] |  HsCatK inhibitor 44 [54] |  HsCatK inhibitor 45 [54] |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | PDB code | Co-crystallized ligand | Edock (kJ/mol) | RMSD (Å) |
|---|---|---|---|---|
| TvMGL | 1E5E | N-(Hydroxy{3-hydroxy-2-methyl-5-[(phosphonooxy)methyl]pyridin-4-yl}methyl)norvaline | -127.6 | 1.13 |
| 1E5F | Pyridoxal-5′-phosphate | -69.4 | 0.57 | |
| TvCPCAC1 | [1FH0] a | N-α-[(4-Methylpiperazin-1-yl)carbonyl]-N-[(3S)-1-phenyl-5-(phenylsulfonyl)pentan-3-yl]-l-phenylalaninamide | -132.6 | 2.69 |
| TvCP2 | [2F7D] | (1R,2R)-N-(2-Aminoethyl)-2-{[(4-methoxyphenyl)sulfonyl]methyl}-cyclohexanecarboxamide | -97.7 | 1.22 |
| TvPNP | 1Z34 | 2-Fluoro-2′-deoxyadenosine | -97.1 | 0.91 |
| 1Z36 | (1S)-1-(7-Amino-1H-pyrazolo[4,3-d]pyrimidin-3-yl)-1,4-anhydro-d-ribitol | -97.7 | 0.21 | |
| 2ISC | (3R,4R)-1-[(4-Amino-5H-pyrrolo [3,2-d]pyrimidin-7-yl),ethyl]-4-(hydroxymethyl)pyrrolidin-3-ol | -104.0 | 0.57 | |
| HsCatK | 1MEM | N-{(1R)-3-Phenyl-1-[2-(phenylsulfonyl)ethyl]propyl}-N-2-(piperazin-1-ylcarbonyl)-l-leucinamide | -136.2 | 2.74 |
| 1U9V | 6-(Cyclohexylamino)-9-[2-(4-methylpiperazin-1-yl)-ethyl]-9H-purine-2-carbonitrile | -41.7 | 4.03 | |
| HsCatL | 3HWN | N-α-[(3-t-Butyl-1-methyl-1H-pyrazol-5-yl)carbonyl]-N-[(2E)-2-iminoethyl]-3-{5-[(Z)-iminomethyl]-1,3,4-oxadiazol-2-yl}-l-phenylalaninamide | -135.1 | 6.65 |
| 3OF8 | N-α-[(Benzyloxy)carbonyl]-N-[(2S)-1-(4-t-butoxyphenyl)-4-hydroxy-3-oxobutan-2-yl]-l-phenylalaninamide | -120.7 | 6.43 | |
| HsPNP | 3BGS | 3-Hydroxy-4-hydroxymethyl-1-(4-oxo-4,4a,5,7a-tetrahydro-3H-pyrrolo[3,2-d]pyrimidin-7-ylmethyl)-pyrrolidinium | -102.8 | 2.93 |
| 3INY | 7-Deazaguanine | -74.0 | 0.61 |
| Synthetic Inhibitor | TvPNP | HsPNP | ||
|---|---|---|---|---|
| Edock | DSnorm | Edock | DSnorm | |
| HsPNP inhibitor 1 [52] | −102.0 | −112.1 | −106.7 | −117.4 |
| HsPNP inhibitor 2 [52] | −103.9 | −116.5 | −108.4 | −121.6 |
| HsPNP inhibitor 3 [52] | −96.6 | −108.0 | −103.7 | −115.9 |
| HsPNP inhibitor 4 [52] | −96.0 | −111.7 | −98.8 | −114.9 |
| HsPNP inhibitor 5 [52] | −103.9 | −114.3 | −110.9 | −122.0 |
| HsPNP inhibitor 6 [52] | −103.0 | −117.8 | −105.6 | −120.8 |
| HsPNP inhibitor 7 [52] | −102.9 | −117.8 | −105.5 | −120.7 |
| HsPNP inhibitor 8 [52] | −94.6 | −107.7 | −102.9 | −117.1 |
| HsPNP inhibitor 9 [52] | −100.5 | −115.1 | −108.7 | −124.4 |
| HsPNP inhibitor 10 [52] | −101.9 | −114.0 | −108.5 | −121.4 |
| HsPNP inhibitor 11 [52] | −89.3 | −106.6 | −93.9 | −112.1 |
| HsPNP inhibitor 12 [52] | −85.8 | −105.1 | −95.7 | −117.3 |
| HsPNP inhibitor 13 [52] | −86.9 | −101.4 | −97.7 | −114.0 |
| HsPNP inhibitor 14 [52] | −86.4 | −100.8 | −100.8 | −117.7 |
| HsPNP inhibitor 15 [52] | −81.0 | −99.2 | −88.6 | −108.5 |
| HsPNP inhibitor 16 [52] | −93.6 | −107.1 | −104.2 | −119.3 |
| HsPNP inhibitor 17 [52] | −103.9 | −116.5 | −108.2 | −121.3 |
| TvPNP inhibitor 1 [29] | −101.6 | −116.4 | −98.9 | −113.3 |
| TvPNP inhibitor 2 [29] | −102.3 | −114.8 | −105.3 | −118.3 |
| TvPNP inhibitor 3 [29] | −102.9 | −117.8 | −105.5 | −120.8 |
| TvPNP inhibitor 4 [29] | −104.3 | −114.8 | −118.3 | −130.2 |
| TvPNP inhibitor 5 [29] | −103.7 | −116.3 | −108.4 | −121.6 |
| TvPNP inhibitor 6 [29] | −102.6 | −115.1 | −113.6 | −127.6 |
| TvPNP inhibitor 7 [29] | −104.9 | −115.7 | −107.0 | −118.0 |
| TvPNP inhibitor 8 [29] | −102.1 | −114.4 | −97.2 | −109.0 |
| TvPNP inhibitor 9 [29] | −89.7 | −98.9 | −99.0 | −109.2 |
| TvPNP inhibitor formycin A [53] | −101.1 | −113.2 | −102.2 | −114.4 |
| Compounds | TvCP2 | HsCatK | TvCPCAC1 | HsCatL | ||||
|---|---|---|---|---|---|---|---|---|
| Edock | DSnorm | Edock | DSnorm | Edock | DSnorm | Edock | DSnorm | |
| AURONES | ||||||||
| 4,6-Dibenzoyl-2-[phenylhydroxymethyl]-3(2H)-benzofuranone | −101.2 | −93.3 | −96.1 | −88.6 | −103.2 | −95.2 | −118.8 | −109.6 |
| 6-Benzoyl-2-[oxomethylpheny]-3-hydroxy-benzofurane | −87.6 | −90.3 | −92.3 | −95.2 | −101.9 | −105.1 | −114.4 | −118.0 |
| CHALCONES | ||||||||
| 1-[2,4-Dihydroxy-3-(3-methyl-2-butenyl)phenyl]-3-(8-hydroxy-2,2-dimethyl-2H-1-benzopyran-6-yl)-2-propen-1-one | −68.1 | −66.3 | −98.0 | −95.5 | −83.2 | −81.0 | −113.3 | −110.4 |
| 2',4,4',6'-Tetrahydroxy-3'-prenylchalcone | −91.8 | −94.8 | −94.3 | −97.5 | −102.0 | −105.4 | −95.8 | −99.0 |
| 2',4,4'-Trihydroxy-3',5'-diprenylchalcone | −82.5 | −81.3 | −103.0 | −101.5 | −96.0 | −94.6 | −112.9 | −111.2 |
| 2',4,4'-Trihydroxy-3'-prenylchalcone | −90.5 | −95.1 | −87.3 | −91.7 | −99.9 | −104.9 | −99.8 | −104.7 |
| 2',4,4'-Trihydroxy-3,3'-diprenylchalcone | −76.0 | −74.8 | −98.7 | −97.3 | −89.1 | −87.8 | −109.0 | −107.4 |
| 2',4,4'-Trihydroxy-6'-methoxy-3-prenylchalcone | −97.8 | −99.7 | −88.5 | −90.3 | −102.3 | −104.3 | −102.7 | −104.7 |
| 2'-Hydroxy-4,4'-dimethoxychalcone | −80.3 | −88.0 | −84.2 | −92.4 | −86.4 | −94.8 | −94.6 | −103.8 |
| 3-Methoxycitrunobin-4-methylether | −92.1 | −90.4 | −92.4 | −90.7 | −96.3 | −94.6 | −110.2 | −108.2 |
| Bipinnatone A | −98.8 | −91.1 | −105.6 | −97.4 | −103.4 | −95.4 | −119.9 | −110.6 |
| Bipinnatone B | −106.5 | −103.4 | −109.0 | −105.8 | −116.5 | −113.0 | −108.2 | −105.1 |
| Crotaorixin | −99.0 | −100.9 | −92.7 | −94.5 | −107.6 | −109.7 | −103.2 | −105.2 |
| Crotaramin | −83.0 | −85.9 | −86.6 | −89.7 | −91.2 | −94.4 | −90.5 | −93.7 |
| FLAVONOIDS | ||||||||
| 3'-O-Methyldiplacone | −97.2 | −92.3 | −104.3 | −99.0 | −116.0 | −110.2 | −105.2 | −99.9 |
| Cannflavin A | −100.3 | −95.4 | −100.3 | −95.4 | −112.8 | −107.3 | −113.5 | −107.9 |
| Diplacone | −102.7 | −98.6 | −91.7 | −88.0 | −117.9 | −113.2 | −110.3 | −105.9 |
| ISOFLAVONOIDS | ||||||||
| 2-Geranyl-3-hydroxy-8,9-methylenedioxypterocarpan | −89.1 | −85.8 | −97.3 | −93.7 | −97.6 | −94.0 | −113.6 | −109.4 |
| Andinermal A | −82.9 | −85.3 | −86.2 | −88.7 | −84.0 | −86.4 | −91.6 | −94.3 |
| Barbigerone | −75.1 | −73.9 | −80.2 | −78.9 | −86.0 | −84.6 | −74.8 | −73.6 |
| COUMARINS | ||||||||
| Auraptene | −83.6 | −90.3 | −93.1 | −100.5 | −100.4 | −108.4 | −103.1 | −111.3 |
| Umbelliprenin | −89.1 | −89.8 | −100.3 | −101.1 | −93.7 | −94.5 | −98.2 | −98.9 |
| LIGNANS | ||||||||
| (7R,8R,7'S,8'R)-3,3',4,5-Tetramethoxy-4',5'-methylenedioxy-7,7'-epoxylignan | −88.4 | −85.4 | −99.5 | −96.1 | −107.1 | −103.5 | −104.6 | −101.0 |
| (7R,8R,7'S,8'R)-3,3ʹ,5-Trimethoxy-4',5'-methylenedioxy-7,7'-epoxylignan | −91.4 | −89.3 | −83.3 | −81.4 | −94.4 | −92.3 | −79.9 | −78.0 |
| (7R,8S,7'S,8'S)-4,5,4',5'-Dimethylenedioxy-3,3'-dimethoxy-7,7'-epoxylignan | −95.7 | −93.7 | −80.8 | −79.1 | −98.6 | −96.5 | −105.0 | −102.7 |
| Austrobailignan 7 | −86.6 | −89.3 | −88.7 | −91.5 | −90.0 | −92.8 | −89.0 | −91.8 |
| Cubebin | −97.0 | −98.7 | −91.4 | −93.0 | −96.7 | −98.4 | −113.4 | −115.4 |
| Epieudesmin | −88.3 | −87.5 | −85.4 | −84.6 | −95.0 | −94.0 | −103.9 | −102.9 |
| Eupomatenoid 5 | −77.9 | −84.5 | −85.3 | −92.5 | −93.1 | −100.9 | −97.9 | −106.1 |
| Eupomatenoid 6 | −72.9 | −81.9 | −79.7 | −89.6 | −86.7 | −97.5 | −92.6 | −104.1 |
| Eupomatenoid 7 | −77.0 | −80.9 | −92.0 | −96.6 | −98.5 | −103.5 | −92.8 | −97.5 |
| Hinokinin | −99.9 | −101.8 | −87.6 | −89.4 | −95.7 | −97.6 | −109.8 | −111.9 |
| Sesamin | −94.9 | −96.7 | −94.5 | −96.3 | −99.6 | −101.5 | −98.1 | −100.0 |
| Yangambin | −91.4 | −86.3 | −75.2 | −71.0 | −85.6 | −80.8 | −89.0 | −84.0 |
| Lyonresinol a | −84.1 | −81.0 | −89.3 | −86.0 | −83.8 | −80.7 | −76.4 | −73.6 |
| MISCELLANEOUS POLYPHENOLICS | ||||||||
| 5-Acetyl-4-hydroxycannabigerol | −85.4 | −85.5 | −93.8 | −93.9 | −91.8 | −91.9 | −99.0 | −99.1 |
| Cannabigerolic acid | −101.1 | −102.5 | −103.8 | −105.2 | −109.9 | −111.4 | −107.6 | −109.0 |
| Curcumin | −87.3 | −87.8 | −99.4 | −100.1 | −105.3 | −106.0 | −112.2 | −112.9 |
| trans-4-(3-Methyl-E-but-1-enyl)-3,5,2',4'-tetrahydroxystilbene | −85.8 | −91.2 | −77.3 | −82.1 | −87.5 | −93.1 | −107.7 | −114.5 |
| trans-4-Isopentenyl-3,5,2',4'-tetrahydroxystilbene | −74.0 | −78.7 | −85.5 | −90.9 | −91.9 | −97.7 | −101.5 | −108.0 |
| Vanillic acid a | −52.5 | −68.6 | −60.8 | −79.4 | −57.6 | −75.3 | −58.0 | −75.8 |
| Veratric acid a | −54.1 | −68.8 | −64.8 | −82.4 | −59.2 | −75.3 | −63.2 | −80.4 |
| 1,3,6,8-Tetrahydroxy-2,5-dimethoxyxanthone a | −70.1 | −74.0 | −77.5 | −81.8 | −77.7 | −81.9 | −79.8 | −84.1 |
| 1,4-Dihydroxy-7-methoxyxanthone a | −64.4 | −72.9 | −70.0 | −79.2 | −74.3 | −84.1 | −75.7 | −85.7 |
| 1,6,8-Trihydroxy-2,3,4,7-tetramethoxyxanthone a | −72.6 | −73.4 | −74.0 | −74.8 | −71.7 | −72.4 | −78.5 | −79.3 |
| Securidacaxanthone A a | −63.8 | −63.6 | −72.5 | −72.3 | −85.3 | −85.0 | −76.8 | −76.6 |
| Securidacaxanthone B a | −63.3 | −63.1 | −75.7 | −75.5 | −76.3 | −76.1 | −72.6 | −72.4 |
| Securidacaxanthone C a | −69.3 | −65.4 | −79.3 | −74.9 | −48.4 | −45.7 | −68.5 | −64.7 |
| ALKALOIDS | ||||||||
| Δ1,6-Juliprosopine | −102.6 | −86.6 | −95.5 | −80.6 | −104.3 | −88.0 | −119.6 | −100.9 |
| Piperine | −78.2 | −85.7 | −80.7 | −88.4 | −90.8 | −99.5 | −97.9 | −107.3 |
| Tabernaemontanine a | −60.7 | −61.9 | −79.0 | −80.6 | −62.0 | −63.2 | −65.0 | −66.3 |
| Lycorine a | −71.8 | −78.5 | −83.9 | −91.7 | −81.9 | −89.6 | −75.8 | −82.9 |
| Candimine a | −85.4 | −87.8 | −87.2 | −89.7 | −77.7 | −79.9 | −79.3 | −81.6 |
| Securinine a | −59.6 | −71.5 | −59.4 | −71.3 | −62.4 | −74.9 | −60.6 | −72.7 |
| 5,6-Dihydroleptidine a | −45.0 | −43.5 | −70.7 | −68.3 | −47.8 | −46.2 | −91.6 | −88.5 |
| SESQUITERPENOIDS | ||||||||
| Lactucopicrin | −93.9 | −91.2 | −93.7 | −91.0 | −99.2 | −96.3 | −98.8 | −95.9 |
| Cnicin a | −87.2 | −87.0 | −105.9 | −105.6 | −96.3 | −96.1 | −86.9 | −86.7 |
| Coriolin a | −68.9 | −76.0 | −75.5 | −83.2 | −68.8 | −75.9 | −77.2 | −85.1 |
| TRITERPENOIDS | ||||||||
| 3β-Hydroxyurs-11-en-28,13β-lactone a | no dock | no dock | −44.1 | −41.4 | −64.0 | −60.1 | −56.9 | −53.4 |
| SYNTHETIC INHIBITORS | ||||||||
| Synthetic cysteine protease inhibitor 43 [54] | −95.2 | −94.2 | −106.2 | −105.0 | −69.8 | −69.0 | −99.9 | −98.8 |
| Synthetic cysteine protease inhibitor 44 [54] | −115.1 | −112.2 | −105.1 | −102.6 | −110.9 | −108.2 | −91.7 | −89.4 |
| Synthetic cysteine protease inhibitor 45 [54] | −69.8 | −79.8 | −74.5 | −85.1 | −73.2 | −83.6 | −80.4 | −91.9 |
| Compounds | TvTPI | HsTPI | TvLDH | TvTrxR | ||||
|---|---|---|---|---|---|---|---|---|
| Edock | DSnorm | Edock | DSnorm | Edock | DSnorm | Edock | DSnorm | |
| AURONES | ||||||||
| 4,6-Dibenzoyl-2-[phenylhydroxymethyl]-3(2H)-benzofuranone | −106.0 | −97.8 | −118.3 | −109.1 | −112.0 | −103.3 | −113.5 | −104.7 |
| 6-Benzoyl-2-[oxomethylpheny]-3-hydroxy-benzofurane | −99.4 | −102.5 | −97.9 | −101.0 | −103.9 | −107.1 | −105.1 | −108.4 |
| CHALCONES | ||||||||
| 1-[2,4-Dihydroxy-3-(3-methyl-2-butenyl)phenyl]-3-(8-hydroxy-2,2-dimethyl-2H-1-benzopyran-6-yl)-2-propen-1-one | −102.7 | −100.1 | −119.1 | −116.0 | −99.5 | −96.9 | −113.8 | −110.9 |
| 2′,4,4′,6′-Tetrahydroxy-3′-prenylchalcone | −100.7 | −104.1 | −105.3 | −108.8 | −96.7 | −99.9 | −75.1 | −77.6 |
| 2′,4,4′-Trihydroxy-3′,5′-diprenylchalcone | −112.9 | −111.3 | −130.3 | −128.4 | −102.9 | −101.4 | −93.3 | −91.9 |
| 2′,4,4′-Trihydroxy-3′-prenylchalcone | −99.9 | −104.9 | −105.4 | −110.6 | −95.1 | −99.9 | −101.7 | −106.8 |
| 2′,4,4′-Trihydroxy-3,3′-diprenylchalcone | −106.2 | −104.6 | −102.7 | −101.2 | −103.1 | −101.6 | −89.5 | −88.2 |
| 2′,4,4′-Trihydroxy-6′-methoxy-3-prenylchalcone | −102.9 | −104.9 | −113.3 | −115.5 | −95.6 | −97.5 | −95.8 | −97.6 |
| 2′-Hydroxy-4,4′-dimethoxychalcone | −86.9 | −95.4 | −91.1 | −100.0 | −87.7 | −96.2 | −85.8 | −94.1 |
| 3-Methoxycitrunobin-4-methylether | −101.6 | −99.8 | −97.5 | −95.8 | −96.7 | −95.0 | −89.2 | −87.6 |
| Bipinnatone A | −104.6 | −96.4 | −107.0 | −98.7 | −105.4 | −97.2 | −113.2 | −104.4 |
| Bipinnatone B | −108.9 | −105.7 | −124.2 | −120.6 | −113.9 | −110.6 | −123.5 | −119.8 |
| Crotaorixin | −96.9 | −98.7 | −106.9 | −109.0 | −100.0 | −101.9 | −105.7 | −107.8 |
| Crotaramin | −95.7 | −99.1 | −101.0 | −104.6 | −89.2 | −92.4 | −83.7 | −86.6 |
| FLAVONOIDS | ||||||||
| 3′-O-Methyldiplacone | −98.0 | −93.0 | −100.3 | −95.2 | −112.5 | −106.9 | −92.1 | −87.5 |
| Cannflavin A | −98.6 | −93.8 | −107.8 | −102.5 | −104.8 | −99.7 | −112.6 | −107.0 |
| Diplacone | −97.3 | −93.4 | −109.8 | −105.4 | −101.3 | −97.2 | −102.2 | −98.1 |
| ISOFLAVONOIDS | ||||||||
| 2-Geranyl-3-hydroxy-8,9-methylenedioxypterocarpan | −95.5 | −92.0 | −110.5 | −106.4 | −119.5 | −115.1 | −112.6 | −108.4 |
| Andinermal A | −92.4 | −95.1 | −88.8 | −91.4 | −86.4 | −89.0 | −102.2 | −105.2 |
| Barbigerone | −81.4 | −80.0 | −86.8 | −85.4 | −87.3 | −85.9 | −66.3 | −65.3 |
| COUMARINS | ||||||||
| Auraptene | −90.5 | −97.7 | −93.4 | −100.8 | −88.6 | −95.7 | −94.5 | −102.0 |
| Umbelliprenin | −93.0 | −93.7 | −113.0 | −113.9 | −95.8 | −96.6 | −90.4 | −91.1 |
| LIGNANS | ||||||||
| (7R,8R,7'S,8'R)-3,3',4,5-Tetramethoxy-4',5'-methylenedioxy-7,7'-epoxylignan | −91.7 | −88.6 | −97.5 | −94.2 | −105.2 | −101.6 | −85.5 | −82.6 |
| (7R,8R,7'S,8'R)-3,3′,5-Trimethoxy-4′,5′-methylenedioxy-7,7′-epoxylignan | −94.7 | −92.5 | −92.8 | −90.6 | −102.3 | −100.0 | −59.7 | −58.3 |
| (7R,8S,7′S,8′S)-4,5,4′,5′-Dimethylenedioxy-3,3′-dimethoxy-7,7′-epoxylignan | −100.2 | −98.0 | −107.7 | −105.4 | −102.6 | −100.4 | −91.1 | −89.2 |
| Austrobailignan 7 | −95.3 | −98.2 | −93.8 | −96.7 | −96.4 | −99.4 | −95.8 | −98.8 |
| Cubebin | −98.9 | −100.6 | −104.9 | −106.7 | −98.0 | −99.8 | −62.6 | −63.7 |
| Epieudesmin | −96.3 | −95.4 | −89.6 | −88.7 | −91.7 | −90.8 | −85.2 | −84.4 |
| Eupomatenoid 5 | −91.0 | −98.7 | −88.5 | −96.0 | −85.6 | −92.9 | −92.0 | −99.8 |
| Eupomatenoid 6 | −80.5 | −90.5 | −88.7 | −99.7 | −79.9 | −89.8 | −90.8 | −102.1 |
| Eupomatenoid 7 | −87.9 | −92.2 | −95.7 | −100.5 | −93.6 | −98.3 | −95.9 | −100.6 |
| Hinokinin | −103.0 | −105.1 | −96.3 | −98.2 | −98.5 | −100.5 | −78.5 | −80.0 |
| Sesamin | −93.8 | −95.6 | −101.8 | −103.7 | −93.6 | −95.5 | −90.1 | −91.8 |
| Yangambin | −100.6 | −95.0 | −95.5 | −90.2 | −93.3 | −88.1 | −64.7 | −61.1 |
| Lyonresinol a | −104.7 | −100.9 | −101.1 | −97.3 | −100.3 | −96.6 | −92.6 | −89.1 |
| MISCELLANEOUS POLYPHENOLICS | ||||||||
| 5-Acetyl-4-hydroxycannabigerol | −88.0 | −88.1 | −98.1 | −98.2 | −93.2 | −93.3 | −105.3 | −105.4 |
| Cannabigerolic acid | −92.3 | −93.6 | −99.2 | −100.6 | −93.8 | −95.1 | −113.0 | −114.6 |
| Curcumin | −101.4 | −102.0 | −108.7 | −109.4 | −101.5 | −102.2 | −115.6 | −116.3 |
| trans-4-(3-Methyl-E-but-1-enyl)-3,5,2',4'-tetrahydroxystilbene | −82.1 | −87.3 | −106.0 | −112.7 | −90.1 | −95.8 | −104.6 | −111.3 |
| trans-4-Isopentenyl-3,5,2',4'-tetrahydroxystilbene | −89.2 | −94.8 | −103.3 | −109.8 | −84.5 | −89.8 | −99.0 | −105.3 |
| Vanillic acid a | −63.1 | −82.5 | −61.7 | −80.6 | −61.0 | −79.8 | −64.3 | −84.1 |
| Veratric acid a | −66.4 | −84.5 | −66.6 | −84.7 | −64.5 | −82.1 | −68.4 | −87.1 |
| 1,3,6,8-Tetrahydroxy-2,5-dimethoxyxanthone a | −85.8 | −90.5 | −71.5 | −75.4 | −75.9 | −80.1 | −75.2 | −79.3 |
| 1,4-Dihydroxy-7-methoxyxanthone a | −76.0 | −86.1 | −66.6 | −75.5 | −72.8 | −82.5 | −64.8 | −73.4 |
| 1,6,8-Trihydroxy-2,3,4,7-tetramethoxyxanthone a | −85.2 | −86.1 | −65.4 | −66.0 | −78.5 | −79.3 | −73.8 | −74.5 |
| Securidacaxanthone A a | −88.3 | −88.0 | −76.7 | −76.5 | −76.3 | −76.1 | −71.8 | −71.6 |
| Securidacaxanthone B a | −85.0 | −84.8 | −74.4 | −74.2 | −74.9 | −74.7 | −70.3 | −70.2 |
| Securidacaxanthone C a | −78.9 | −74.5 | −81.3 | −76.8 | −82.3 | −77.7 | −78.5 | −74.1 |
| ALKALOIDS | ||||||||
| Δ1,6-Juliprosopine | −107.2 | −90.5 | −113.2 | −95.5 | −112.7 | −95.1 | −97.9 | −82.5 |
| Piperine | −82.3 | −90.1 | −87.1 | −95.4 | −83.1 | −91.1 | −77.3 | −84.7 |
| Tabernaemontanine a | −76.6 | −78.1 | −75.6 | −77.1 | −83.3 | −84.9 | −75.4 | −76.8 |
| Lycorine a | −71.8 | −78.4 | −81.0 | −88.5 | −77.9 | −85.1 | −74.5 | −81.5 |
| Candimine a | −86.0 | −88.4 | −77.6 | −79.7 | −86.6 | −89.1 | −72.6 | −74.7 |
| Securinine a | −65.1 | −78.1 | −60.5 | −72.5 | −70.7 | −84.8 | −54.0 | −64.7 |
| 5,6-Dihydroleptidine a | −54.2 | −52.3 | −77.7 | −75.1 | −91.4 | −88.4 | −56.6 | −54.7 |
| SESQUITERPENOIDS | ||||||||
| Lactucopicrin | −105.8 | −102.7 | −99.8 | −96.9 | −102.5 | −99.5 | −99.6 | −96.7 |
| Cnicin a | −99.6 | −99.4 | −102.7 | −102.4 | −98.0 | −97.7 | −92.5 | −92.3 |
| Coriolin a | −73.2 | −80.7 | −78.6 | −86.7 | −72.4 | −79.7 | −71.7 | −79.0 |
| TRITERPENOIDS | ||||||||
| 3β-Hydroxyurs-11-en-28,13β-lactone a | −47.7 | −44.8 | −63.7 | −59.7 | −70.0 | −65.7 | −50.7 | −47.6 |
| SYNTHETIC INHIBITORS | ||||||||
| Metronidazole | −64.4 | −83.7 | ||||||
| Tinidazole | −78.5 | −90.2 | ||||||
| Compounds | TvMGL | TvPNP | HsPNP | |||
|---|---|---|---|---|---|---|
| Edock | DSnorm | Edock | DSnorm | Edock | DSnorm | |
| AURONES | ||||||
| 4,6-Dibenzoyl-2-[phenylhydroxymethyl]-3(2H)-benzofuranone | −98.8 | −91.2 | −141.6 | −130.6 | −118.3 | −109.1 |
| 6-Benzoyl-2-[oxomethylpheny]-3-hydroxy-benzofurane | −121.7 | −125.5 | −130.8 | −134.9 | −120.2 | −124.0 |
| CHALCONES | ||||||
| 1-[2,4-Dihydroxy-3-(3-methyl-2-butenyl)phenyl]-3-(8-hydroxy-2,2-dimethyl-2H-1-benzopyran-6-yl)-2-propen-1-one | −118.5 | −115.4 | −137.2 | −133.6 | −92.1 | −89.7 |
| 2',4,4',6'-Tetrahydroxy-3'-prenylchalcone | −115.7 | −119.6 | −122.2 | −126.3 | −112.3 | −116.1 |
| 2',4,4'-Trihydroxy-3',5'-diprenylchalcone | −97.4 | −96.0 | −134.9 | −132.9 | −119.1 | −117.3 |
| 2',4,4'-Trihydroxy-3'-prenylchalcone | −116.0 | −121.8 | −122.9 | −129.0 | −111.6 | −117.2 |
| 2',4,4'-Trihydroxy-3,3'-diprenylchalcone | −126.5 | −124.7 | −143.4 | −141.3 | −107.5 | −105.9 |
| 2',4,4'-Trihydroxy-6'-methoxy-3-prenylchalcone | −107.1 | −109.2 | −130.6 | −133.2 | −124.2 | −126.6 |
| 2'-Hydroxy-4,4'-dimethoxychalcone | −103.5 | −113.6 | −114.9 | −126.0 | −100.7 | −110.5 |
| 3-Methoxycitrunobin-4-methylether | −102.5 | −100.6 | −129.5 | −127.1 | −108.7 | −106.8 |
| Bipinnatone A | −122.7 | −113.1 | −146.4 | −135.1 | −142.5 | −131.4 |
| Bipinnatone B | −120.3 | −116.7 | −139.3 | −135.2 | −133.4 | −129.5 |
| Crotaorixin | −120.2 | −122.5 | −127.3 | −129.7 | −117.0 | −119.3 |
| Crotaramin | −114.8 | −118.9 | −121.1 | −125.3 | −107.1 | −110.9 |
| FLAVONOIDS | ||||||
| 3'-O-Methyldiplacone | −106.2 | −100.8 | −135.6 | −128.8 | −117.3 | −111.4 |
| Cannflavin A | −118.0 | −112.2 | −138.5 | −131.7 | −125.4 | −119.3 |
| Diplacone | −109.2 | −104.8 | −137.5 | −132.0 | −133.7 | −128.4 |
| ISOFLAVONOIDS | ||||||
| 2-Geranyl-3-hydroxy-8,9-methylenedioxypterocarpan | −75.5 | −72.7 | −139.1 | −134.0 | −125.9 | −121.3 |
| Andinermal A | −104.6 | −107.7 | −121.4 | −125.0 | −104.0 | −107.0 |
| Barbigerone | −94.9 | −93.4 | −132.3 | −130.1 | −100.2 | −98.6 |
| COUMARINS | ||||||
| Auraptene | −104.5 | −112.9 | −117.0 | −126.3 | −107.3 | −115.8 |
| Umbelliprenin | −119.3 | −120.3 | −129.9 | −130.9 | −110.3 | −111.2 |
| LIGNANS | ||||||
| (7R,8R,7'S,8'R)-3,3',4,5-Tetramethoxy-4',5'-methylenedioxy-7,7'-epoxylignan | −76.2 | −73.6 | −131.8 | −127.4 | −99.0 | −95.6 |
| (7R,8R,7'S,8'R)-3,3ʹ,5-Trimethoxy-4',5'-methylenedioxy-7,7'-epoxylignan | −89.7 | −87.6 | −129.7 | −126.7 | −107.6 | −105.1 |
| (7R,8S,7'S,8'S)-4,5,4',5'-Dimethylenedioxy-3,3'-dimethoxy-7,7'-epoxylignan | −95.7 | −93.7 | −134.2 | −131.3 | −101.2 | −99.1 |
| Austrobailignan 7 | −85.0 | −87.6 | −122.9 | −126.8 | −106.1 | −109.4 |
| Cubebin | −110.0 | −112.0 | −125.9 | −128.1 | −110.2 | −112.1 |
| Epieudesmin | −94.2 | −93.3 | −129.9 | −128.7 | −100.8 | −99.9 |
| Eupomatenoid 5 | −120.1 | −130.3 | −110.7 | −120.1 | −121.1 | −131.3 |
| Eupomatenoid 6 | −116.9 | −131.4 | −105.7 | −118.8 | −113.1 | −127.1 |
| Eupomatenoid 7 | −128.3 | −134.7 | −112.0 | −117.6 | −111.7 | −117.3 |
| Hinokinin | −113.2 | −115.4 | −124.5 | −126.9 | −105.6 | −107.6 |
| Sesamin | −104.4 | −106.4 | −127.3 | −129.8 | −106.3 | −108.4 |
| Yangambin | −97.0 | −91.6 | −134.1 | −126.6 | −97.8 | −92.3 |
| Lyonresinol a | −50.0 | −48.1 | −99.7 | −96.0 | −91.0 | −87.6 |
| MISCELLANEOUS POLYPHENOLICS | ||||||
| 5-Acetyl-4-hydroxycannabigerol | −127.4 | −127.6 | −115.6 | −115.7 | −106.0 | −106.1 |
| Cannabigerolic acid | −134.3 | −136.2 | −122.5 | −124.1 | −112.4 | −113.9 |
| Curcumin | −113.9 | −114.6 | −135.2 | −136.1 | −134.5 | −135.3 |
| trans-4-(3-Methyl-E-but-1-enyl)-3,5,2',4'-tetrahydroxystilbene | −120.2 | −127.8 | −117.8 | −125.3 | −117.5 | −124.9 |
| trans-4-Isopentenyl-3,5,2',4'-tetrahydroxystilbene | −119.2 | −126.8 | −115.3 | −122.6 | −111.3 | −118.3 |
| Vanillic acid a | −72.5 | −94.7 | −70.9 | −92.6 | −70.1 | −91.7 |
| Veratric acid a | −75.7 | −96.3 | −70.9 | −90.2 | −72.0 | −91.6 |
| 1,3,6,8-Tetrahydroxy-2,5-dimethoxyxanthone a | −93.2 | −98.2 | −91.3 | −96.3 | −91.2 | −96.2 |
| 1,4-Dihydroxy-7-methoxyxanthone a | −85.2 | −96.5 | −86.8 | −98.3 | −99.3 | −112.5 |
| 1,6,8-Trihydroxy-2,3,4,7-tetramethoxyxanthone a | −95.8 | −96.8 | −92.9 | −93.9 | −85.3 | −86.2 |
| Securidacaxanthone A a | −78.1 | −77.9 | −92.2 | −92.0 | −60.1 | −60.0 |
| Securidacaxanthone B a | −91.8 | −91.5 | −93.9 | −93.7 | −69.1 | −68.9 |
| Securidacaxanthone C a | −104.5 | −98.6 | −108.6 | −102.5 | −82.6 | −78.0 |
| ALKALOIDS | ||||||
| Δ1,6-Juliprosopine | −113.8 | −96.0 | −159.7 | −134.7 | −117.8 | −99.3 |
| Piperine | −79.8 | −87.5 | −123.3 | −135.1 | −101.1 | −110.8 |
| Tabernaemontanine a | −55.0 | −56.0 | −91.3 | −93.1 | −88.0 | −89.7 |
| Lycorine a | −85.3 | −93.3 | −88.1 | −96.3 | −88.8 | −97.1 |
| Candimine a | −72.7 | −74.8 | −89.5 | −92.0 | −89.4 | −91.9 |
| Securinine a | −66.7 | −80.0 | −75.1 | −90.1 | −80.0 | −96.0 |
| 5,6-Dihydroleptidine a | −27.4 | −26.4 | −86.0 | −83.2 | −90.2 | −87.2 |
| SESQUITERPENOIDS | ||||||
| Lactucopicrin | −115.0 | −111.6 | −133.1 | −129.2 | −125.4 | −121.8 |
| Cnicin a | −69.8 | −69.6 | −123.9 | −123.6 | −97.0 | −96.7 |
| Coriolin a | no dock | no dock | −87.9 | −96.9 | −83.0 | −91.5 |
| TRITERPENOIDS | ||||||
| 3β-Hydroxyurs-11-en-28,13β-lactone a | −15.2 | −14.3 | −69.9 | −65.6 | −73.2 | −68.7 |
| SYNTHETIC INHIBITORS | ||||||
| Purine nucleoside phosphorylase inhibitor 4 [52] | −101.4 | −118.0 | −96.9 | −112.7 | −102.6 | −119.3 |
| Purine nucleoside phosphorylase inhibitor 5 [52] | −94.3 | −103.7 | −103.9 | −114.3 | −111.5 | −122.6 |
| Purine nucleoside phosphorylase inhibitor 12 [52] | −84.4 | −103.4 | −85.8 | −105.1 | −95.7 | −117.2 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Setzer, M.S.; Byler, K.G.; Ogungbe, I.V.; Setzer, W.N. Natural Products as New Treatment Options for Trichomoniasis: A Molecular Docking Investigation. Sci. Pharm. 2017, 85, 5. https://doi.org/10.3390/scipharm85010005
Setzer MS, Byler KG, Ogungbe IV, Setzer WN. Natural Products as New Treatment Options for Trichomoniasis: A Molecular Docking Investigation. Scientia Pharmaceutica. 2017; 85(1):5. https://doi.org/10.3390/scipharm85010005
Chicago/Turabian StyleSetzer, Mary Snow, Kendall G. Byler, Ifedayo Victor Ogungbe, and William N. Setzer. 2017. "Natural Products as New Treatment Options for Trichomoniasis: A Molecular Docking Investigation" Scientia Pharmaceutica 85, no. 1: 5. https://doi.org/10.3390/scipharm85010005
APA StyleSetzer, M. S., Byler, K. G., Ogungbe, I. V., & Setzer, W. N. (2017). Natural Products as New Treatment Options for Trichomoniasis: A Molecular Docking Investigation. Scientia Pharmaceutica, 85(1), 5. https://doi.org/10.3390/scipharm85010005