
A Spatial Approach for Modeling Amphibian Road-Kills: Comparison of Regression Techniques



Abstract
1. Introduction
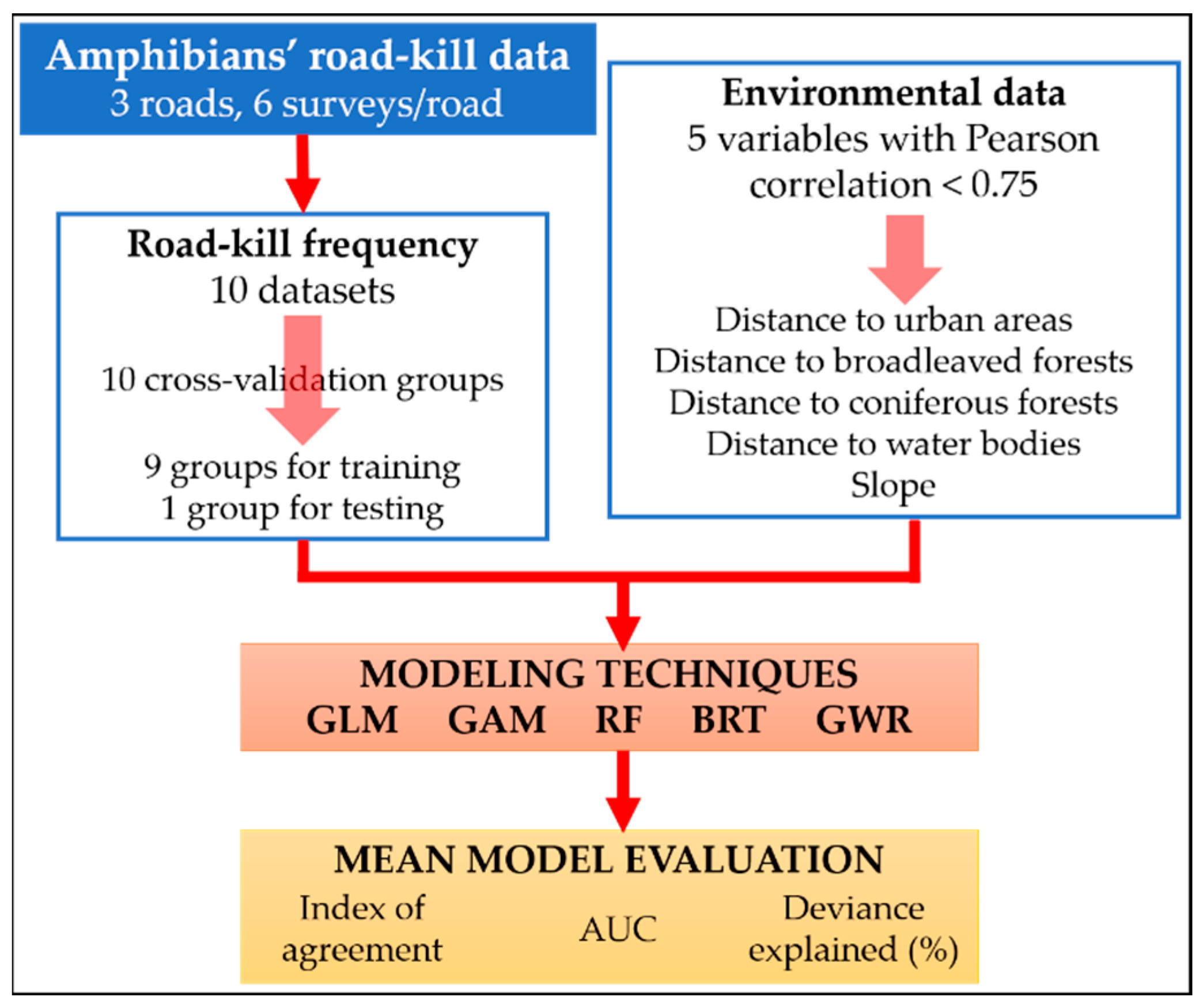
2. Materials and Methods
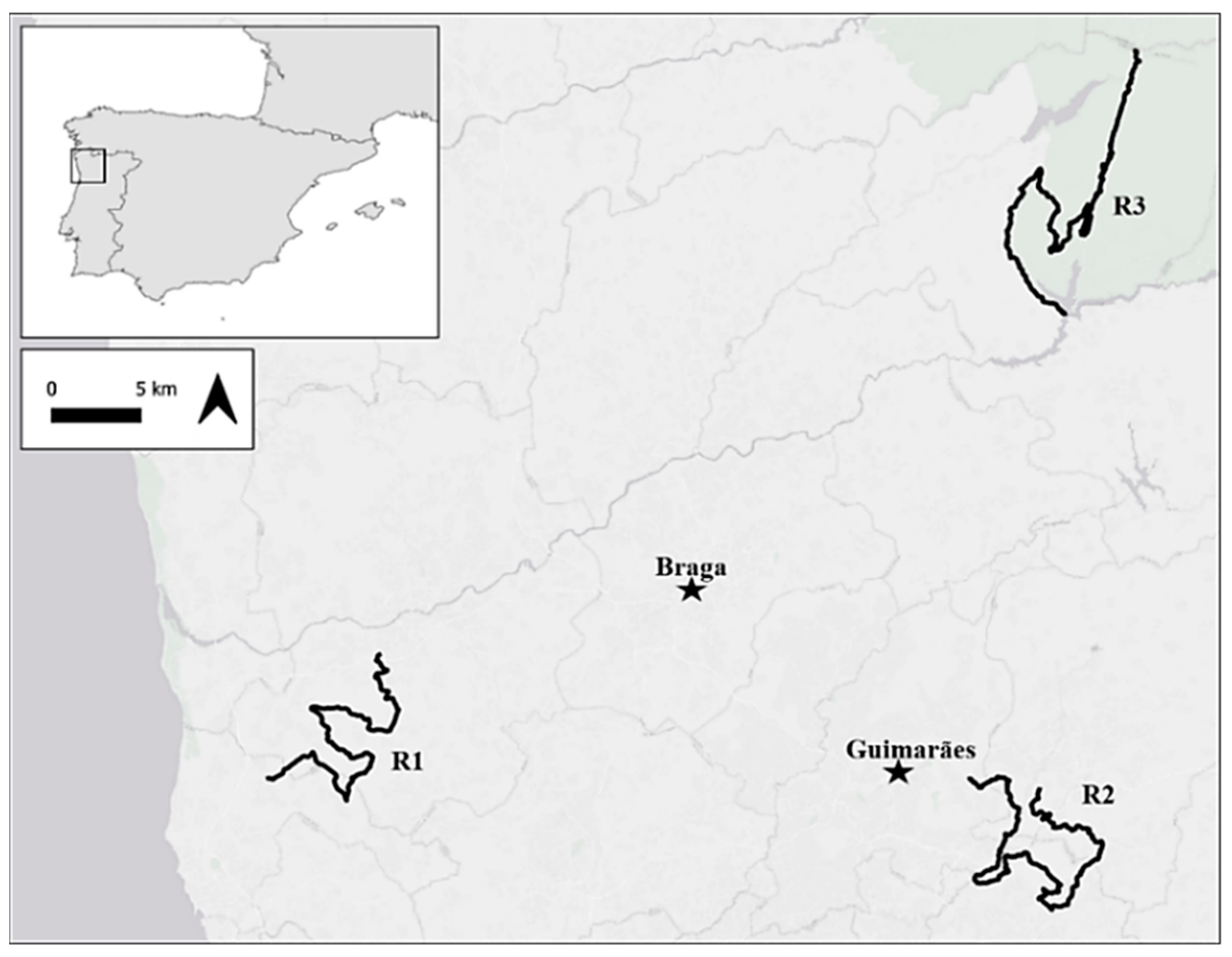
2.1. Road-Kills’ Data
2.2. Environmental Data
2.3. Modeling Techniques
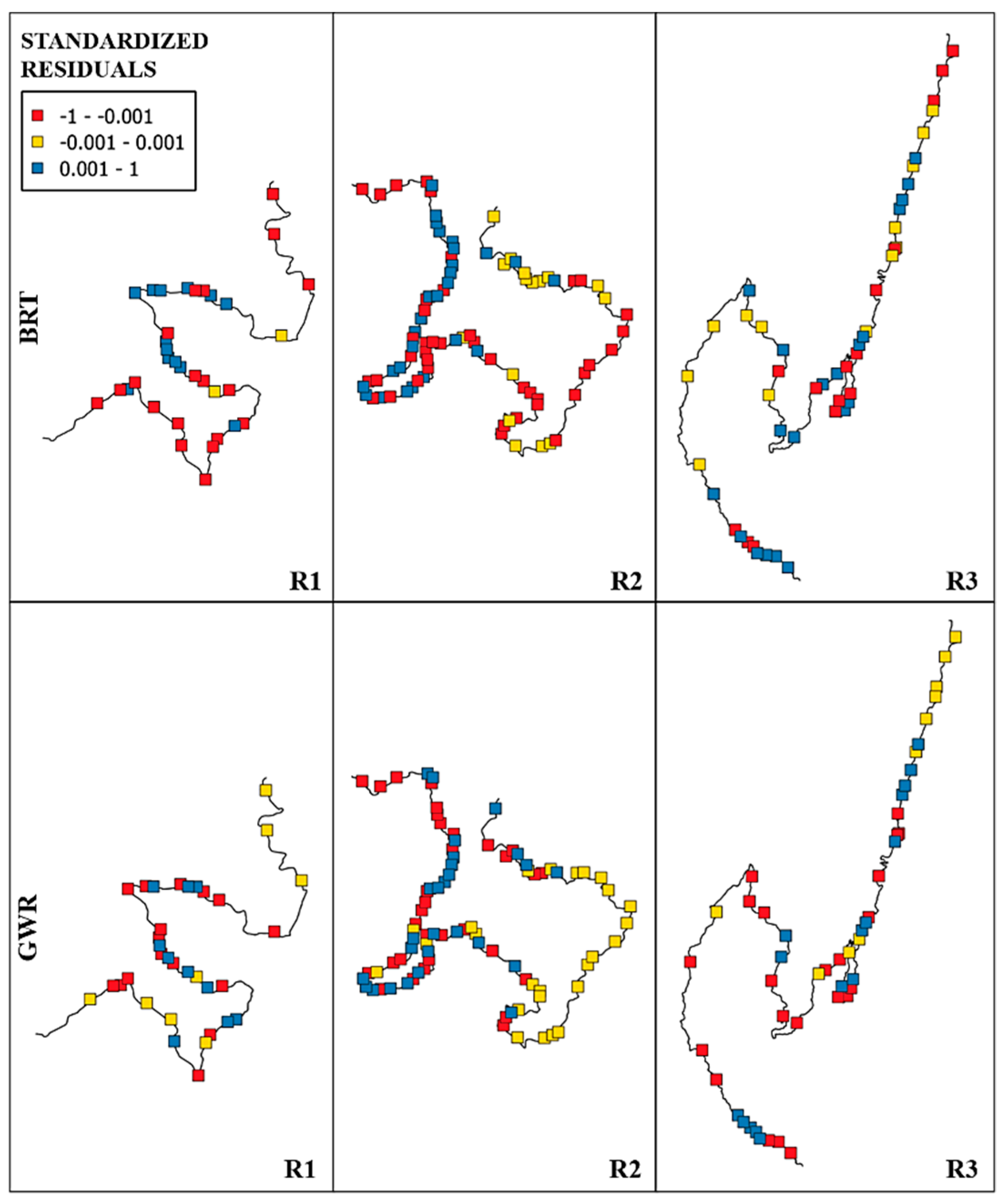
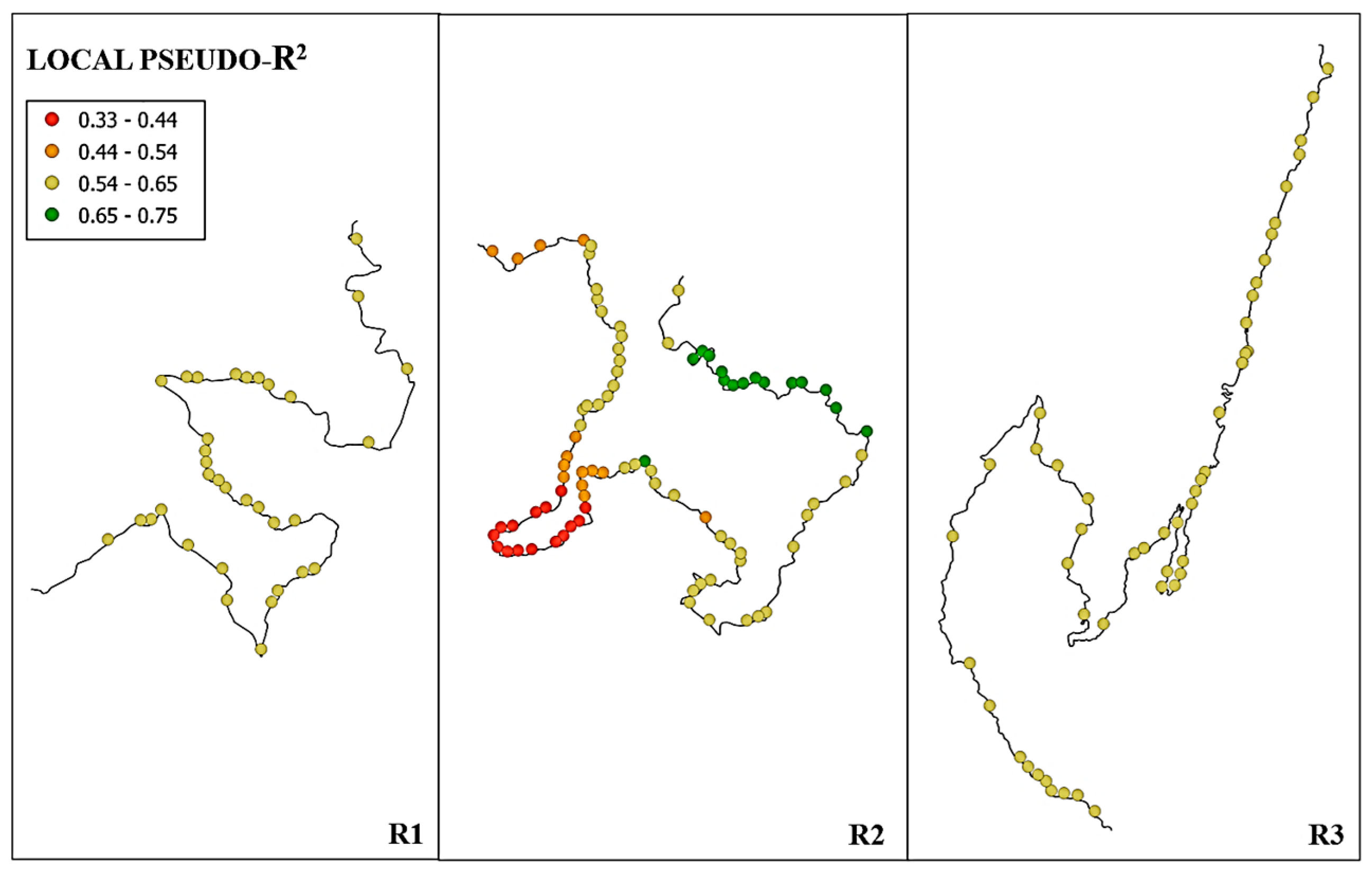
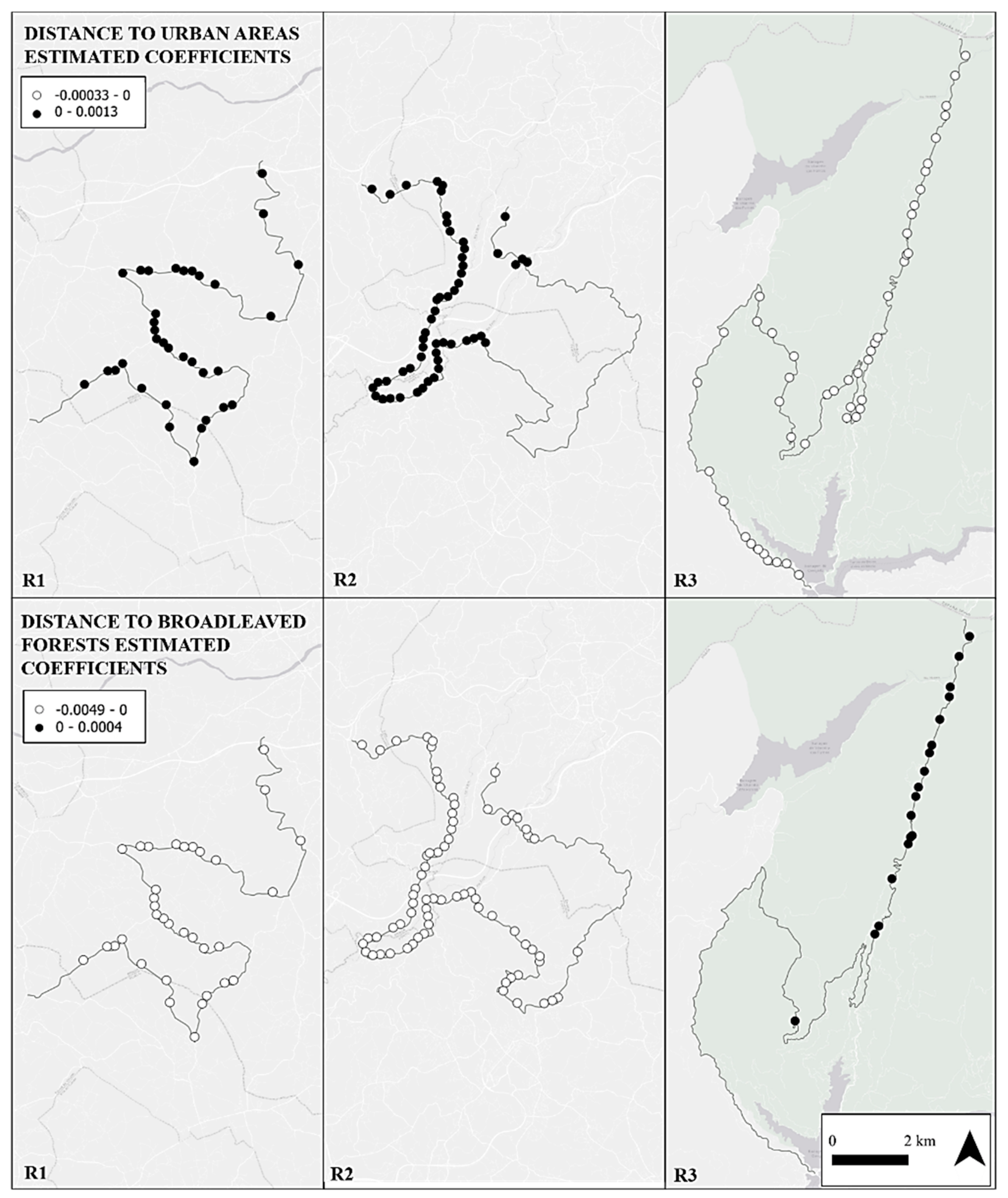
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goosem, M. Fragmentation impacts caused by roads through rainforests. Curr. Sci. 2007, 93, 1587–1595. [Google Scholar]
- Ibisch, P.; Hoffmann, M.T.; Kreft, S.; Pe’er, G.; Kati, V.; Biber-Freudenberger, L.; DellaSala, D.A.; Vale, M.M.; Hobson, P.R.; Selva, N. A global map of roadless areas and their conservation status. Science 2016, 354, 1423–1427. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of Habitat Fragmentation on Biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef]
- Seiler, A. The Toll of the Automobile: Wildlife and Roads in Sweden. Ph.D. Thesis, Swedish University of Agricultural Sciences, Uppsala, Alnarp, Skara, Sweden, 2003. [Google Scholar]
- Ewers, R.M.; Didham, R.K. Confounding factors in the detection of species responses to habitat fragmentation. Biol. Rev. 2006, 81, 117–142. [Google Scholar] [CrossRef] [PubMed]
- Carr, L.W.; Fahrig, L. Effect of road traffic on two amphibian species of differing vagility. Conserv. Biol. 2001, 15, 1071–1078. [Google Scholar] [CrossRef]
- Sillero, N. Amphibian mortality levels on Spanish country roads: Descriptive and spatial analysis. Amphib. Reptil. 2008, 29, 337–347. [Google Scholar] [CrossRef]
- Garriga, N.; Santos, X.; Montori, A.; Richter-Boix, A.; Franch, M.; Llorente, G.A. Are protected areas truly protected? The impact of road traffic on vertebrate fauna. Biodivers. Conserv. 2012, 21, 2761–2774. [Google Scholar] [CrossRef]
- Matos, C.; Sillero, N.; Argana, E. Spatial analysis of Amphibian road mortality levels in northern Portugal country roads. Amphib. Reptil. 2012, 33, 469–483. [Google Scholar] [CrossRef]
- Gibbs, J.P.; Shriver, W.G. Can road mortality limit populations of pool-breeding amphibians? Wetl. Ecol. Manag. 2005, 13, 281–289. [Google Scholar] [CrossRef]
- Lima, S.L.; Blackwell, B.F.; DeVault, T.L.; Fernández-Juricic, E. Animal reactions to oncoming vehicles: A conceptual review. Biol. Rev. 2015, 90, 60–76. [Google Scholar] [CrossRef] [PubMed]
- Alford, R.A.; Richards, S.J. Global amphibian declines: A problem in applied ecology. Annu. Rev. Ecol. Syst. 1999, 30, 133–165. [Google Scholar] [CrossRef]
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.; Fischman, D.L.; Waller, R.W. Status and trends of amphibian declines and extinctions worldwide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef] [PubMed]
- Meijer, J.R.; Huijbregts, M.A.J.; Schotten, K.C.G.J.; Schipper, A.M. Global patterns of current and future road infrastructure. Environ. Res. Lett. 2018, 13, 064006. [Google Scholar] [CrossRef]
- Carvalho, F.; Mira, A. Comparing annual vertebrate road kills over two time periods, 9 years apart: A case study in Mediterranean farmland. Eur. J. Wildl. Res. 2011, 57, 157–174. [Google Scholar] [CrossRef]
- Clevenger, A.P.; Chruszcz, B.; Gunson, K.E. Spatial patterns and factors influencing small vetebrate fauna road-kill aggregations. Biodivers. Conserv. 2003, 109, 15–26. [Google Scholar]
- Seiler, A.; Helldin, J.; Seiler, C. Road mortality in Swedish mammals: Results of a drivers’ questionnaire. Wildl. Biol. 2004, 10, 183–191. [Google Scholar] [CrossRef]
- Seiler, A. Predicting locations of moose-vehicle collisions in Sweden. J. Appl. Ecol. 2005, 2, 371–382. [Google Scholar] [CrossRef]
- Coelho, I.P.; Teixeira, F.Z.; Colombo, P.; Coelho, A.V.; Kindel, A. Anuran road-kills neighboring a peri-urban reserve in the Atlantic Forest, Brazil. J. Environ. Manag. 2012, 112, 17–26. [Google Scholar] [CrossRef]
- Bennett, V.J. Effects of Road Density and Pattern on the Conservation of Species and Biodiversity. Curr. Landsc. Ecol. Rep. 2017, 2, 1–11. [Google Scholar] [CrossRef]
- Mestre, F.; Lopes, H.; Pinto, T.; Sousa, L.G.; Mira, A.; Santos, S.M. Bad moon rising? The influence of the lunar cycle on amphibian roadkills. Eur. J. Wildl. Res. 2019, 65, 58. [Google Scholar] [CrossRef]
- Sillero, N.; Poboljsaj, K.; Lesnik, A.; Salamun, A. Influence of landscape factors on amphibian roadkills at the national level. Diversity 2019, 11, 13. [Google Scholar] [CrossRef]
- D’Amico, M.; Román, J.; de los Reyes, L.; Revilla, E. Vertebrate road-kill patterns in Mediterranean habitats: Who, when and where. Biol. Conserv. 2015, 191, 234–242. [Google Scholar] [CrossRef]
- Ascensão, F.; Kindel, A.; Zimmermann Teixeira, F.; Barrientos, R.; D’Amico, M.; Borda-de-Água, L.; Pereira, H.M. Beware that the lack of wildlife mortality records can mask a serious impact of linear infrastructures. Glob. Ecol. Conserv. 2019, 19, e00661. [Google Scholar] [CrossRef]
- Hasan, M.T.; Sneddon, G.; Ma, R. Modeling binomial amphibian roadkill data in distance sampling while accounting for zero-inflation, serial correlation and varying cluster sizes simultaneously. Environ. Ecol. Stat. 2017, 24, 201–217. [Google Scholar] [CrossRef]
- Lin, Y.P.; Anthony, J.; Lin, W.C.; Lien, W.Y.; Petway, J.R.; Lin, T.E. Spatiotemporal identification of roadkill probability and systematic conservation planning. Landsc. Ecol. 2019, 34, 717–735. [Google Scholar] [CrossRef]
- Petrovan, S.O.; Vale, C.G.; Sillero, N. Using citizen science in road surveys for large-scale amphibian monitoring: Are biased data representative for species distribution? Biodivers. Conserv. 2020, 29, 1767–1781. [Google Scholar] [CrossRef]
- Grilo, C.; Ascensão, F.; Santos-Reis, M.; Bissonette, J.A. Do well-connected landscapes promote road-related mortality? Eur. J. Wildl. Res. 2011, 57, 707–716. [Google Scholar] [CrossRef]
- Sheehan, K.R.; Strager, M.P.; Welsh, S.A. Advantages of geographically weighted regression for modelling benthic substrate in two greater Yellowstone ecosystem streams. Eviron. Model. Assess. 2013, 18, 209–219. [Google Scholar] [CrossRef]
- Graham, H.; Elith, J.; Hijmans, J.; Guisan, A.; Townsend Peterson, A.; Loiselle, B.A.; NCEAS Predicting Species Distributions Working Group. The influence of spatial errors in species occurrence data used in distribution models. J. Appl. Ecol. 2008, 45, 239–247. [Google Scholar] [CrossRef]
- Ali, K.; Partridge, D.; Olfert, M.R. Can geographically weighted regressions improve regional analysis and policy making? Int. Reg. Sci. Rev. 2007, 30, 300–329. [Google Scholar] [CrossRef]
- Gao, J.; Li, S. Detecting spatially non-stationary and scale-dependent relationships between urban landscape fragmentation and related factors using geographically weighted regression. Appl. Geogr. 2011, 31, 292–302. [Google Scholar] [CrossRef]
- Fotheringham, A.S.; Brunsdon, C.; Charlton, M. Geographically Weighted Regression: The Analysis of Spatially Varying Relationships; John Wiley & Sons Inc.: Chichester, UK, 2002. [Google Scholar]
- Kala, A.K.; Tiwari, C.; Mikler, A.R.; Atkinson, S.F. A comparison of least squares regression and geographically weighted regression modeling of West Nile virus risk based on environmental parameters. PeerJ 2017, 5, e3070. [Google Scholar] [CrossRef]
- Erdogan, S. Explorative spatial analysis of traffic accident statistics and road mortality among the provinces of Turkey. J. Saf. Res. 2009, 40, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Robinson, R.M.; Khattak, A.; Wang, X. All Accidents are Not Equal: Using Geographically Weighted Regressions Models to Assess and Forecast Accident Impacts. In Proceedings of the 3rd International Conference on Road Safety and Simulation, Indianapolis, IN, USA, 14–16 September 2011. [Google Scholar]
- Austin, M. Species distribution models and ecological theory: A critical assessment and some possible new approaches. Ecol. Model. 2007, 200, 1–19. [Google Scholar] [CrossRef]
- Kupfer, J.A.; Farris, C.A. Incorporating spatial non-stationarity of regression coefficients into predictive vegetation models. Landsc. Ecol. 2007, 22, 837–852. [Google Scholar] [CrossRef]
- Nogués-Bravo, D. Comparing regression methods to predict species richness patterns. Web Ecol. 2009, 9, 58–67. [Google Scholar] [CrossRef]
- Ye, X.; Yu, X.; Wang, T. Investigating spatial non-stationary environmental effects on the distribution of giant pandas in the Qinling Mountains, China. Glob. Ecol. Conserv. 2020, 21, e00894. [Google Scholar] [CrossRef]
- Loureiro, A.; Ferrand, N.; Carretero, M.A.; Paulo, O. Atlas dos Anfíbios e Répteis de Portugal, 1st ed.; Esfera do Caos: Lisbon, Portugal, 2010. [Google Scholar]
- Ribeiro, R.; Torres, J.; Gomes, V.; Carretero, M.A.; Sillero, N.; Llorente, G.A. Unsuspected richness near home: New herpetological records in Porto Metropolitan Area (NW Portugal). Bol. De La Asoc. Herpetol. Esp. 2010, 21, 27–30. [Google Scholar]
- Orlowski, G. Spatial distribution and seasonal pattern in road mortality of the common toad Bufo bufo in an agricultural landscape of south-western Poland. Amphib. Reptil. 2007, 28, 25–31. [Google Scholar] [CrossRef]
- Glista, D.J.; DeVault, T.L.; DeWoody, J.A. Vertebrate road mortality predominantly impacts amphibians. Herpetol. Conserv. Biol. 2008, 3, 77–87. [Google Scholar]
- Santos, X.; Llorente, G.A.; Montori, A.; Carretero, M.A.; Franch, M.; Garriga, N.; Richter-Boix, A. Evaluating factors affecting amphibian mortality on roads: The case of the Common Toad Bufo bufo, near a breeding place. Anim. Biodivers. Conserv. 2007, 30, 97–104. [Google Scholar]
- Santos, S.M.; Carvalho, F.; Mira, A. How long do the dead survive on the road? Carcass persistence probability and implications for road-kill monitoring surveys. PLoS ONE 2011, 6, e25383. [Google Scholar] [CrossRef] [PubMed]
- Preatoni, D.G.; Tattoni, C.; Bisi, F.; Masseroni, E.; D’Acunto, D.; Lunardi, S.; Grimod, I.; Martinoli, A.; Tosi, G. Open source evaluation of kilometric indexes of abundance. Ecol. Inform. 2012, 7, 35–40. [Google Scholar] [CrossRef]
- Semlitsch, R.D.; Bodie, J.R. Biological criteria for buffer zones around wetlands and riparian habitats for amphibians and reptiles. Conserv. Biol. 2003, 17, 1219–1228. [Google Scholar] [CrossRef]
- Smith, A.M.; Green, D.M. Dispersal and the metapopulation paradigm in amphibian ecology and conservation: Are all amphibian populations metapopulations? Ecography 2005, 28, 110–128. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Field, A.; Miles, J.; Field, Z. Discovering Statistics Using R; Sage Publications Ltd.: London, UK, 2012. [Google Scholar]
- Breiman, L. Random forest. Mach. Learn. 1999, 45, 1–35. [Google Scholar]
- Elith, J.; Leathwick, J.R.; Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef]
- Miller, J.A. Species distribution models spatial autocorrelation and non-stationarity. Prog. Phys. Geogr. 2012, 36, 681692. [Google Scholar] [CrossRef]
- Mellin, C.; Mengersen, K.; Bradshaw, C.J.A.; Caley, M.J. Generalizing the use of geographical weights in biodiversity modelling. Glob. Ecol. Biogeogr. 2014, 23, 1314–1323. [Google Scholar] [CrossRef]
- Ascensão, F.; Desbiez, A.L.J.; Medici, E.P.; Bager, A. Spatial patterns of road mortality of medium-large mammals in Mato Grosso do Sul, Brazil. Wildl. Res. 2017, 44, 135–146. [Google Scholar] [CrossRef]
- Oshan, T.M.; Li, Z.; Kang, W.; Wolf, L.J.; Fotheringham, A.S. mgwr: A Python implementation of multiscale geographically weighted regression for investigating process spatial heterogeneity and scale. ISPRS Int. J. Geo-Inf. 2019, 8, 269. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, P.R.; Dudík, M.; Ferrier, S.; Guisan, A.; JHijmans, R.; Huettmann, F.; RLeathwick, J.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Willmott, C.J.; Robeson, S.M.; Matsuura, K. A refined index of model performance. Int. J. Climatol. 2011, 32, 2088–2094. [Google Scholar] [CrossRef]
- Zou, K.H.; O’Malley, A.J.; Mauri, L. Receiver-operating characteristic analysis for evaluating diagnostic tests and predictive models. Circulation 2007, 115, 654–657. [Google Scholar] [CrossRef]
- Windle, M.J.S.; Rose, G.A.; Devillers, R.; Fortin, M.-J. Exploring spatial non-stationarity of fisheries survey data using geographically weighted regression (GWR): An example from the Northwest Atlantic. ICES J. Mar. Sci. 2010, 67, 145–154. [Google Scholar] [CrossRef]
- Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Qiao, H.; Soberón, J.; Peterson, A.T. No silver bullets in correlative ecological niche modelling: Insights from testing among many potential algorithms for niche estimation. Methods Ecol. Evol. 2015, 6, 1126–1136. [Google Scholar] [CrossRef]
- Lopatin, J.; Dolos, K.; Hernández, H.J.; Galleguillos, M.; Fassnacht, F.E. Comparing generalized linear models and random forest to model vascular plant species richness using LiDAR data in a natural forest in central Chile. Remote Sens. Environ. 2016, 173, 200–210. [Google Scholar] [CrossRef]
- Gu, C.; Wahba, G. Minimizing GCV/GML scores with multiple smoothing parameters via the Newton method. SIAM J. Sci. Stat. Comput. 1991, 12, 383–398. [Google Scholar] [CrossRef]
- Li, M.; Zhang, C.; Xu, B.; Xue, Y.; Ren, Y. A comparison of GAM and GWR in modelling spatial distribution of Japanese mantis shrimp (Oratosquilla oratoria) in coastal waters. Estuar. Coast. Shelf Sci. 2020, 244, 106928. [Google Scholar] [CrossRef]
- Wright, P.G.R.; Coomber, F.G.; Bellamy, C.C.; Perkins, S.E.; Mathews, F. Predicting hedgehog mortality risks on British roads using habitat suitability modelling. PeerJ 2020, 7, e8154. [Google Scholar] [CrossRef] [PubMed]
- Grilo, C.; Koroleva, E.; Andrášik, R.; Bíl, M.; González-Suárez, M. Roadkill risk and population vulnerability in European birds and mammals. Front. Ecol. Environ. 2020, 18, 323–328. [Google Scholar] [CrossRef]
- Goovaerts, P. Geostatistical analysis of health data: State-of-the-art and perspectives. In GeoENV VI—Geostatistics for Environmental Applications (Quantitative Geology and Geostatistics); Soares, A., Pereira, M.J., Dimitrakopoulos, R., Eds.; Springer: Dordrecht, The Netherlands, 2008; Volume 15. [Google Scholar]
- Osborne, P.E.; Foody, G.M.; Suárez-Seoane, S. Non-stationarity and local approaches to modelling the distributions of wildlife. Divers. Distrib. 2007, 13, 313–323. [Google Scholar] [CrossRef]
- INE. Estatísticas dos Transportes e Comunicações 2019—Instituto Nacional de Estatística (I.N.E); INE: Lisboa, Portugal, 2020; p. 106. [Google Scholar]
- Baptista, N. Amphibian Roadkills: Hotspot Analysis and Locations of Amphibian Underpasses Using Gorelick’s Index. Master’s Thesis, University of Évora, Largo dos Colegiais, Portugal, 2006. [Google Scholar]
- Garriga, N.; Franch, M.; Santos, X.; Montori, A.; Llorente, G.A. Seasonal variation in vertebrate traffic casualties and its implications for mitigation measures. Landsc. Urban Plan. 2017, 157, 36–44. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Description | Unit | Source |
|---|---|---|---|
| Distance to urban areas | Linear distance to the closest urban and artificial surface | m | Corine Land Cover (v2012) |
| Distance to irrigated land | Linear distance to the closest permanently irrigated land | m | Corine Land Cover (v2012) |
| Distance to agricultural areas | Linear distance to the closest agricultural areas, mainly composed of vineyards, pastures, annual crops, and complex cultivation patterns | m | Corine Land Cover (v2012) |
| Distance to broadleaved forests | Linear distance to the closest broad-leaved forest, mainly composed of trees, some shrubs, and bushes | m | Corine Land Cover (v2012) |
| Distance to coniferous forests | Linear distance to the closest coniferous forest, mainly composed of trees, shrubs, and bushes | m | Corine Land Cover (v2012) |
| Distance to mixed forests | Linear distance to the closest mixed forest | m | Corine Land Cover (v2012) |
| Distance to shrubs | Linear distance to scrub or herbaceous vegetation associations, mainly composed of natural grasslands, moors and heathland, and transitional woodland shrub | m | Corine Land Cover (v2012) |
| Distance to open areas | Linear distance to open spaces with little or no vegetation, composed of bare rocks, sparsely vegetated areas, and burnt areas | m | Corine Land Cover (v2012) |
| Distance to water bodies | Linear distance to the closest water body | m | Corine Land Cover (v2012) |
| Fires | Frequency of forest fires between 1975 and 2013 | - | ICNF, Territórios ardidos 1975–2013 |
| Slope | - | Sistema Nacional de Informação Geográfica—DGT |
| ROAD | Location | Road Segments | Segments with Road-Kills | Road-Kills | Mean AKI (ind/km) |
|---|---|---|---|---|---|
| R1 | Barcelos | 111 (27.70 km) | 17 | 89 | 0.54 |
| R2 | Guimarães | 143 (35.70 km) | 42 | 171 | 0.80 |
| R3 | Gerês | 157 (39.10 km) | 24 | 83 | 0.35 |
| TOTAL = | 411 (102.50 km) | 83 | 343 | MEAN = 0.56 | |
| Methods | Index of Agreement | AUC | Deviance Explained (%) |
|---|---|---|---|
| GLM | 0.567 (0.554–0.586) | 0.812 (0.787–0.831) | 22.9 (21.0–27.0) |
| GAM | 0.635 (0.618–0.655) | 0.843 (0.822–0.871) | 48.4 (41.7–53.1) |
| RF | 0.593 (0.577–0.610) | 0.817 (0.794–0.840) | - |
| BRT | 0.755 (0.699–0.792) | 0.933 (0.900–0.961) | 70.8 (61.8–76.6) |
| GWR | 0.700 (0.675–0.719) | 0.881 (0.860–0.902) | 61.9 (55.3–66.7) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sousa-Guedes, D.; Franch, M.; Sillero, N. A Spatial Approach for Modeling Amphibian Road-Kills: Comparison of Regression Techniques. ISPRS Int. J. Geo-Inf. 2021, 10, 343. https://doi.org/10.3390/ijgi10050343
Sousa-Guedes D, Franch M, Sillero N. A Spatial Approach for Modeling Amphibian Road-Kills: Comparison of Regression Techniques. ISPRS International Journal of Geo-Information. 2021; 10(5):343. https://doi.org/10.3390/ijgi10050343
Chicago/Turabian StyleSousa-Guedes, Diana, Marc Franch, and Neftalí Sillero. 2021. "A Spatial Approach for Modeling Amphibian Road-Kills: Comparison of Regression Techniques" ISPRS International Journal of Geo-Information 10, no. 5: 343. https://doi.org/10.3390/ijgi10050343
APA StyleSousa-Guedes, D., Franch, M., & Sillero, N. (2021). A Spatial Approach for Modeling Amphibian Road-Kills: Comparison of Regression Techniques. ISPRS International Journal of Geo-Information, 10(5), 343. https://doi.org/10.3390/ijgi10050343