Axin Family of Scaffolding Proteins in Development: Lessons from C. elegans


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
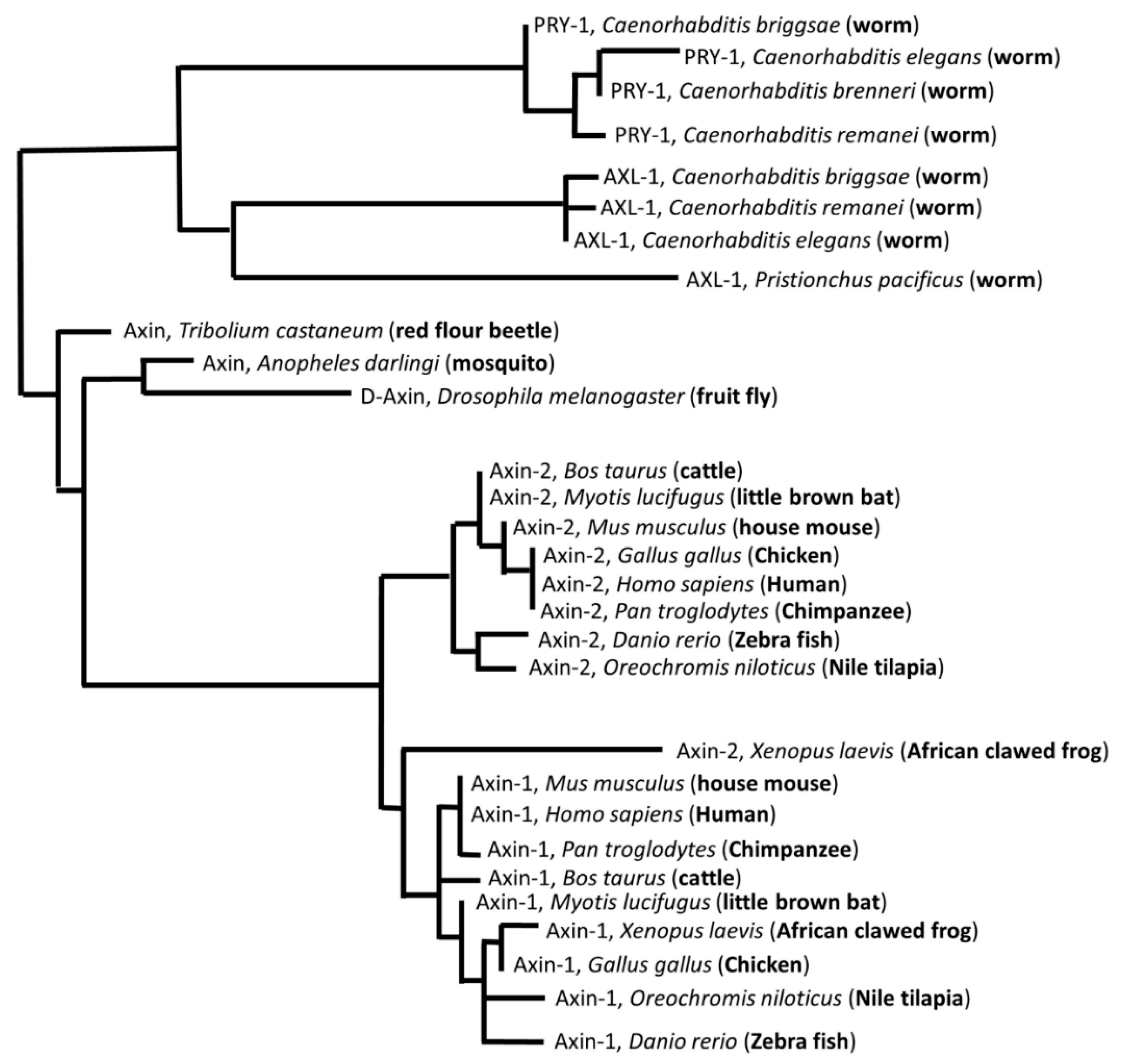
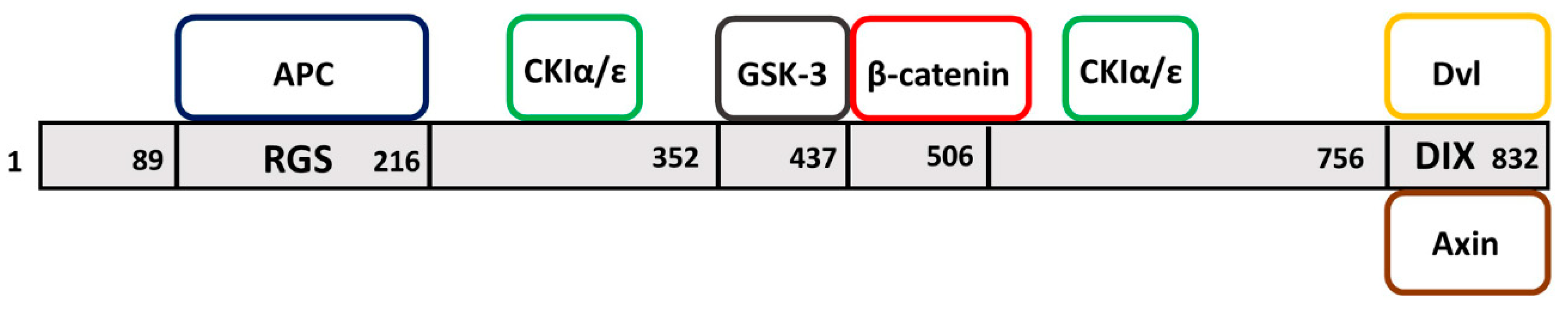
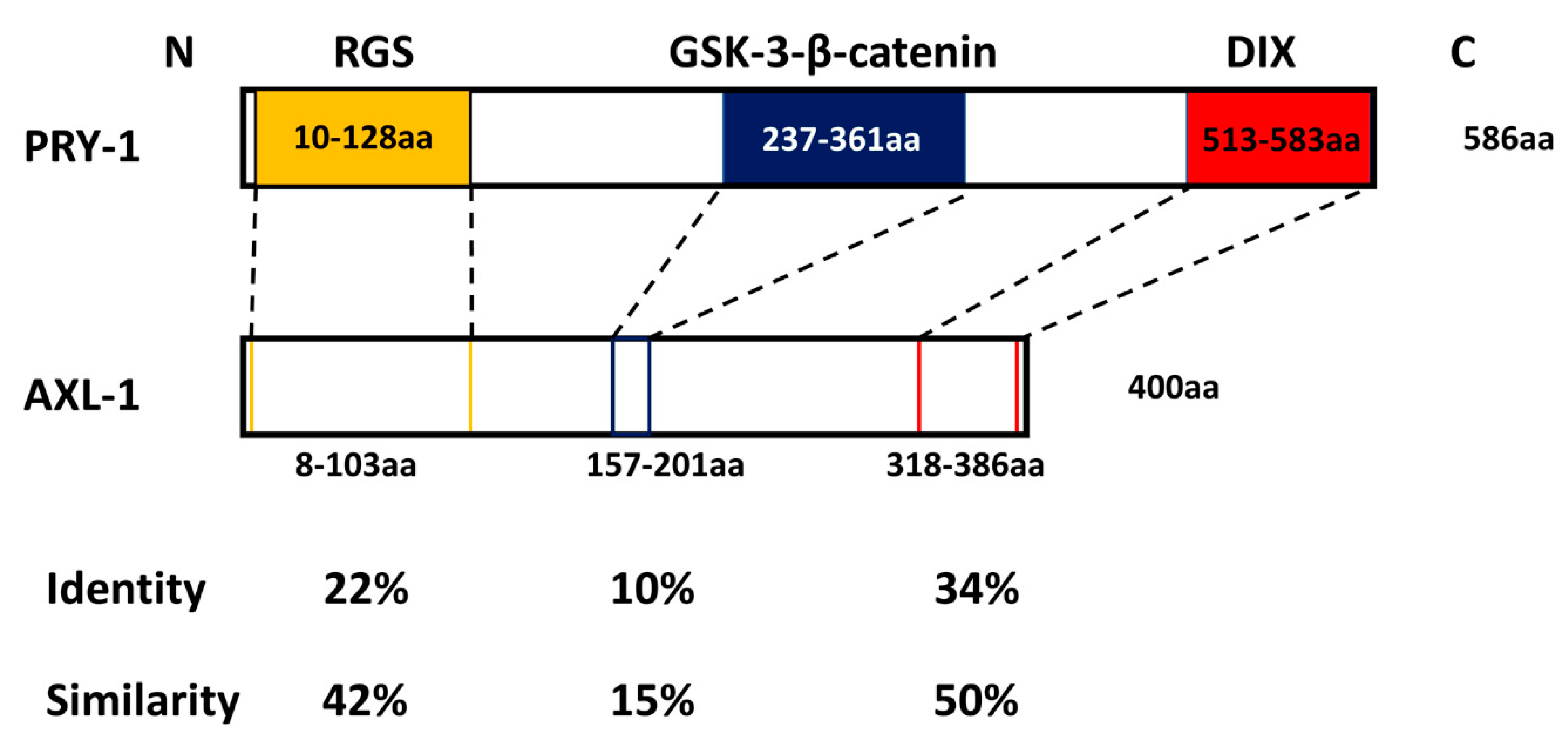
:1. Axin Domains
2. Overview of the Developmental Roles of the Axin Family
2.1. Vertebrate Models
2.2. Invertebrate Models
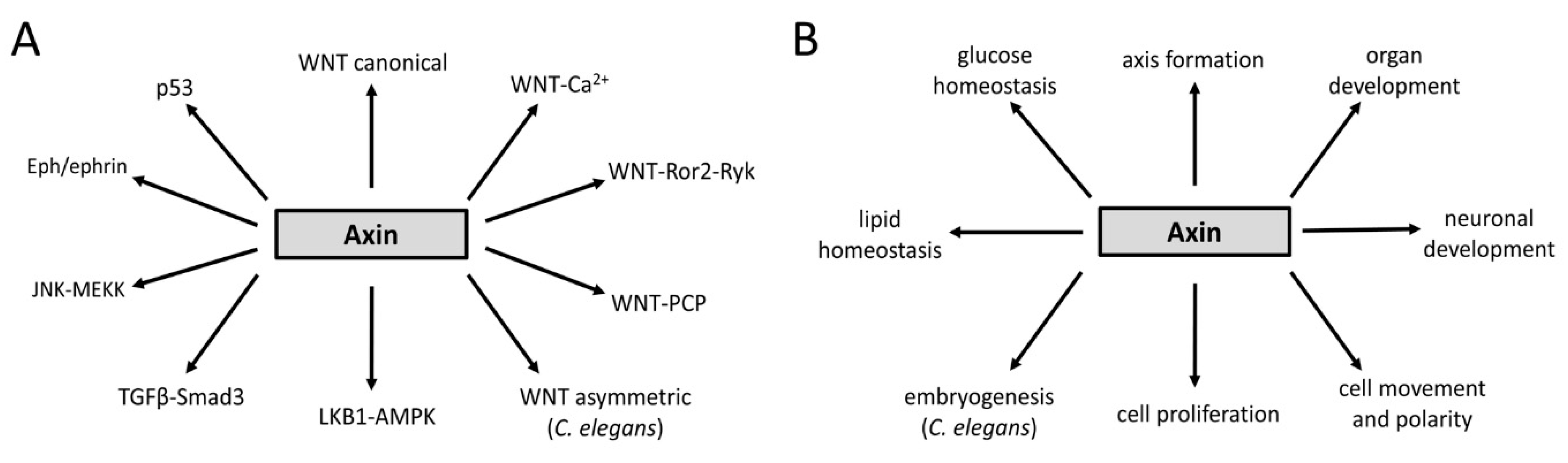
3. Axin Proteins Interact with Many Factors Including Signaling Pathway Components
4. Regulation of Axin Functions
5. Regulation of Developmental Processes in C. elegans
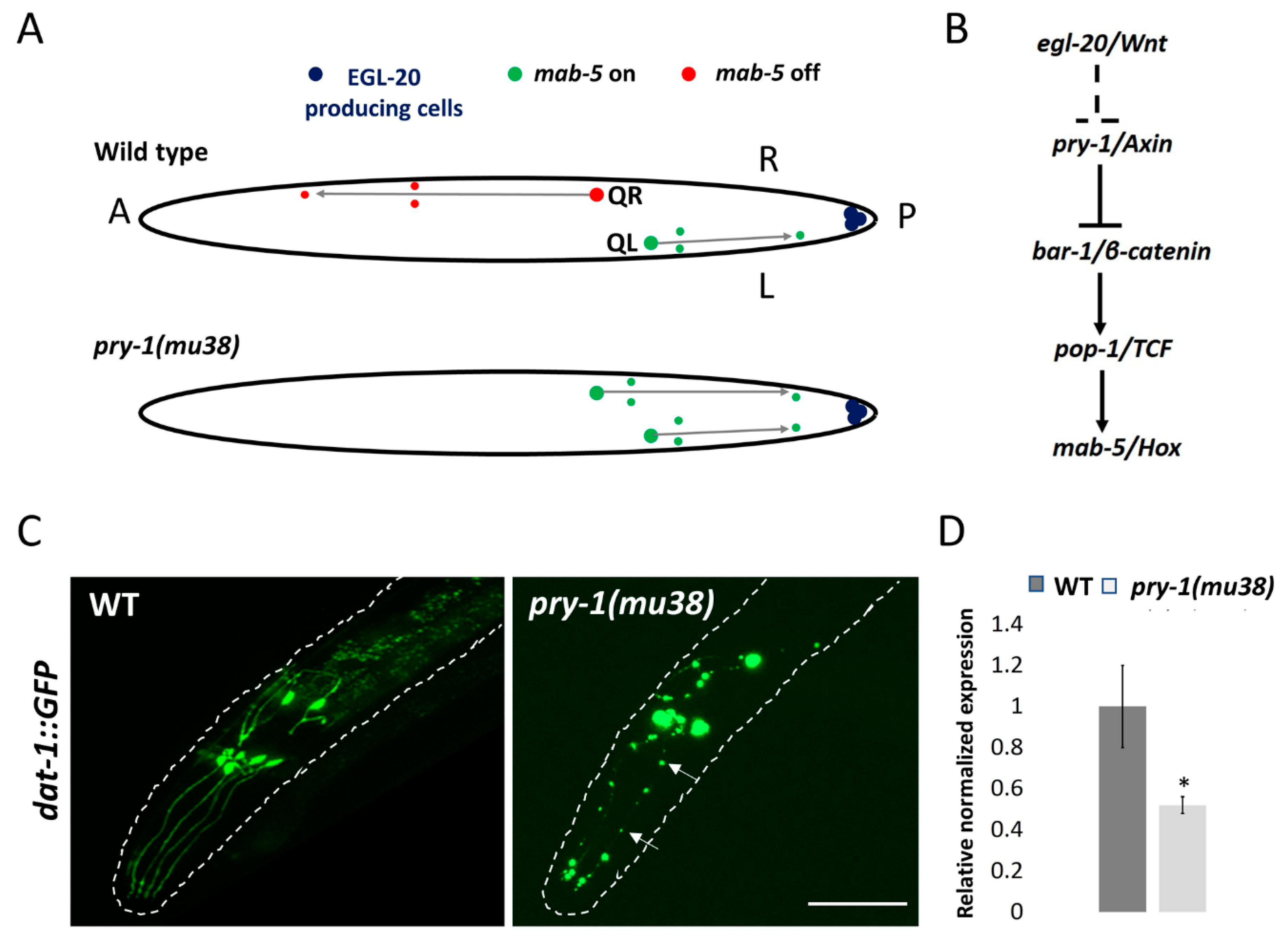
5.1. Neuronal Development
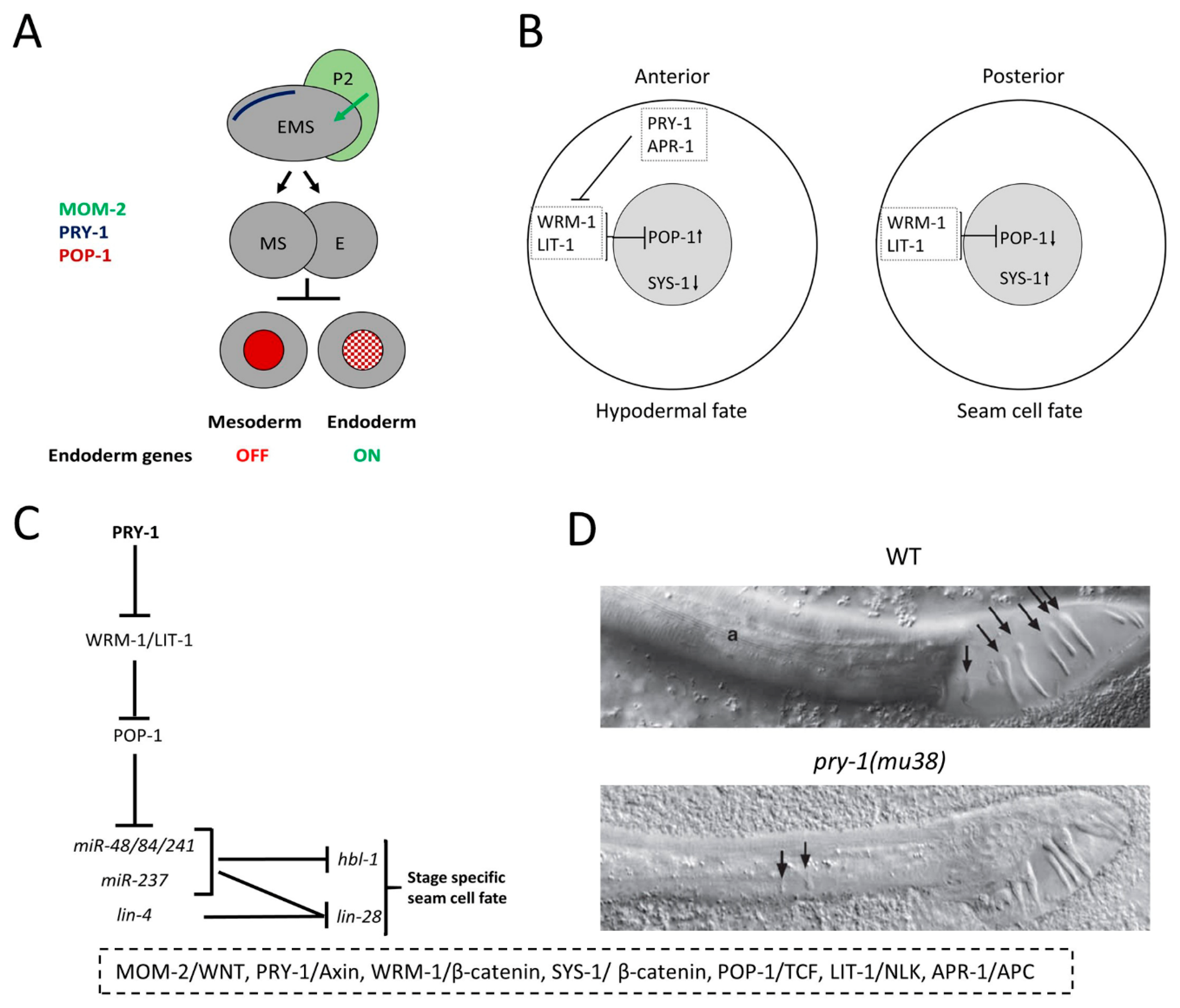
5.2. Embryogenesis
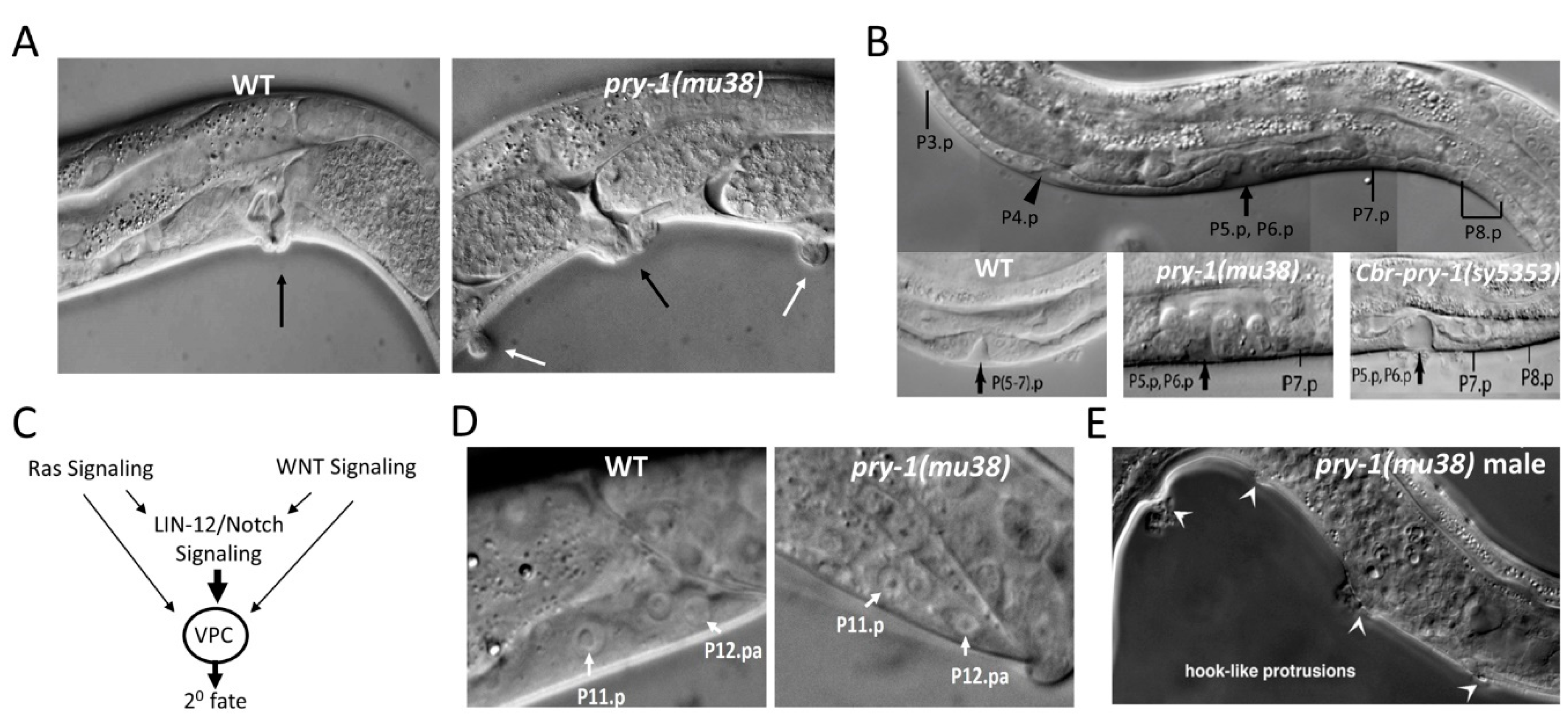
5.3. Seam Cell Development
5.4. Vulva Development
5.5. P11/12 Development
5.6. Male Hook Development
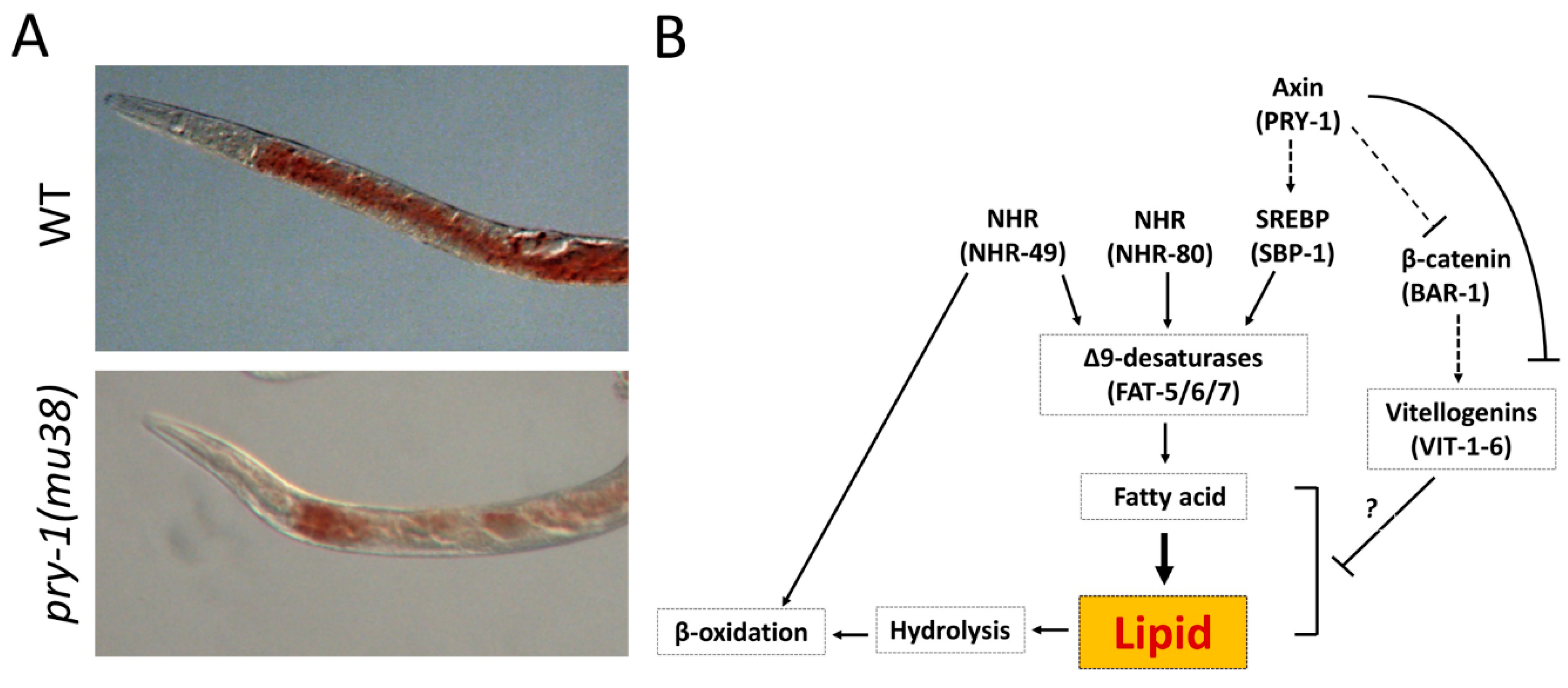
5.7. Lipid Metabolism
6. Major Findings from C. elegans Studies
7. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Zeng, L.; Fagotto, F.; Zhang, T.; Hsu, W.; Vasicek, T.J.; Perry, W.L., 3rd; Lee, J.J.; Tilghman, S.M.; Gumbiner, B.M.; Costantini, F. The mouse Fused locus encodes Axin, an inhibitor of the Wnt signaling pathway that regulates embryonic axis formation. Cell 1997, 90, 181–192. [Google Scholar] [CrossRef]
- Gluecksohn-Schoenheimer, S. The effects of a lethal mutation responsible for duplications and twinning in mouse embryos. J. Exp. Zool. 1949, 110, 47–76. [Google Scholar] [CrossRef]
- Jacobs-Cohen, R.J.; Spiegelman, M.; Cookingham, J.C.; Bennett, D. Knobbly, a new dominant mutation in the mouse that affects embryonic ectoderm organization. Genet. Res. 1984, 43, 43–50. [Google Scholar] [CrossRef]
- Perry, W.L., 3rd; Vasicek, T.J.; Lee, J.J.; Rossi, J.M.; Zeng, L.; Zhang, T.; Tilghman, S.M.; Costantini, F. Phenotypic and molecular analysis of a transgenic insertional allele of the mouse Fused locus. Genetics 1995, 141, 321–332. [Google Scholar]
- Ikeda, S.; Kishida, S.; Yamamoto, H.; Murai, H.; Koyama, S.; Kikuchi, A. Axin, a negative regulator of the Wnt signaling pathway, forms a complex with GSK-3beta and beta-catenin and promotes GSK-3beta-dependent phosphorylation of beta-catenin. EMBO J. 1998, 17, 1371–1384. [Google Scholar] [CrossRef]
- Yamamoto, H.; Kishida, S.; Uochi, T.; Ikeda, S.; Koyama, S.; Asashima, M.; Kikuchi, A. Axil, a member of the Axin family, interacts with both glycogen synthase kinase 3beta and beta-catenin and inhibits axis formation of Xenopus embryos. Mol. Cell. Biol. 1998, 18, 2867–2875. [Google Scholar] [CrossRef]
- Mao, J.; Wang, J.; Liu, B.; Pan, W.; Farr, G.H., 3rd; Flynn, C.; Yuan, H.; Takada, S.; Kimelman, D.; Li, L.; et al. Low-density lipoprotein receptor-related protein-5 binds to Axin and regulates the canonical Wnt signaling pathway. Mol. Cell 2001, 7, 801–809. [Google Scholar] [CrossRef]
- Wehrli, M.; Dougan, S.T.; Caldwell, K.; O’Keefe, L.; Schwartz, S.; Vaizel-Ohayon, D.; Schejter, E.; Tomlinson, A.; Dinardo, S. arrow encodes an LDL-receptor-related protein essential for Wingless signalling. Nature 2000, 407, 527–530. [Google Scholar] [CrossRef]
- Tolwinski, N.S.; Wehrli, M.; Rives, A.; Erdeniz, N.; Dinardo, S.; Wieschaus, E. Wg/Wnt signal can be transmitted through arrow/LRP5,6 and Axin independently of Zw3/Gsk3beta activity. Dev. Cell 2003, 4, 407–418. [Google Scholar] [CrossRef]
- Yamamoto, H.; Kishida, S.; Kishida, M.; Ikeda, S.; Takada, S.; Kikuchi, A. Phosphorylation of Axin, a Wnt Signal Negative Regulator, by Glycogen Synthase Kinase-3β Regulates Its Stability. J. Boil. Chem. 1999, 274, 10681–10684. [Google Scholar] [CrossRef]
- Kishida, S.; Yamamoto, H.; Ikeda, S.; Kishida, M.; Sakamoto, I.; Koyama, S.; Kikuchi, A. Axin, a negative regulator of the wnt signaling pathway, directly interacts with adenomatous polyposis coli and regulates the stabilization of beta-catenin. J. Boil. Chem. 1998, 273, 10823–10826. [Google Scholar] [CrossRef]
- Hart, M.J.; Santos, R.D.L.; Albert, I.N.; Rubinfeld, B.; Polakis, P. Downregulation of beta-catenin by human Axin and its association with the APC tumor suppressor, beta-catenin and GSK3 beta. Curr. Boil. 1998, 8, 573–581. [Google Scholar] [CrossRef]
- Nakamura, T.; Hamada, F.; Ishidate, T.; Anai, K.-I.; Kawahara, K.; Toyoshima, K.; Akiyama, T. Axin, an inhibitor of the Wnt signalling pathway, interacts with beta-catenin, GSK-3beta and APC and reduces the beta-catenin level. Genes Cells 1998, 3, 395–403. [Google Scholar] [CrossRef]
- Hsu, W.; Zeng, L.; Costantini, F. Identification of a Domain of Axin That Binds to the Serine/Threonine Protein Phosphatase 2A and a Self-binding Domain. J. Boil. Chem. 1999, 274, 3439–3445. [Google Scholar] [CrossRef] [Green Version]
- Schwarz-Romond, T.; Asbrand, C.; Bakkers, J.; Kuhl, M.; Schaeffer, H.-J.; Huelsken, J.; Behrens, J.; Hammerschmidt, M.; Birchmeier, W. The ankyrin repeat protein Diversin recruits Casein kinase Iε to the β-catenin degradation complex and acts in both canonical Wnt and Wnt/JNK signaling. Genome Res. 2002, 16, 2073–2084. [Google Scholar]
- Shiomi, K.; Uchida, H.; Keino-Masu, K.; Masu, M. Ccd1, a novel protein with a DIX domain, is a positive regulator in the Wnt signaling during zebrafish neural patterning. Curr. Boil. 2003, 13, 73–77. [Google Scholar] [CrossRef]
- Li, L.; Yuan, H.; Weaver, C.D.; Mao, J.; Farr, G.H., 3rd; Sussman, D.J.; Jonkers, J.; Kimelman, D.; Wu, D. Axin and Frat1 interact with dvl and GSK, bridging Dvl to GSK in Wnt-mediated regulation of LEF-1. EMBO J. 1999, 18, 4233–4240. [Google Scholar] [CrossRef]
- Rubinfeld, B.; Tice, D.A.; Polakis, P. Axin-dependent Phosphorylation of the Adenomatous Polyposis Coli Protein Mediated by Casein Kinase 1epsilon. J. Boil. Chem. 2001, 276, 39037–39045. [Google Scholar] [CrossRef]
- Hedgepeth, C.M.; Deardorff, M.A.; Rankin, K.; Klein, P.S. Regulation of Glycogen Synthase Kinase 3β and Downstream Wnt Signaling by Axin. Mol. Cell. Boil. 1999, 19, 7147–7157. [Google Scholar] [CrossRef]
- Hamada, F.; Tomoyasu, Y.; Takatsu, Y.; Nakamura, M.; Nagai, S.-I.; Suzuki, A.; Fujita, F.; Shibuya, H.; Toyoshima, K.; Ueno, N.; et al. Negative Regulation of Wingless Signaling by D-Axin, a Drosophila Homolog of Axin. Science 1999, 283, 1739–1742. [Google Scholar] [CrossRef]
- Oosterveen, T.; Coudreuse, D.Y.; Yang, P.-T.; Fraser, E.; Bergsma, J.; Dale, T.C.; Korswagen, H.C. Two functionally distinct Axin-like proteins regulate canonical Wnt signaling in C. elegans. Dev. Boil. 2007, 308, 438–448. [Google Scholar] [CrossRef]
- Behrens, J.; Jerchow, B.A.; Würtele, M.; Grimm, J.; Asbrand, C.; Wirtz, R.; Kühl, M.; Wedlich, D.; Birchmeier, W. Functional interaction of an axin homolog, conductin, with beta-catenin, APC, and GSK3beta. Science 1998, 280, 596–599. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.-M.I.; Jerchow, B.; Sheu, T.-J.; Liu, B.; Costantini, F.; Puzas, J.E.; Birchmeier, W.; Hsu, W. The role of Axin2 in calvarial morphogenesis and craniosynostosis. Development 2005, 132, 1995–2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, L.; Mahaffey, J.P.; Alcorn, H.L.; Anderson, K.V. Tissue-specific roles of Axin2 in the inhibition and activation of Wnt signaling in the mouse embryo. Proc. Natl. Acad. Sci. USA 2011, 108, 8692–8697. [Google Scholar] [CrossRef] [Green Version]
- Jho, E.-H.; Zhang, T.; Domon, C.; Joo, C.-K.; Freund, J.-N.; Costantini, F. Wnt/β-Catenin/Tcf Signaling Induces the Transcription of Axin2, a Negative Regulator of the Signaling Pathway. Mol. Cell. Boil. 2002, 22, 1172–1183. [Google Scholar] [CrossRef]
- Lustig, B.; Jerchow, B.; Sachs, M.; Weiler, S.; Pietsch, T.; Karsten, U.; Van De Wetering, M.; Clevers, H.; Schlag, P.M.; Birchmeier, W.; et al. Negative Feedback Loop of Wnt Signaling through Upregulation of Conductin/Axin2 in Colorectal and Liver Tumors. Mol. Cell. Boil. 2002, 22, 1184–1193. [Google Scholar] [CrossRef] [Green Version]
- Hinoi, T.; Yamamoto, H.; Kishida, M.; Takada, S.; Kishida, S.; Kikuchi, A. Complex Formation of Adenomatous Polyposis Coli Gene Product and Axin Facilitates Glycogen Synthase Kinase-3β-dependent Phosphorylation of β-Catenin and Down-regulates β-Catenin. J. Boil. Chem. 2000, 275, 34399–34406. [Google Scholar] [CrossRef]
- Kishida, M.; Koyama, S.; Kishida, S.; Matsubara, K.; Nakashima, S.; Higano, K.; Takada, R.; Takada, S.; Kikuchi, A. Axin prevents Wnt-3a-induced accumulation of β-catenin. Oncogene 1999, 18, 979–985. [Google Scholar] [CrossRef]
- Dajani, R.; Fraser, E.; Roe, S.; Yeo, M.; Good, V.M.; Thompson, V.; Dale, T.C.; Pearl, L.H. Structural basis for recruitment of glycogen synthase kinase 3β to the axin–APC scaffold complex. EMBO J. 2003, 22, 494–501. [Google Scholar] [CrossRef]
- Zhang, Y.; Qiu, W.-J.; Chan, S.C.; Han, J.; He, X.; Lin, S.-C. Casein Kinase I and Casein Kinase II Differentially Regulate Axin Function in Wnt and JNK Pathways. J. Boil. Chem. 2002, 277, 17706–17712. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Li, Y.; Semënov, M.; Han, C.; Baeg, G.-H.; Tan, Y.; Zhang, Z.; Lin, X.; He, X. Control of β-Catenin Phosphorylation/Degradation by a Dual-Kinase Mechanism. Cell 2002, 108, 837–847. [Google Scholar] [CrossRef]
- Li, V.S.; Ng, S.S.; Boersema, P.J.; Low, T.Y.; Karthaus, W.R.; Gerlach, J.P.; Mohammed, S.; Heck, A.J.; Maurice, M.M.; Mahmoudi, T.; et al. Wnt Signaling through Inhibition of β-Catenin Degradation in an Intact Axin1 Complex. Cell 2012, 149, 1245–1256. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Zou, H.; Jin, L.; Lin, S.; Li, Q.; Ye, Z.; Rui, H.; Lin, S.C. Axin contains three separable domains that confer intramolecular, homodimeric, and heterodimeric interactions involved in distinct functions. J. Biol. Chem. 2005, 280, 5054–5060. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Lin, S.-C. Axin: A Master Scaffold for Multiple Signaling Pathways. Neurosignals 2004, 13, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Figeac, N.; Zammit, P.S. Coordinated action of Axin1 and Axin2 suppresses β-catenin to regulate muscle stem cell function. Cell Signal. 2015, 27, 1652–1665. [Google Scholar] [CrossRef] [PubMed]
- Huraskin, D.; Eiber, N.; Reichel, M.; Zidek, L.M.; Kravic, B.; Bernkopf, D.; Von Maltzahn, J.; Behrens, J.; Hashemolhosseini, S. Wnt/β-catenin signaling via Axin2 is required for myogenesis and, together with YAP/Taz and Tead1, active in IIa/IIx muscle fibers. Development 2016, 143, 3128–3142. [Google Scholar] [CrossRef]
- Carl, M.; Bianco, I.H.; Bajoghli, B.; Aghaallaei, N.; Czerny, T.; Wilson, S.W. Wnt/Axin1/beta-catenin signaling regulates asymmetric nodal activation, elaboration, and concordance of CNS asymmetries. Neuron 2007, 55, 393–405. [Google Scholar] [CrossRef]
- Heisenberg, C.-P.; Houart, C.; Take-Uchi, M.; Rauch, G.-J.; Young, N.; Coutinho, P.; Masai, I.; Caneparo, L.; Concha, M.L.; Geisler, R.; et al. A mutation in the Gsk3–binding domain of zebrafish Masterblind/Axin1 leads to a fate transformation of telencephalon and eyes to diencephalon. Genome Res. 2001, 15, 1427–1434. [Google Scholar] [CrossRef]
- Kofron, M.; Klein, P.; Zhang, F.; Houston, D.W.; Schaible, K.; Wylie, C.; Heasman, J. The Role of Maternal Axin in Patterning the Xenopus Embryo. Dev. Boil. 2001, 237, 183–201. [Google Scholar] [CrossRef] [Green Version]
- Ye, T.; Fu, A.K.Y.; Ip, N.Y. Emerging roles of Axin in cerebral cortical development. Front. Cell. Neurosci. 2015, 9, 217. [Google Scholar] [CrossRef]
- He, C.-W.; Liao, C.-P.; Pan, C.-L. Wnt signalling in the development of axon, dendrites and synapses. Open Boil. 2018, 8, 180116. [Google Scholar] [CrossRef] [PubMed]
- Lyu, J.; Costantini, F.; Jho, E.-H.; Joo, C.-K. Ectopic Expression of Axin Blocks Neuronal Differentiation of Embryonic Carcinoma P19 Cells. J. Boil. Chem. 2003, 278, 13487–13495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orme, M.H.; Giannini, A.L.; Vivanco, M.D.; Kypta, R.M. Glycogen synthase kinase-3 and Axin function in a beta-catenin-independent pathway that regulates neurite outgrowth in neuroblastoma cells. Mol. Cell. Neurosci. 2003, 24, 673–686. [Google Scholar] [CrossRef]
- Endo, Y.; Beauchamp, E.; Woods, D.; Taylor, W.G.; Toretsky, J.A.; Üren, A.; Rubin, J.S. Wnt-3a and Dickkopf-1 Stimulate Neurite Outgrowth in Ewing Tumor Cells via a Frizzled3- and c-Jun N-Terminal Kinase-Dependent Mechanism. Mol. Cell. Boil. 2008, 28, 2368–2379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, W.-Q.; Ip, J.P.K.; Li, R.; Ng, Y.P.; Lin, S.-C.; Chen, Y.; Fu, A.K.Y.; Ip, N.Y. Cdk5-Mediated Phosphorylation of Axin Directs Axon Formation during Cerebral Cortex Development. J. Neurosci. 2011, 31, 13613–13624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, W.-Q.; Chen, W.-W.; Fu, A.K.; Ip, N.Y. Axin Directs the Amplification and Differentiation of Intermediate Progenitors in the Developing Cerebral Cortex. Neuron 2013, 79, 665–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Fu, A.K.; Ip, N.Y. Axin: An emerging key scaffold at the synapse. IUBMB Life 2013, 65, 685–691. [Google Scholar] [CrossRef]
- Chen, Y.; Liang, Z.; Fei, E.; Chen, Y.; Zhou, X.; Fang, W.; Fu, W.-Y.; Fu, A.K.Y.; Ip, N.Y. Axin Regulates Dendritic Spine Morphogenesis through Cdc42-Dependent Signaling. PLoS ONE 2015, 10, e0133115. [Google Scholar] [CrossRef]
- Peifer, M. Wnt Signaling in Oncogenesis and Embryogenesis—A Look Outside the Nucleus. Science 2000, 287, 1606–1609. [Google Scholar] [CrossRef]
- Jin, L.-H.; Shao, Q.-J.; Luo, W.; Ye, Z.-Y.; Li, Q.; Lin, S.-C. Detection of point mutations of the Axin1 gene in colorectal cancers. Int. J. Cancer 2003, 107, 696–699. [Google Scholar] [CrossRef]
- Yang, L.H.; Xu, H.T.; Li, Q.C.; Jiang, G.Y.; Zhang, X.P.; Zhao, H.Y.; Xu, K.; Wang, E.H. Abnormal hypermethylation and clinicopathological significance of Axin gene in lung cancer. Tumour Biol. 2013, 34, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. Axin and hepatocellular carcinomas. Nat. Genet. 2000, 24, 206–208. [Google Scholar] [CrossRef] [PubMed]
- Satoh, S.; Daigo, Y.; Furukawa, Y.; Kato, T.; Miwa, N.; Nishiwaki, T.; Kawasoe, T.; Ishiguro, H.; Fujita, M.; Tokino, T.; et al. AXIN1 mutations in hepatocellular carcinomas, and growth suppression in cancer cells by virus-mediated transfer of AXIN1. Nat. Genet. 2000, 24, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Abitbol, S.; Dahmani, R.; Coulouarn, C.; Ragazzon, B.; Mlecnik, B.; Senni, N.; Savall, M.; Bossard, P.; Sohier, P.; Drouet, V.; et al. AXIN deficiency in human and mouse hepatocytes induces hepatocellular carcinoma in the absence of β-catenin activation. J. Hepatol. 2018, 68, 1203–1213. [Google Scholar] [CrossRef] [PubMed]
- Feng, G.J.; Cotta, W.; Wei, X.Q.; Poetz, O.; Evans, R.; Jardé, T.; Reed, K.; Méniel, V.; Williams, G.T.; Clarke, A.R.; et al. Conditional Disruption of Axin1 Leads to Development of Liver Tumors in Mice. Gastroenterology 2012, 143, 1650–1659. [Google Scholar] [CrossRef] [PubMed]
- Neo, S.Y.; Zhang, Y.; Yaw, L.P.; Li, P.; Lin, S.-C. Axin-Induced Apoptosis Depends on the Extent of Its JNK Activation and Its Ability to Down-Regulate β-Catenin Levels. Biochem. Biophys. Res. Commun. 2000, 272, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.; Shakya, R.; Costantini, F. Impaired mammary gland and lymphoid development caused by inducible expression of Axin in transgenic mice. J. Cell Boil. 2001, 155, 1055–1064. [Google Scholar] [CrossRef]
- Zhang, Y.-L.; Guo, H.; Zhang, C.-S.; Lin, S.-Y.; Yin, Z.; Peng, Y.; Luo, H.; Shi, Y.; Lian, G.; Zhang, C.; et al. AMP as a Low-Energy Charge Signal Autonomously Initiates Assembly of AXIN-AMPK-LKB1 Complex for AMPK Activation. Cell Metab. 2013, 18, 546–555. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Youm, Y.-H.; Sun, Y.; Rim, J.-S.; Galbán, C.J.; Vandanmagsar, B.; Dixit, V.D. Axin expression in thymic stromal cells contributes to an age-related increase in thymic adiposity and is associated with reduced thymopoiesis independently of ghrelin signaling. J. Leukoc. Boil. 2009, 85, 928–938. [Google Scholar] [CrossRef] [Green Version]
- Willert, K.; Logan, C.Y.; Arora, A.; Fish, M.; Nusse, R. A Drosophila Axin homolog, Daxin, inhibits Wnt signaling. Development 1999, 126, 4165–4173. [Google Scholar]
- Song, Y.; Chung, S.; Kunes, S. Combgap Relays Wingless Signal Reception to the Determination of Cortical Cell Fate in the Drosophila Visual System. Mol. Cell 2000, 6, 1143–1154. [Google Scholar] [CrossRef]
- Gultekin, Y.; Steller, H. Axin proteolysis by Iduna is required for the regulation of stem cell proliferation and intestinal homeostasis in Drosophila. Development 2019, 146. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Posnien, N.; Bolognesi, R.; Fischer, T.D.; Rayl, P.; Oberhofer, G.; Kitzmann, P.; Brown, S.J.; Bucher, G. Asymmetrically expressed axin required for anterior development in Tribolium. Proc. Natl. Acad. Sci. USA 2012, 109, 7782–7786. [Google Scholar] [CrossRef] [PubMed]
- Foulquier, S.; Daskalopoulos, E.P.; Lluri, G.; Hermans, K.C.M.; Deb, A.; Blankesteijn, W.M. WNT Signaling in Cardiac and Vascular Disease. Pharmacol. Rev. 2018, 70, 68–141. [Google Scholar] [CrossRef] [PubMed]
- Gleason, J.E.; Eisenmann, D.M. Wnt signaling controls the stem cell-like asymmetric division of the epithelial seam cells during C. elegans larval development. Dev. Boil. 2010, 348, 58–66. [Google Scholar] [CrossRef]
- Mallick, A.; Ranawade, A.; Gupta, B.P. Role of PRY-1/Axin in heterochronic miRNA-mediated seam cell development. BMC Dev. Boil. 2019, 19, 17. [Google Scholar] [CrossRef]
- Zhang, Y.; Neo, S.Y.; Wang, X.; Han, J.; Lin, S.-C. Axin Forms a Complex with MEKK1 and Activates c-Jun NH2-terminal Kinase/Stress-activated Protein Kinase through Domains Distinct from Wnt Signaling. J. Boil. Chem. 1999, 274, 35247–35254. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Ng, W.W.; Jin, L.-H.; Ye, Z.; Han, J.; Lin, S.-C. Axin Utilizes Distinct Regions for Competitive MEKK1 and MEKK4 Binding and JNK Activation. J. Boil. Chem. 2003, 278, 37451–37458. [Google Scholar] [CrossRef] [Green Version]
- Rui, Y.; Xu, Z.; Xiong, B.; Cao, Y.; Lin, S.; Zhang, M.; Chan, S.-C.; Luo, W.; Han, Y.; Lu, Z.; et al. A β-Catenin-Independent Dorsalization Pathway Activated by Axin/JNK Signaling and Antagonized by Aida. Dev. Cell 2007, 13, 268–282. [Google Scholar] [CrossRef]
- Luo, K. Signaling Cross Talk between TGF-β/Smad and Other Signaling Pathways. Cold Spring Harb. Perspect. Boil. 2017, 9, a022137. [Google Scholar] [CrossRef]
- Guo, X.; Ramirez, A.; Waddell, D.S.; Li, Z.; Liu, X.; Wang, X.-F. Axin and GSK3-β control Smad3 protein stability and modulate TGF-β signaling. Genome Res. 2008, 22, 106–120. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Rui, H.; Wang, J.; Lin, S.; He, Y.; Chen, M.; Li, Q.; Ye, Z.; Zhang, S.; Chan, S.C.; et al. Axin is a scaffold protein in TGF-β signaling that promotes degradation of Smad7 by Arkadia. EMBO J. 2006, 25, 1646–1658. [Google Scholar] [CrossRef] [PubMed]
- Rui, Y.; Xu, Z.; Lin, S.; Li, Q.; Rui, H.; Luo, W.; Zhou, H.-M.; Cheung, P.-Y.; Wu, Z.; Ye, Z.; et al. Axin stimulates p53 functions by activation of HIPK2 kinase through multimeric complex formation. EMBO J. 2004, 23, 4583–4594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Lin, S.; Wang, X.; Lian, G.; Lu, Z.; Guo, H.; Ruan, K.; Wang, Y.; Ye, Z.; Han, J.; et al. Axin determines cell fate by controlling the p53 activation threshold after DNA damage. Nature 2009, 11, 1128–1134. [Google Scholar]
- Jung, Y.-S.; Qian, Y.; Chen, X. Pirh2 RING-finger E3 ubiquitin ligase: Its role in tumorigenesis and cancer therapy. FEBS Lett. 2012, 586, 1397–1402. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wang, X.; Wu, X.; Rui, Y.; Liu, W.; Wang, J.; Wang, X.; Liou, Y.-C.; Ye, Z.; Lin, S.-C. Daxx Cooperates with the Axin/HIPK2/p53 Complex to Induce Cell Death. Cancer Res. 2007, 67, 66–74. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, H.; Nusse, R. Identification of DCAP, a drosophila homolog of a glucose transport regulatory complex. Mech. Dev. 2002, 119, 115–119. [Google Scholar] [CrossRef]
- Yamazaki, H.; Yanagawa, S.-I. Axin and the Axin/Arrow-binding protein DCAP mediate glucose–glycogen metabolism. Biochem. Biophys. Res. Commun. 2003, 304, 229–235. [Google Scholar] [CrossRef]
- Guo, H.-L.; Zhang, C.; Liu, Q.; Li, Q.; Lian, G.; Wu, D.; Li, X.; Zhang, W.; Shen, Y.; Ye, Z.; et al. The Axin/TNKS complex interacts with KIF3A and is required for insulin-stimulated GLUT4 translocation. Cell Res. 2012, 22, 1246–1257. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.Y.; Mo, C.L.; Cai, Y.H.; Wang, W.J.; Hong, X.X.; Zhang, K.K.; Liu, Q.F.; Liu, Y.J.; Hong, J.J.; He, T.; et al. Pygo2 Regulates Adiposity and Glucose Homeostasis via beta-Catenin-Axin2-GSK3beta Signaling Pathway. Diabetes 2018, 67, 2569–2584. [Google Scholar] [CrossRef]
- Gho, M.; Schweisguth, F. Frizzled signalling controls orientation of asymmetric sense organ precursor cell divisions in Drosophila. Nature 1998, 393, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Cadigan, K.M.; Nusse, R. Wnt signaling: A common theme in animal development. Genes Dev. 1997, 11, 3286–3305. [Google Scholar] [CrossRef] [PubMed]
- Cowan, C.A.; Henkemeyer, M. The SH2/SH3 adaptor Grb4 transduces B-ephrin reverse signals. Nature 2001, 413, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. Wnt/β-Catenin Signaling in Development and Disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef]
- Gorrepati, L.; Krause, M.W.; Chen, W.; Brodigan, T.M.; Correa-Mendez, M.; Eisenmann, D.M. Identification of Wnt Pathway Target Genes Regulating the Division and Differentiation of Larval Seam Cells and Vulval Precursor Cells in Caenorhabditis elegans. G3 Genes Genomes Genet. 2015, 5, 1551–1566. [Google Scholar] [CrossRef]
- Ranawade, A.; Mallick, A.; Gupta, B.P. PRY-1/Axin signaling regulates lipid metabolism in Caenorhabditis elegans. PLoS ONE 2018, 13, e0206540. [Google Scholar] [CrossRef]
- Choi, S.-H.; Choi, K.-M.; Ahn, H.-J. Coexpression and protein-protein complexing of DIX domains of human Dvl1 and Axin1 protein. BMB Rep. 2010, 43, 609–613. [Google Scholar] [CrossRef]
- Fiedler, M.; Mendoza-Topaz, C.; Rutherford, T.J.; Mieszczanek, J.; Bienz, M. Dishevelled interacts with the DIX domain polymerization interface of Axin to interfere with its function in down-regulating β-catenin. Proc. Natl. Acad. Sci. USA 2011, 108, 1937–1942. [Google Scholar] [CrossRef]
- Sakanaka, C.; Williams, L.T. Functional domains of axin. Importance of the C terminus as an oligomerization domain. J. Boil. Chem. 1999, 274, 14090–14093. [Google Scholar] [CrossRef]
- Yokoyama, N.; Markova, N.G.; Wang, H.-Y.; Malbon, C.C. Assembly of Dishevelled 3-based supermolecular complexes via phosphorylation and Axin. J. Mol. Signal. 2012, 7, 8. [Google Scholar] [CrossRef]
- Schwarz-Romond, T.; Fiedler, M.; Shibata, N.; Butler, P.J.G.; Kikuchi, A.; Higuchi, Y.; Bienz, M. The DIX domain of Dishevelled confers Wnt signaling by dynamic polymerization. Nat. Struct. Mol. Boil. 2007, 14, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Schwarz-Romond, T.; Metcalfe, C.; Bienz, M. Dynamic recruitment of axin by Dishevelled protein assemblies. J. Cell Sci. 2007, 120, 2402–2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; Wang, S.; Li, L. New insights into the regulation of Axin function in canonical Wnt signaling pathway. Protein Cell 2014, 5, 186–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, C.; Xiao, G.; Hu, J. Regulation of Wnt/β-catenin signaling by posttranslational modifications. Cell Biosci. 2014, 4, 13. [Google Scholar] [CrossRef]
- Jho, E.-H.; Lomvardas, S.; Costantini, F. A GSK3β Phosphorylation Site in Axin Modulates Interaction with β-Catenin and Tcf-Mediated Gene Expression. Biochem. Biophys. Res. Commun. 1999, 266, 28–35. [Google Scholar] [CrossRef]
- Gao, Z.-H.; Seeling, J.M.; Hill, V.; Yochum, A.; Virshup, D.M. Casein kinase I phosphorylates and destabilizes the β-catenin degradation complex. Proc. Natl. Acad. Sci. USA 2002, 99, 1182–1187. [Google Scholar] [CrossRef]
- Willert, K.; Shibamoto, S.; Nusse, R. Wnt-induced dephosphorylation of Axin releases beta -catenin from the Axin complex. Genes Dev. 1999, 13, 1768–1773. [Google Scholar] [CrossRef] [Green Version]
- Strovel, E.T.; Wu, D.; Sussman, D.J. Protein Phosphatase 2C Dephosphorylates Axin and Activates LEF-1-dependent Transcription. J. Boil. Chem. 2000, 275, 2399–2403. [Google Scholar] [CrossRef]
- Tacchelly-Benites, O.; Wang, Z.; Yang, E.; Benchabane, H.; Tian, A.; Randall, M.P.; Ahmed, Y. Axin phosphorylation in both Wnt-off and Wnt-on states requires the tumor suppressor APC. PLoS Genet. 2018, 14, e1007178. [Google Scholar] [CrossRef]
- Kim, S.-E.; Huang, H.; Zhao, M.; Zhang, X.; Zhang, A.; Semonov, M.V.; Macdonald, B.T.; Zhang, X.; Abreu, J.G.; Peng, L.; et al. Wnt stabilization of β-catenin reveals principles for morphogen receptor-scaffold assemblies. Science 2013, 340, 867–870. [Google Scholar] [CrossRef]
- Huang, S.-M.A.; Mishina, Y.M.; Liu, S.; Cheung, A.; Stegmeier, F.; Michaud, G.A.; Charlat, O.; Wiellette, E.; Zhang, Y.; Wiessner, S.; et al. Tankyrase inhibition stabilizes axin and antagonizes Wnt signalling. Nature 2009, 461, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, S.; Mickanin, C.; Feng, Y.; Charlat, O.; Michaud, G.A.; Schirle, M.; Shi, X.; Hild, M.; Bauer, A.; et al. RNF146 is a poly(ADP-ribose)-directed E3 ligase that regulates axin degradation and Wnt signalling. Nature 2011, 13, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Callow, M.G.; Tran, H.; Phu, L.; Lau, T.; Lee, J.; Sandoval, W.N.; Liu, P.S.; Bheddah, S.; Tao, J.; Lill, J.R.; et al. Ubiquitin Ligase RNF146 Regulates Tankyrase and Axin to Promote Wnt Signaling. PLoS ONE 2011, 6, e22595. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Tacchelly-Benites, O.; Yang, E.; Ahmed, Y. Dual Roles for Membrane Association of Drosophila Axin in Wnt Signaling. PLoS Genet. 2016, 12, e1006494. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Li, X.; Ray, L.; Song, H.; Qu, J.; Lin, S.; Lin, X. The Drosophila tankyrase regulates Wg signaling depending on the concentration of Daxin. Cell Signal. 2014, 26, 1717–1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, L.; Jiang, B.; Jiang, X.; Charlat, O.; Chen, A.; Mickanin, C.; Bauer, A.; Xu, W.; Yan, X.; Cong, F. The SIAH E3 ubiquitin ligases promote Wnt/β-catenin signaling through mediating Wnt-induced Axin degradation. Genes Dev. 2017, 31, 904–915. [Google Scholar] [CrossRef]
- Wang, Z.; Tacchelly-Benites, O.; Noble, G.P.; Johnson, M.K.; Gagne, J.P.; Poirier, G.G.; Ahmed, Y. A Context-Dependent Role for the RNF146 Ubiquitin Ligase in Wingless/Wnt Signaling in Drosophila. Genetics 2019, 211, 913–923. [Google Scholar] [CrossRef]
- Fei, C.; Li, Z.; Li, C.; Chen, Y.; Chen, Z.; He, X.; Mao, L.; Wang, X.; Zeng, R.; Li, L. Smurf1-Mediated Lys29-Linked Nonproteolytic Polyubiquitination of Axin Negatively Regulates Wnt/β-Catenin Signaling. Mol. Cell. Boil. 2013, 33, 4095–4105. [Google Scholar] [CrossRef]
- Kim, S.; Jho, E.-H. The Protein Stability of Axin, a Negative Regulator of Wnt Signaling, Is Regulated by Smad Ubiquitination Regulatory Factor 2 (Smurf2). J. Boil. Chem. 2010, 285, 36420–36426. [Google Scholar] [CrossRef] [Green Version]
- Kadoya, T.; Kishida, S.; Fukui, A.; Hinoi, T.; Michiue, T.; Asashima, M.; Kikuchi, A. Inhibition of Wnt Signaling Pathway by a Novel Axin-binding Protein. J. Boil. Chem. 2000, 275, 37030–37037. [Google Scholar] [CrossRef] [Green Version]
- Kadoya, T.; Yamamoto, H.; Suzuki, T.; Yukita, A.; Fukui, A.; Michiue, T.; Asahara, T.; Tanaka, K.; Asashima, M.; Kikuchi, A. Desumoylation Activity of Axam, a Novel Axin-Binding Protein, Is Involved in Downregulation of β-Catenin. Mol. Cell. Boil. 2002, 22, 3803–3819. [Google Scholar] [CrossRef]
- Rui, H.-L.; Fan, E.; Zhou, H.-M.; Xu, Z.; Zhang, Y.; Lin, S.-C. SUMO-1 Modification of the C-terminal KVEKVD of Axin Is Required for JNK Activation but Has No Effect on Wnt Signaling. J. Boil. Chem. 2002, 277, 42981–42986. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.J.; Chia, I.V.; Costantini, F. SUMOylation target sites at the C terminus protect Axin from ubiquitination and confer protein stability. FASEB J. 2008, 22, 3785–3794. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Li, M.; Ding, Y.; Zhang, L.-S.; Xi, Y.; Pan, W.-J.; Tao, D.-L.; Wang, J.-Y.; Li, L. Identification of Zinc-finger BED Domain-containing 3 (Zbed3) as a Novel Axin-interacting Protein That Activates Wnt/β-Catenin Signaling. J. Boil. Chem. 2009, 284, 6683–6689. [Google Scholar] [CrossRef]
- Wang, S.; Yin, J.; Chen, D.; Nie, F.; Song, X.; Fei, C.; Miao, H.; Jing, C.; Ma, W.; Wang, L.; et al. Small-molecule modulation of Wnt signaling via modulating the Axin-LRP5/6 interaction. Nat. Methods 2013, 9, 579–585. [Google Scholar] [CrossRef]
- Silver, S.J.; Hagen, J.W.; Okamura, K.; Perrimon, N.; Lai, E.C. Functional screening identifies miR-315 as a potent activator of Wingless signaling. Proc. Natl. Acad. Sci. USA 2007, 104, 18151–18156. [Google Scholar] [CrossRef] [Green Version]
- Egea, V.; Zahler, S.; Rieth, N.; Neth, P.; Popp, T.; Kehe, K.; Jochum, M.; Ries, C. Tissue inhibitor of metalloproteinase-1 (TIMP-1) regulates mesenchymal stem cells through let-7f microRNA and Wnt/β-catenin signaling. Proc. Natl. Acad. Sci. USA 2012, 109, E309–E316. [Google Scholar] [CrossRef]
- Lee, I.; Ajay, S.S.; Yook, J.I.; Kim, H.S.; Hong, S.H.; Kim, N.H.; Dhanasekaran, S.M.; Chinnaiyan, A.M.; Athey, B.D. New class of microRNA targets containing simultaneous 5′-UTR and 3′-UTR interaction sites. Genome Res. 2009, 19, 1175–1183. [Google Scholar] [CrossRef]
- Kim, J.S.; Park, S.Y.; Lee, S.A.; Park, M.G.; Yu, S.K.; Lee, M.H.; Park, M.R.; Kim, S.G.; Oh, J.S.; Lee, S.Y.; et al. MicroRNA-205 suppresses the oral carcinoma oncogenic activity via down-regulation of Axin-2 in KB human oral cancer cell. Mol. Cell. Biochem. 2014, 387, 71–79. [Google Scholar] [CrossRef]
- Korswagen, H.C.; Coudreuse, D.Y.; Betist, M.C.; Van De Water, S.; Zivkovic, D.; Clevers, H.C. The Axin-like protein PRY-1 is a negative regulator of a canonical Wnt pathway in C. elegans. Genome Res. 2002, 16, 1291–1302. [Google Scholar] [CrossRef]
- Sawa, H.; Korswagen, H.C. Wnt signaling in C. elegans. WormBook 2013. [Google Scholar] [CrossRef]
- Seetharaman, A.; Cumbo, P.; Bojanala, N.; Gupta, B.P. Conserved mechanism of Wnt signaling function in the specification of vulval precursor fates in C. elegans and C. briggsae. Dev. Biol. 2010, 346, 128–139. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Ou, Y.; Li, Y.; Hu, S.; Shao, L.-W.; Liu, Y. Metformin extends C. elegans lifespan through lysosomal pathway. eLife 2017, 6. [Google Scholar] [CrossRef]
- Larkin, M.; Blackshields, G.; Brown, N.; Chenna, R.; Mcgettigan, P.; Mc William, H.; Valentin, F.; Wallace, I.; Wilm, A.; López, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Stothard, P. The Sequence Manipulation Suite: JavaScript Programs for Analyzing and Formatting Protein and DNA Sequences. Biotechniques 2000, 28, 1102–1104. [Google Scholar] [CrossRef] [Green Version]
- Rella, L.; Póvoa, E.E.F.; Korswagen, H.C. The Caenorhabditis elegans Q neuroblasts: A powerful system to study cell migration at single-cell resolutionin vivo. Genesis 2016, 54, 198–211. [Google Scholar] [CrossRef]
- Schmitz, C.; Kinge, P.; Hutter, H. Axon guidance genes identified in a large-scale RNAi screen using the RNAi-hypersensitive Caenorhabditis elegans strain nre-1(hd20) lin-15b(hd126). Proc. Natl. Acad. Sci. USA 2007, 104, 834–839. [Google Scholar] [CrossRef]
- Schneider, J.; Skelton, R.L.; Von Stetina, S.E.; Middelkoop, T.C.; van Oudenaarden, A.; Korswagen, H.C.; Miller, D.M., 3rd. UNC-4 antagonizes Wnt signaling to regulate synaptic choice in the C. elegans motor circuit. Development 2012, 139, 2234–2245. [Google Scholar] [CrossRef]
- Inestrosa, N.C.; Toledo, E.M. The role of Wnt signaling in neuronal dysfunction in Alzheimer’s Disease. Mol. Neurodegener. 2008, 3, 9. [Google Scholar] [CrossRef]
- Berwick, D.C.; Harvey, K. The importance of Wnt signalling for neurodegeneration in Parkinson’s disease. Biochem. Soc. Trans. 2012, 40, 1123–1128. [Google Scholar] [CrossRef]
- Richman, C.; Rashid, S.; Prashar, S.; Mishra, R.; Selvaganapathy, P.R.; Gupta, B.P. C. elegans MANF Homolog Is Necessary for the Protection of Dopaminergic Neurons and ER Unfolded Protein Response. Front. Mol. Neurosci. 2018, 12, 544. [Google Scholar] [CrossRef] [Green Version]
- Rocheleau, C.E.; Downs, W.D.; Lin, R.; Wittmann, C.; Bei, Y.; Cha, Y.-H.; Ali, M.; Priess, J.R.; Mello, C.C. Wnt Signaling and an APC-Related Gene Specify Endoderm in Early C. elegans Embryos. Cell 1997, 90, 707–716. [Google Scholar] [CrossRef] [Green Version]
- Thorpe, C.J.; Schlesinger, A.; Carter, J.; Bowerman, B. Wnt Signaling Polarizes an Early C. elegans Blastomere to Distinguish Endoderm from Mesoderm. Cell 1997, 90, 695–705. [Google Scholar] [CrossRef] [Green Version]
- Mizumoto, K.; Sawa, H. Cortical β-Catenin and APC Regulate Asymmetric Nuclear β-Catenin Localization during Asymmetric Cell Division in C. elegans. Dev. Cell 2007, 12, 287–299. [Google Scholar] [CrossRef]
- Walston, T.; Tuskey, C.; Edgar, L.; Hawkins, N.; Ellis, G.; Bowerman, B.; Wood, W.; Hardin, J. Multiple Wnt Signaling Pathways Converge to Orient the Mitotic Spindle in Early C. elegans Embryos. Dev. Cell 2004, 7, 831–841. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Kim, S.; Ishidate, T.; Bei, Y.; Pang, K.; Shirayama, M.; Trzepacz, C.; Brownell, D.R.; Mello, C.C. Wnt signaling drives WRM-1/beta-catenin asymmetries in early C. elegans embryos. Genes Dev. 2005, 19, 1749–1754. [Google Scholar] [CrossRef]
- Takeshita, H.; Sawa, H. Asymmetric cortical and nuclear localizations of WRM-1/beta-catenin during asymmetric cell division in C. elegans. Genes Dev. 2005, 19, 1743–1748. [Google Scholar] [CrossRef]
- Huang, S.; Shetty, P.; Robertson, S.M.; Lin, R. Binary cell fate specification during C. elegans embryogenesis driven by reiterated reciprocal asymmetry of TCF POP-1 and its coactivator {beta}-catenin SYS-1. Development 2007, 134, 2685–2695. [Google Scholar] [CrossRef]
- Lo, M.-C.; Gay, F.; Odom, R.; Shi, Y.; Lin, R. Phosphorylation by the beta-catenin/MAPK complex promotes 14-3-3-mediated nuclear export of TCF/POP-1 in signal-responsive cells in C. elegans. Cell 2004, 117, 95–106. [Google Scholar] [CrossRef]
- Phillips, B.T.; Kidd, A.R., 3rd; King, R.; Hardin, J.; Kimble, J. Reciprocal asymmetry of SYS-1/beta-catenin and POP-1/TCF controls asymmetric divisions in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2007, 104, 3231–3236. [Google Scholar] [CrossRef]
- Lin, R.; Hill, R.J.; Priess, J.R. POP-1 and Anterior–Posterior Fate Decisions in C. elegans Embryos. Cell 1998, 92, 229–239. [Google Scholar] [CrossRef]
- Lin, R.; Thompson, S.; Priess, J.R. pop-1 Encodes an HMG box protein required for the specification of a mesoderm precursor in Early C. elegans embryos. Cell 1995, 83, 599–609. [Google Scholar] [CrossRef] [Green Version]
- Page, A.P.; Johnstone, I.L. The cuticle. WormBook 2007, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulston, J.; Horvitz, H. Post-embryonic cell lineages of the nematode, Caenorhabditis elegans. Dev. Boil. 1977, 56, 110–156. [Google Scholar] [CrossRef]
- Park, F.D.; Tenlen, J.R.; Priess, J.R. C. elegans MOM-5/Frizzled Functions in MOM-2/Wnt-Independent Cell Polarity and Is Localized Asymmetrically prior to Cell Division. Curr. Boil. 2004, 14, 2252–2258. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, A.T.; Clemons, A.M.; Phillips, B.T. Unique and redundant β-catenin regulatory roles of two Dishevelled paralogs during C. elegans asymmetric cell division. J. Cell Sci. 2016, 129, 983–993. [Google Scholar] [CrossRef]
- Park, F.D.; Ma, L.; Lei, L.; Eng, S.R.; Turner, E.; Parada, L.F. Establishment of POP-1 asymmetry in early C. elegans embryos. Development 2003, 130, 3547–3556. [Google Scholar] [CrossRef]
- Maloof, J.N.; Whangbo, J.; Harris, J.M.; Jongeward, G.D.; Kenyon, C. A Wnt signaling pathway controls hox gene expression and neuroblast migration in C. elegans. Development 1999, 126, 37–49. [Google Scholar]
- Baldwin, A.T.; Phillips, B.T. The tumor suppressor APC differentially regulates multiple β-catenins through the function of axin and CKIα during C. elegans asymmetric stem cell divisions. J. Cell Sci. 2014, 127, 2771–2781. [Google Scholar] [CrossRef]
- Moss, E.G. Heterochronic Genes and the Nature of Developmental Time. Curr. Boil. 2007, 17, R425–R434. [Google Scholar] [CrossRef] [Green Version]
- Ambros, V. MicroRNAs and developmental timing. Curr. Opin. Genet. Dev. 2011, 21, 511–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salser, S.J.; Kenyon, C. A C. elegans Hox gene switches on, off, on and off again to regulate proliferation, differentiation and morphogenesis. Development 1996, 122, 1651–1661. [Google Scholar] [PubMed]
- Emmons, S.W. Male development. WormBook 2005, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sternberg, P.W. Vulval development. WormBook 2005. [Google Scholar] [CrossRef] [PubMed]
- Gleason, J.E.; Korswagen, H.C.; Eisenmann, D.M. Activation of Wnt signaling bypasses the requirement for RTK/Ras signaling during C. elegans vulval induction. Genome Res. 2002, 16, 1281–1290. [Google Scholar] [CrossRef] [PubMed]
- Eisenmann, D.M.; Kim, S.K. Protruding vulva mutants identify novel loci and Wnt signaling factors that function during Caenorhabditis elegans vulva development. Genetics 2000, 156, 1097–1116. [Google Scholar] [PubMed]
- Yu, H.; Seah, A.; Sternberg, P.W. Re-programming of C. elegans male epidermal precursor fates by Wnt, Hox, and LIN-12/Notch activities. Dev. Boil. 2010, 345, 1–11. [Google Scholar] [CrossRef]
- Wang, X.; Sommer, R.J. Antagonism of LIN-17/Frizzled and LIN-18/Ryk in nematode vulva induction reveals evolutionary alterations in core developmental pathways. PLoS Boil. 2011, 9, e1001110. [Google Scholar] [CrossRef]
- Watts, J.L.; Ristow, M. Lipid and Carbohydrate Metabolism in Caenorhabditis elegans. Genetics 2017, 207, 413–446. [Google Scholar]
- Watts, J.L.; Browse, J. A palmitoyl-CoA-specific delta9 fatty acid desaturase from Caenorhabditis elegans. Biochem. Biophys. Res. Commun. 2000, 272, 263–269. [Google Scholar] [CrossRef]
- Admasu, T.D.; Batchu, K.C.; Barardo, D.; Ng, L.F.; Lam, V.Y.M.; Xiao, L.; Cazenave-Gassiot, A.; Wenk, M.R.; Tolwinski, N.S.; Gruber, J. Drug Synergy Slows Aging and Improves Healthspan through IGF and SREBP Lipid Signaling. Dev. Cell 2018, 47, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Hou, N.S.; Taubert, S. Function and Regulation of Lipid Biology in Caenorhabditis elegans Aging. Front. Physiol. 2012, 3, 143. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mallick, A.; Taylor, S.K.B.; Ranawade, A.; Gupta, B.P. Axin Family of Scaffolding Proteins in Development: Lessons from C. elegans. J. Dev. Biol. 2019, 7, 20. https://doi.org/10.3390/jdb7040020
Mallick A, Taylor SKB, Ranawade A, Gupta BP. Axin Family of Scaffolding Proteins in Development: Lessons from C. elegans. Journal of Developmental Biology. 2019; 7(4):20. https://doi.org/10.3390/jdb7040020
Chicago/Turabian StyleMallick, Avijit, Shane K. B. Taylor, Ayush Ranawade, and Bhagwati P. Gupta. 2019. "Axin Family of Scaffolding Proteins in Development: Lessons from C. elegans" Journal of Developmental Biology 7, no. 4: 20. https://doi.org/10.3390/jdb7040020
APA StyleMallick, A., Taylor, S. K. B., Ranawade, A., & Gupta, B. P. (2019). Axin Family of Scaffolding Proteins in Development: Lessons from C. elegans. Journal of Developmental Biology, 7(4), 20. https://doi.org/10.3390/jdb7040020