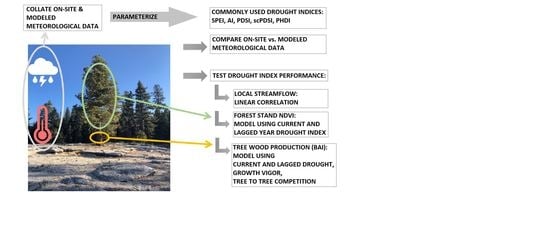
1. Introduction
Over the last two decades, droughts have been more extreme, extended, and hotter in the Pacific and Southwestern U.S. [
1,
2,
3,
4,
5], challenging our current understanding of drought and its effects on forests and humans alike. Massive tree die-offs in the Sierra Nevada of California [
6,
7] and surprise at its magnitude [
8,
9] have drawn world-wide attention. Linking the level, duration, and scale of drought to impacts on hydrological and biological resources is a critical step in anticipating future forest resource response and loss [
10].
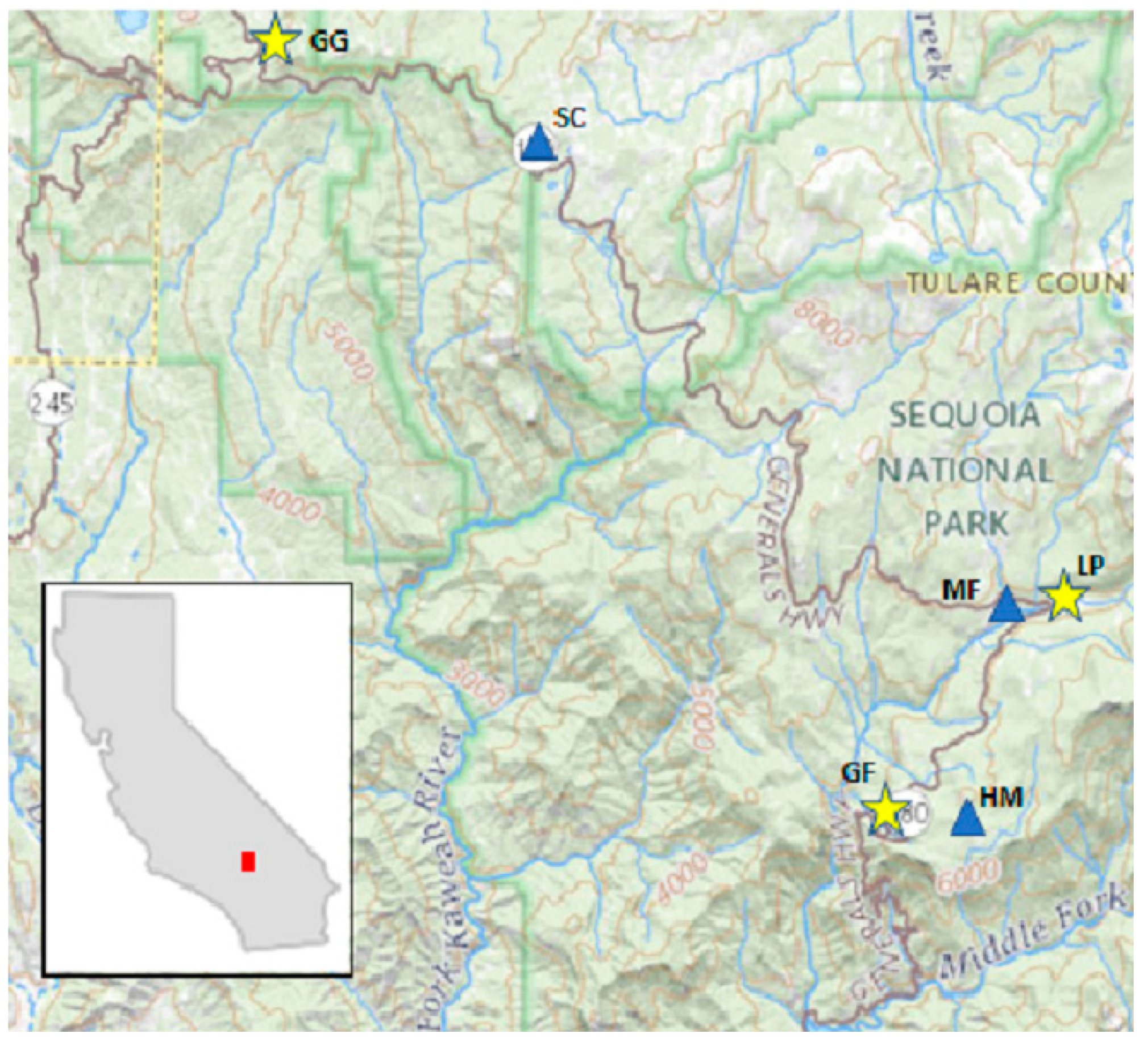
Water availability is a demonstrably vulnerable source of water for these forested ecosystems, which rely on, buffer, and supply water for human use. The Sierra Nevada provides ~60% of Californian water resources utilized by humans [
11]. Water resources are evaluated via drought indices (DIs), and these are used to assign the level of drought severity and help communicate and allocate near-term water resources for direct human consumption, agriculture, and industrial uses. Accurate estimates of water resources and their effects on biological responses are also needed (ecological drought) [
12]. Modeled meteorological data (PRISM, or Parameter-elevation Relationships on Independent Slopes Model) [
13,
14] are broadly used for a range of applications, and in particular to calculate DIs. The performance of PRISM in the foothills of the Appalachian Mountains was tested and the model performed well [
15]. On the eastern slope of the Sierra Nevada, PRISM temperature was greater than that measured on-site and there were seasonal differences in error [
16,
17]. Here, we tested point- (on-site) vs. pixel-based (4 km
2) meteorological parameterizations of commonly used DIs in complex, mid-elevation terrain (~2100 m) on the western slope of the Sierra Nevada. The indices chosen were SPEI (Standardized Precipitation and Evaporation Index) [
18], AI (Aridity Index) [
19,
20], PDSI and PHDI (Palmer Drought Severity Index, Palmer Hydrological Drought Index) [
21], and scPDSI (self-calibrating PDSI) [
22].
DIs rely on a combination of meteorological (precipitation and temperature), locational (slope, aspect, elevation, and latitude as a proxy for radiatively-driven evapotranspiration (ET)), and pedological measures (ET, soil runoff, soil moisture loss, and recharge). Although PDSI and derivatives provide default values for soil available water content (AWC), both this and depth to water table are unknown and possibly unknowable for much of the mid- to high-elevation forests on the western slope of the southern Sierra Nevada. In terms of vegetation in this ecosystem, Jeffrey pine (
Pinus jeffreyi Grev. & Balf.) rely on moisture and nutrients in patches of soil and or pockets of soil in bedrock interstices [
23,
24,
25] that are either large enough to support that tree or fed by ephemeral, surficial, or underground seeps.
DIs have been tested against hydrological [
26,
27] and biological metrics, including NDVI (Normalized Difference Vegetation Index) [
28,
29,
30,
31] and wood production [
32], but few studies tested more than one validation metric concurrently [
33,
34]. In this study on the western slope of the southern Sierra Nevada, we asked how closely aligned point- and pixel-based parameterizations were within a DI, and among the DIs, within the same parameterization using the strength of the correlation coefficients. Did the DIs with on-site or modeled parameterizations predict short term (7 yr) local streamflow, or stand NDVI (20 yr) equally well? How effective were DIs in predicting wood production (here basal area increment, BAI) of Jeffrey pine, as a decline in BAI can lead to tree mortality [
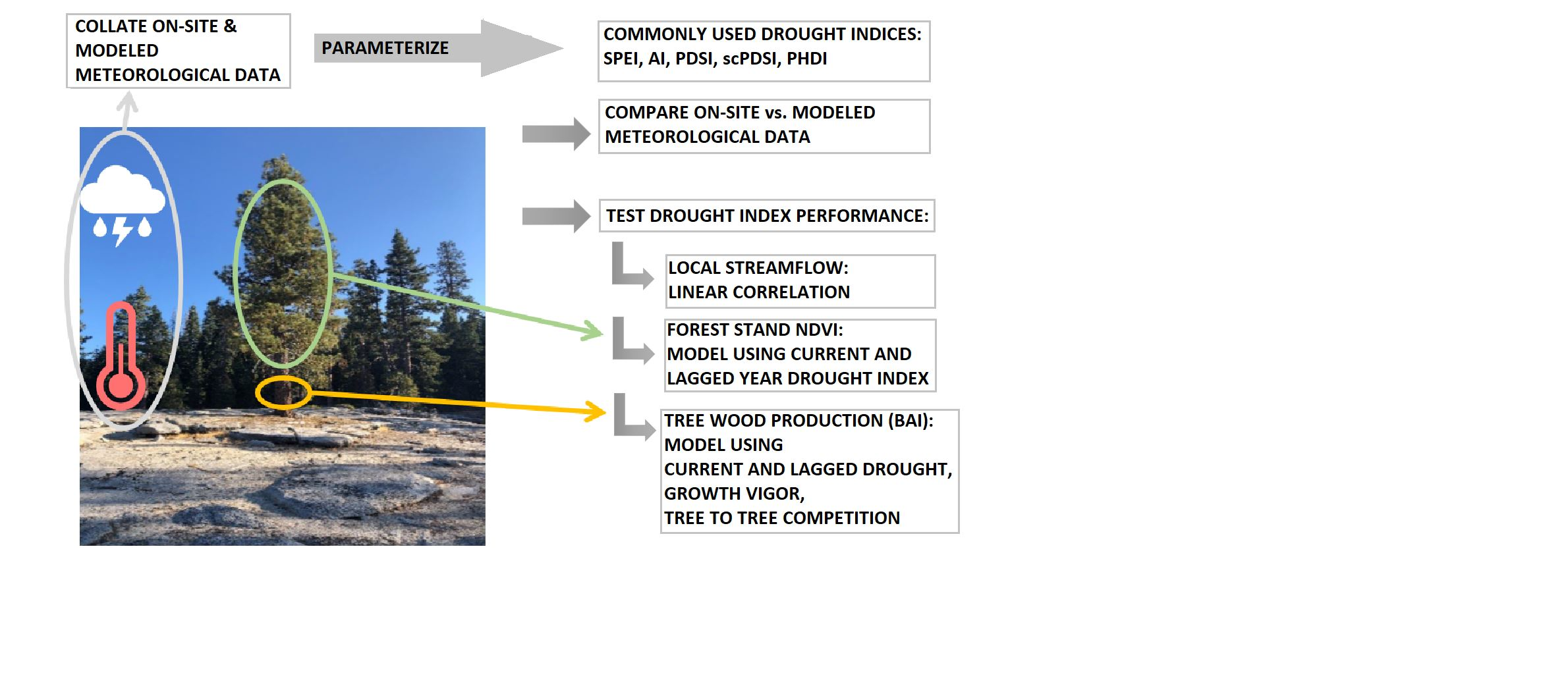
35]? To test the degree to which DIs could be used to anticipate tree decline in multi-year hydrologic droughts, we developed a model to predict BAI using current year and temporally lagged DIs as the primary drivers [
36,
37], with statistically selected biological covariates expected to modify (amplify or degrade) tree response to the environmental drivers, here crown vigor, tree bole growth rates at a mid-point in the analysis [
38,
39], and tree to tree competition of the target tree [
40].
4. Discussion
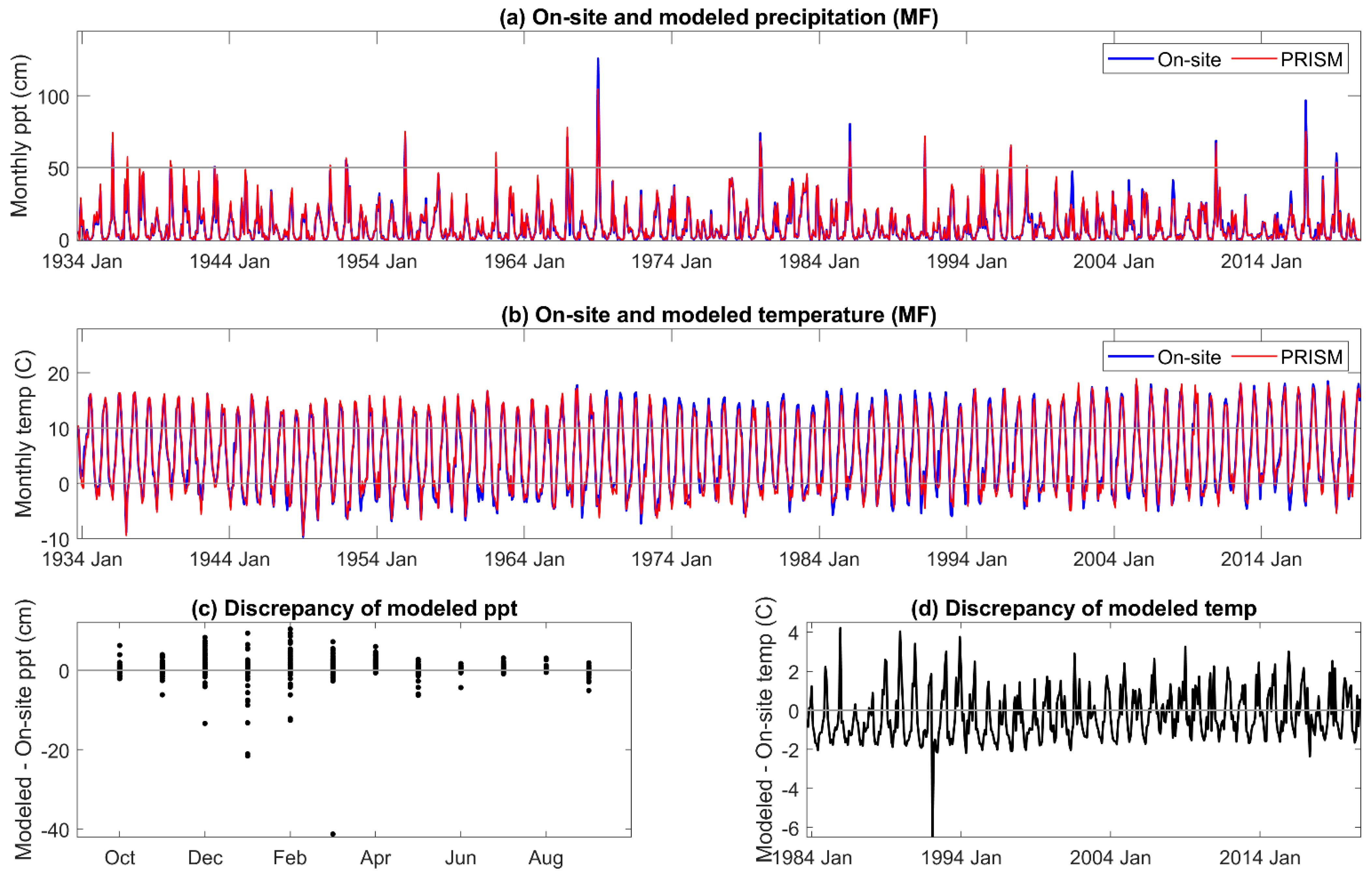
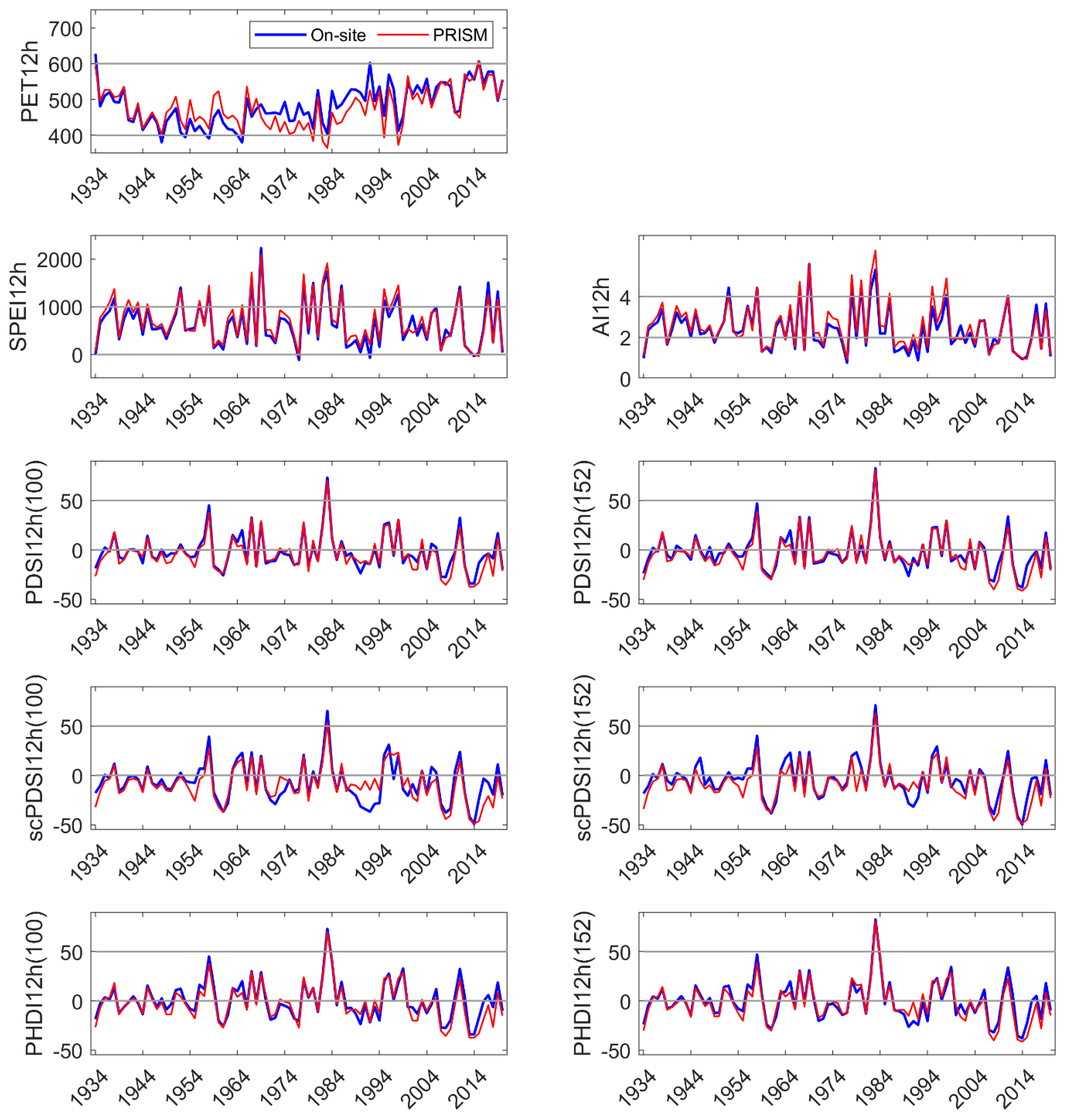
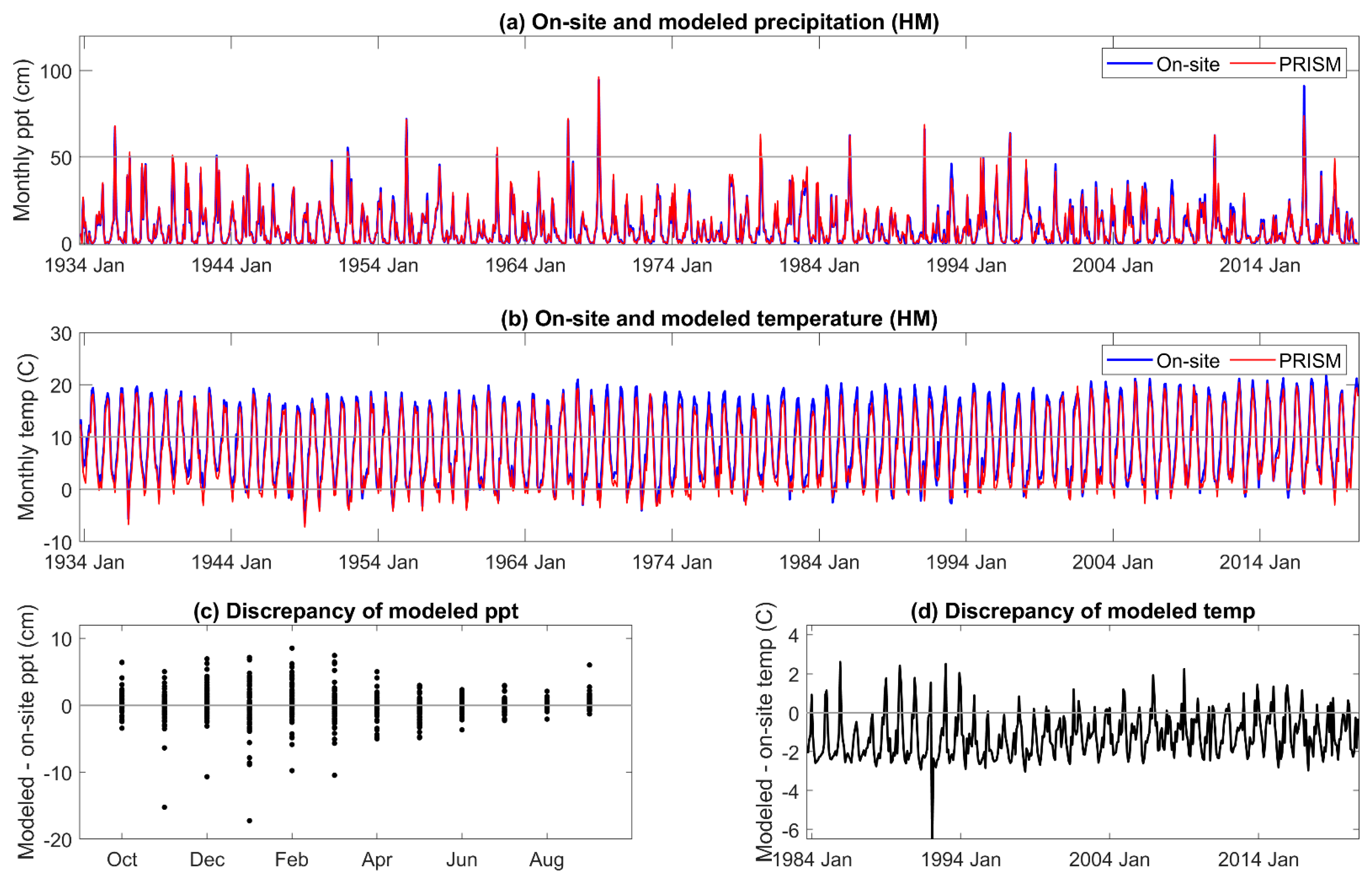
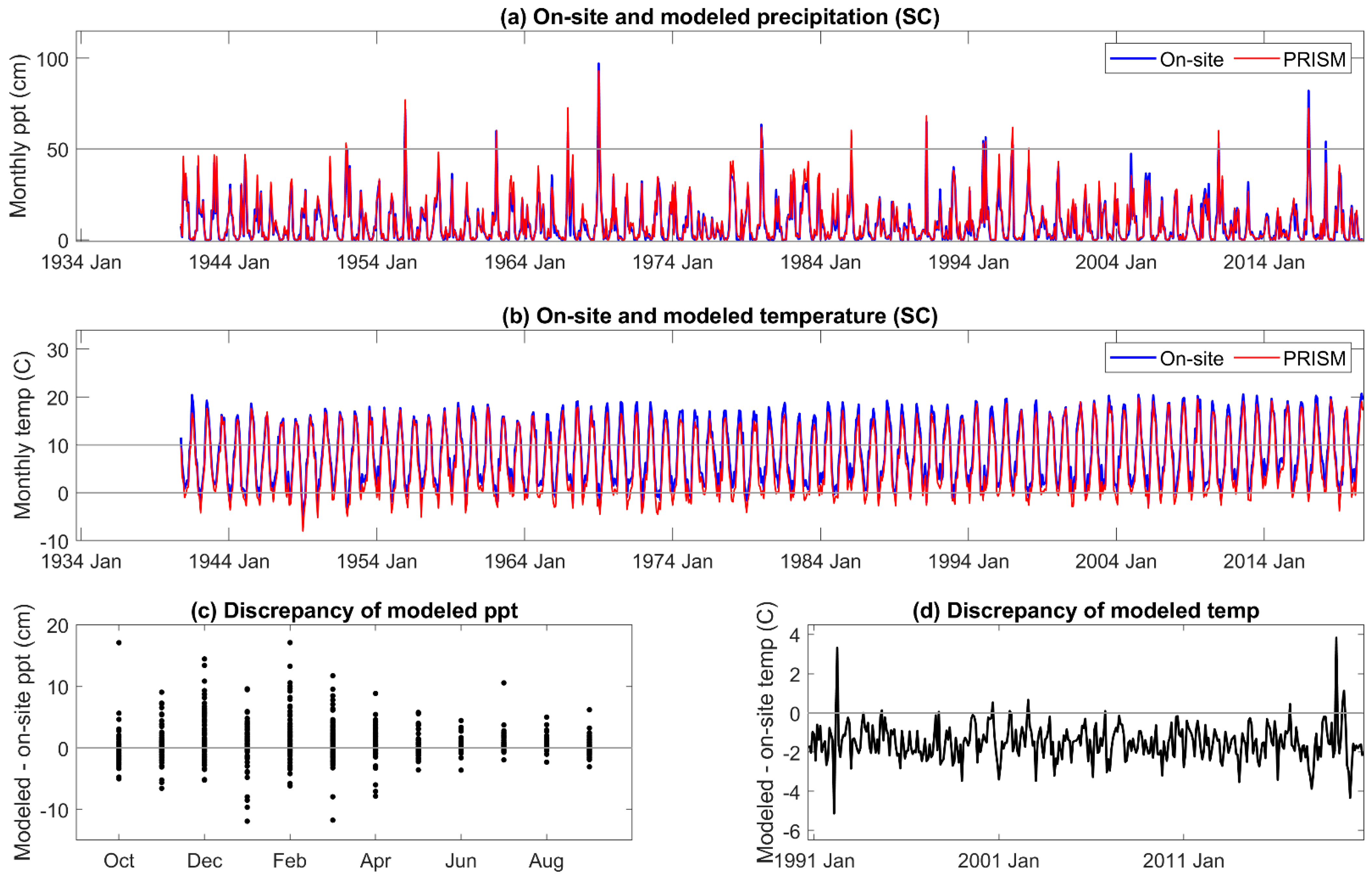
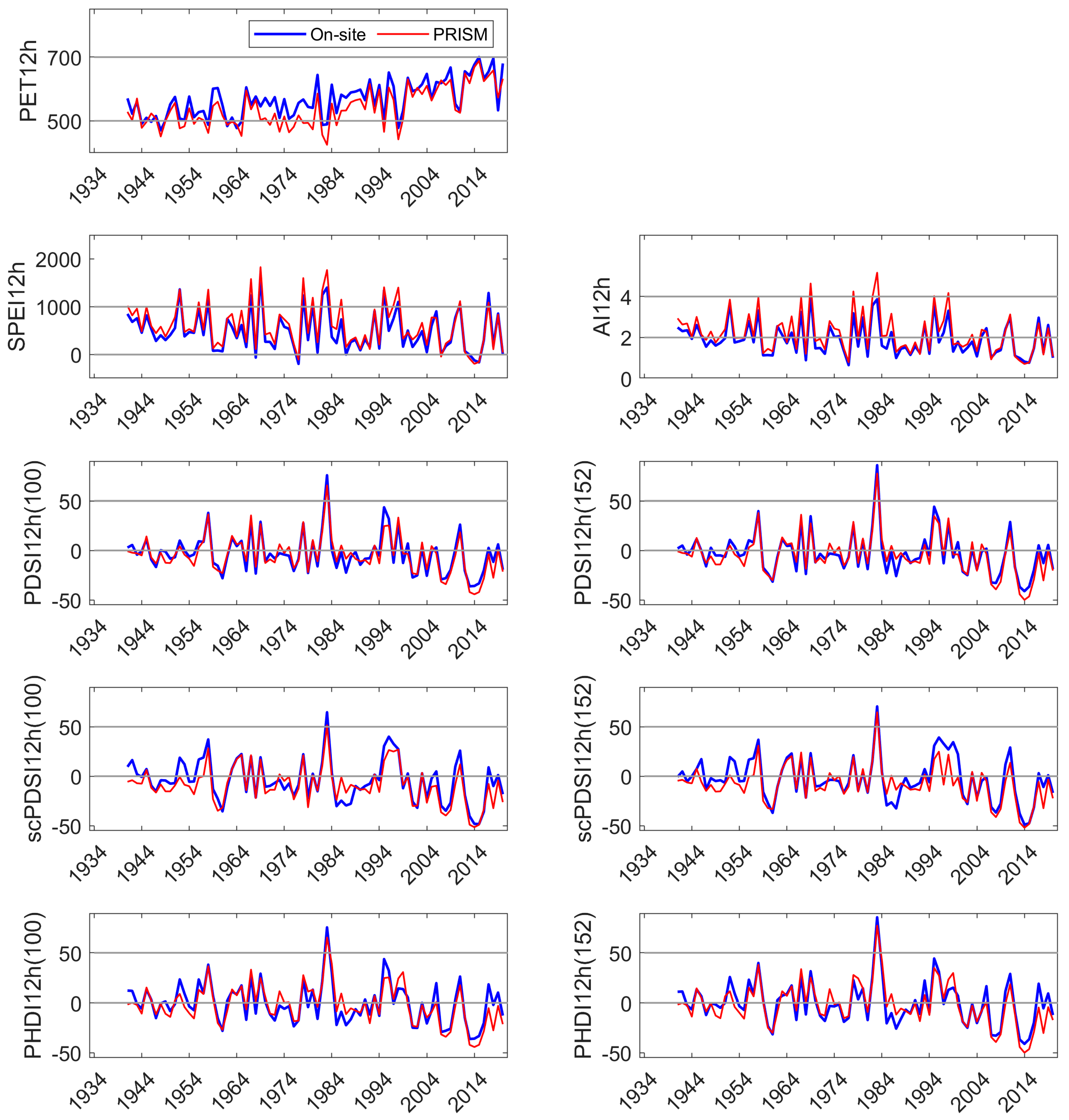
Two types of DIs were evaluated, one based on meteorological and physics-based inputs (SPEI and AI), and the other on meteorological-, physics-, and pedogenic-based inputs (PDSI and derivatives scPDSI and PHDI) in mid-elevation, complex terrain. Relatively few comparisons have been made between on-site and modeled meteorology in these landscapes. While on-site and PRISM monthly mean temperature and summed precipitation were highly correlated, periodic discrepancies in temperature were identified at our sites on the western slope of the Sierra Nevada. PRISM mean temperature was higher than that observed from November to January, and lower for April, May, and June. Higher temperatures in the winter increase ET but would have a low impact on vegetation. Lower temperatures in late spring would decrease calculated DIs and underestimate the level of drought severity when it occurs. In another study across a mid- to high elevation basin on the eastern slope of the Sierra Nevada [
16], PRISM maximum temperatures were lower relative to on-site values in early spring (March, April), and mid-summer (July, August). Of interest is that there was an overlap but not an identical match of underestimated monthly temperature between the western (this study) and eastern slopes [
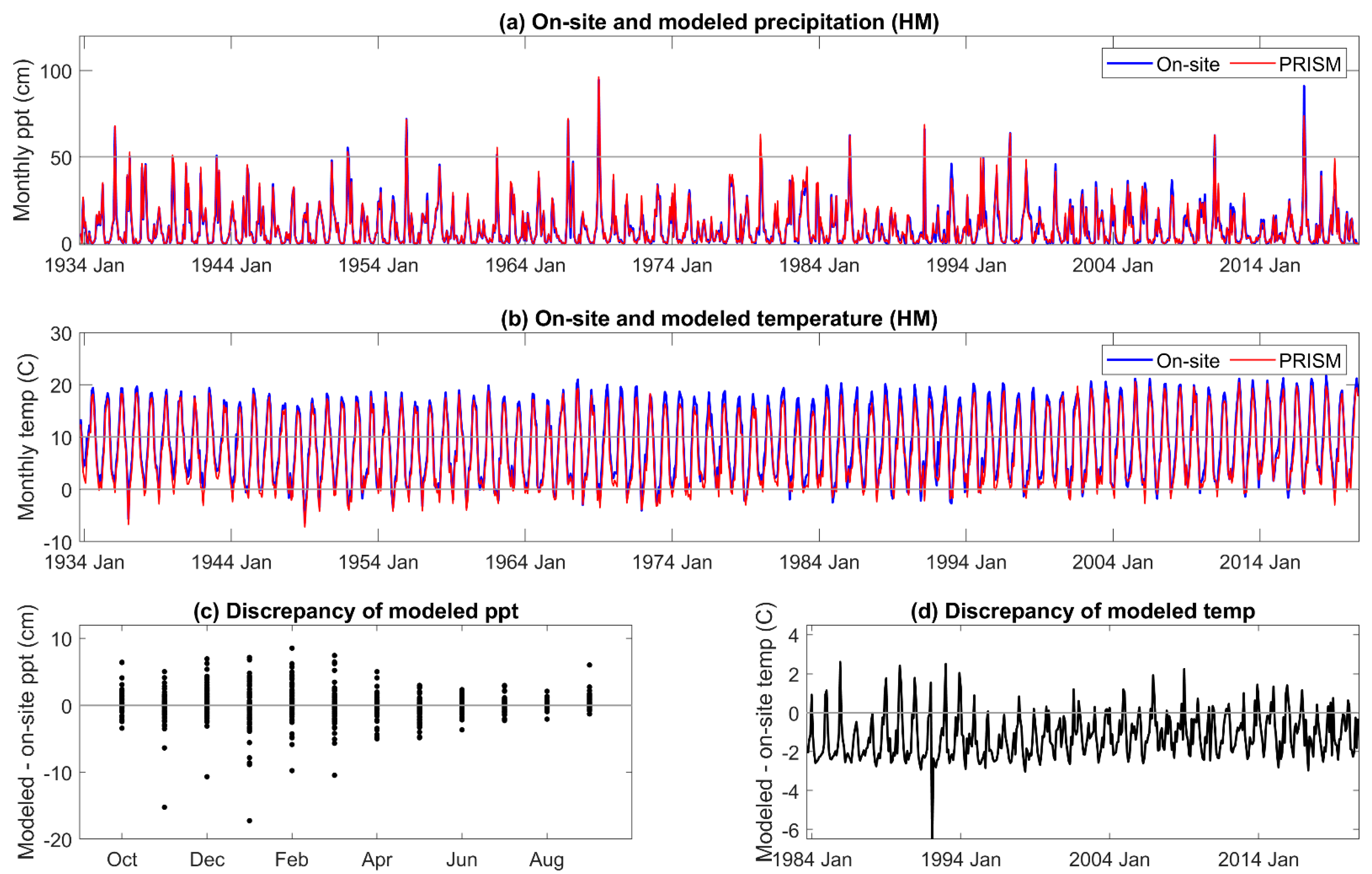
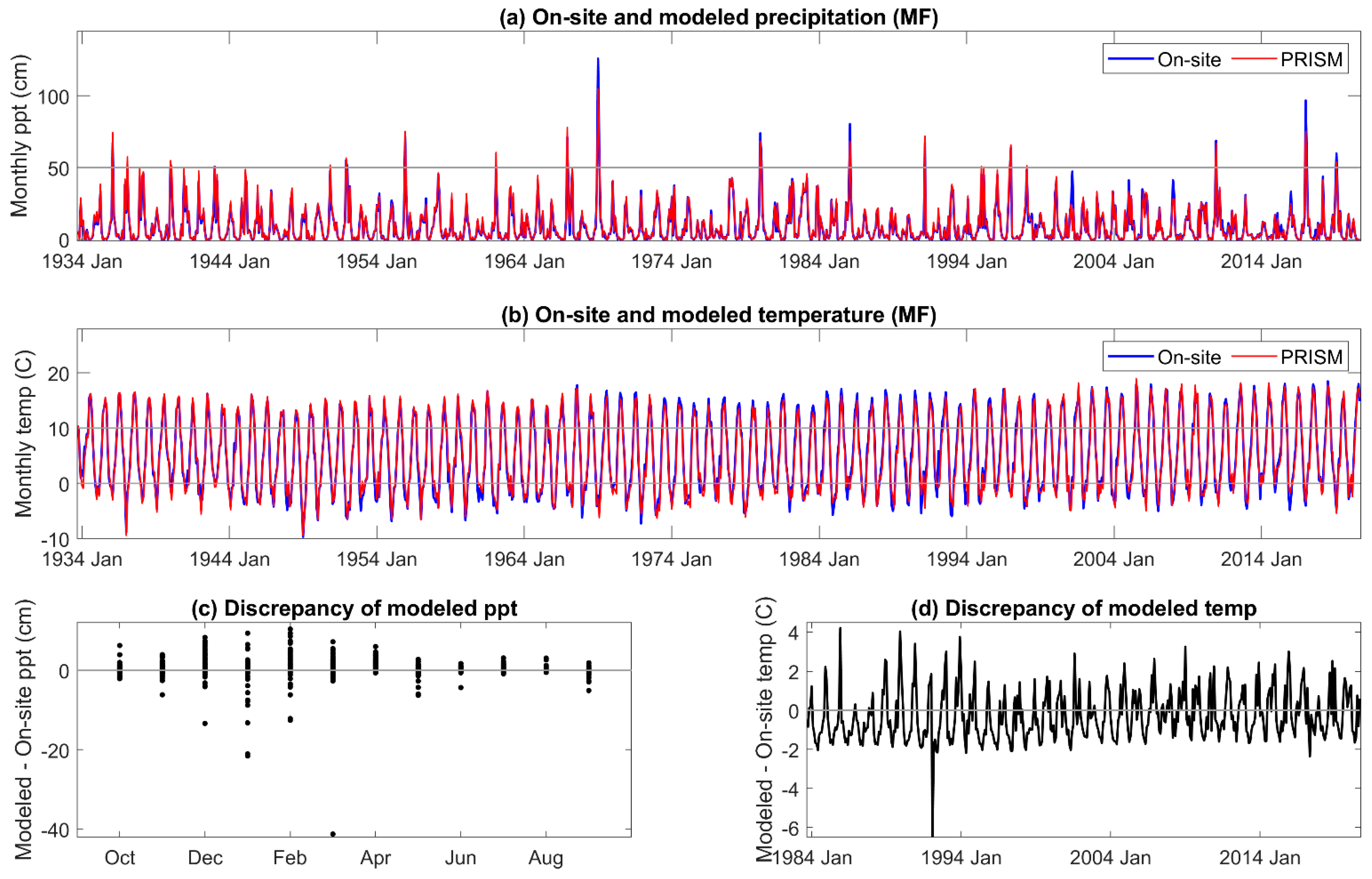
16] of the Sierra Nevada, and that the impact of the temperature discrepency would be greater in the latter study as water deficits increase through the summer. During the period of highest precipitation, from November to March, the estimated error variance of PRISM precipitation was high in our study. Extremes, both higher and lower than received, in on-site precipitation exceeded 5% of that estimated by PRISM. Overall, the annual precipitation estimated by PRISM was 3% to 8% greater than received across the three sites. The temperature and precipitation discrepancies of PRISM relative to on-site meteorology both contribute to an underestimation of DI severity during the growing season.
An extreme cold event was not predicted by PRISM (>5 °C discrepancy between modeled and on-site temperature) in December 1992 and January 1993 [
67]. Extremely low temperatures on the western slope of the Sierra Nevada are rare but can occur with strong equatorial warming and broadening and deepening of the now permanent year-round lobe of the “winter” arctic jet stream. In December 1992, ‘frigid’ arctic air was reported as far south as southern California, as were exceptionally low minimum temperatures for the first and second weeks of January 1993 [
67]. One of the effects of variability in the winter jet stream is that aberrant storm tracks are driven further south along the Pacific coastline [
68]. The arctic air mass could have come in over the top of the lower elevation marine air, moving the cold air further up the mountain, as the adjacent, lower elevation Central Valley just east of Visalia had typical daily minimum air temperatures during those events around 0 °C (PRISM daily data for 36.3815, −119.0839, 132 m; explanation suggested by R. Neilson, pers. comm.). An approach to better predicting extreme atmospheric conditions has been proposed [
69].
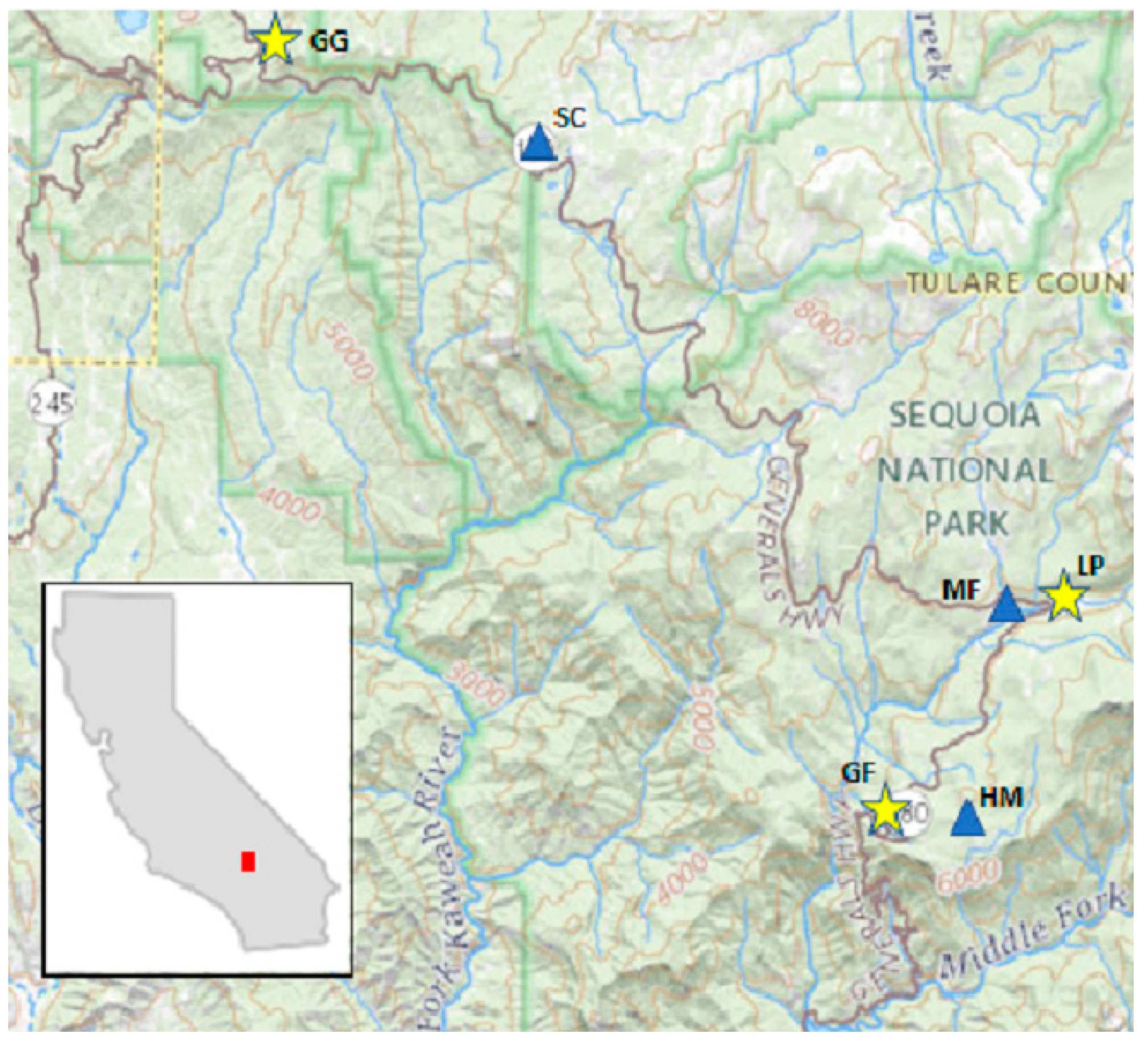
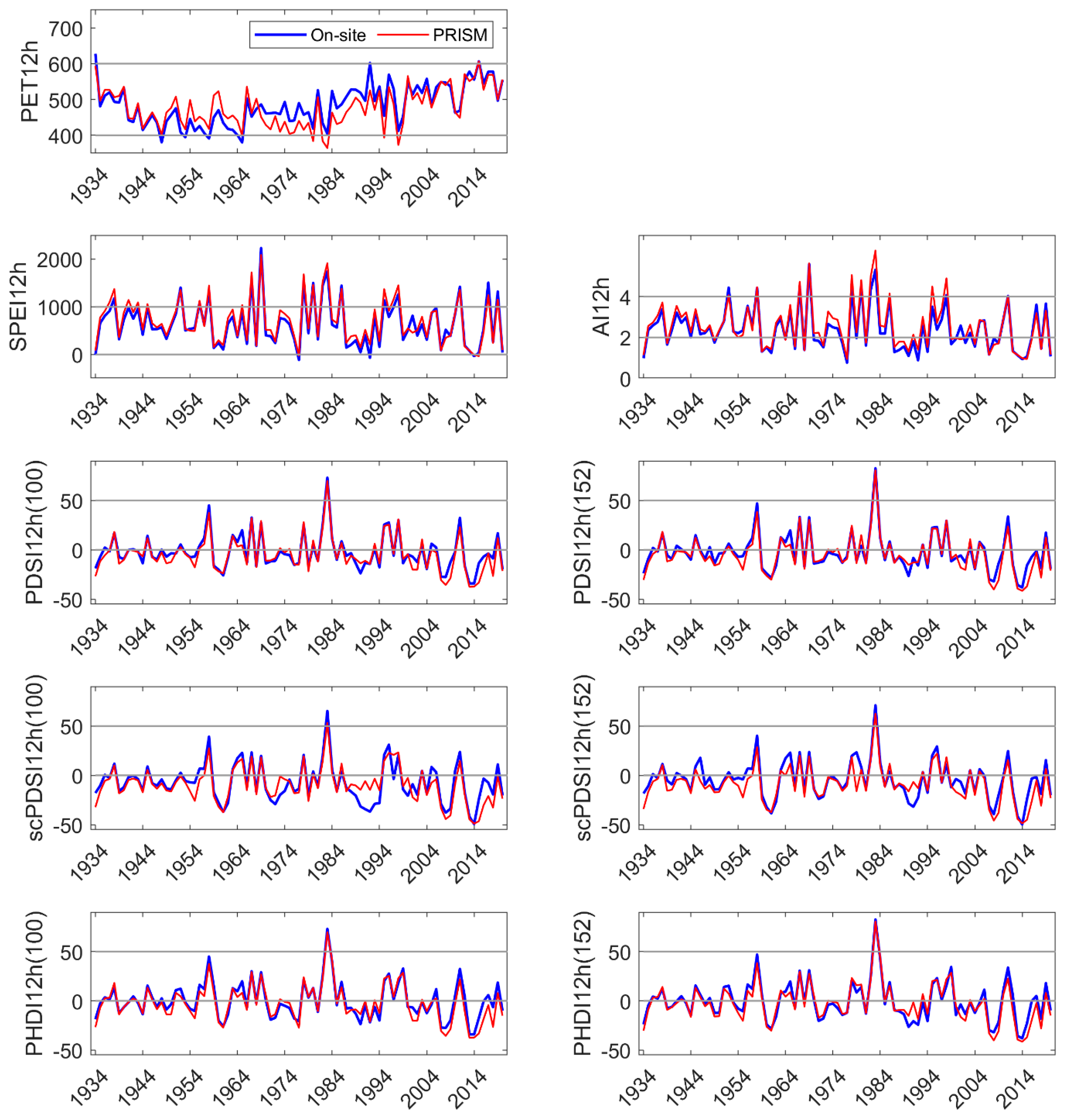
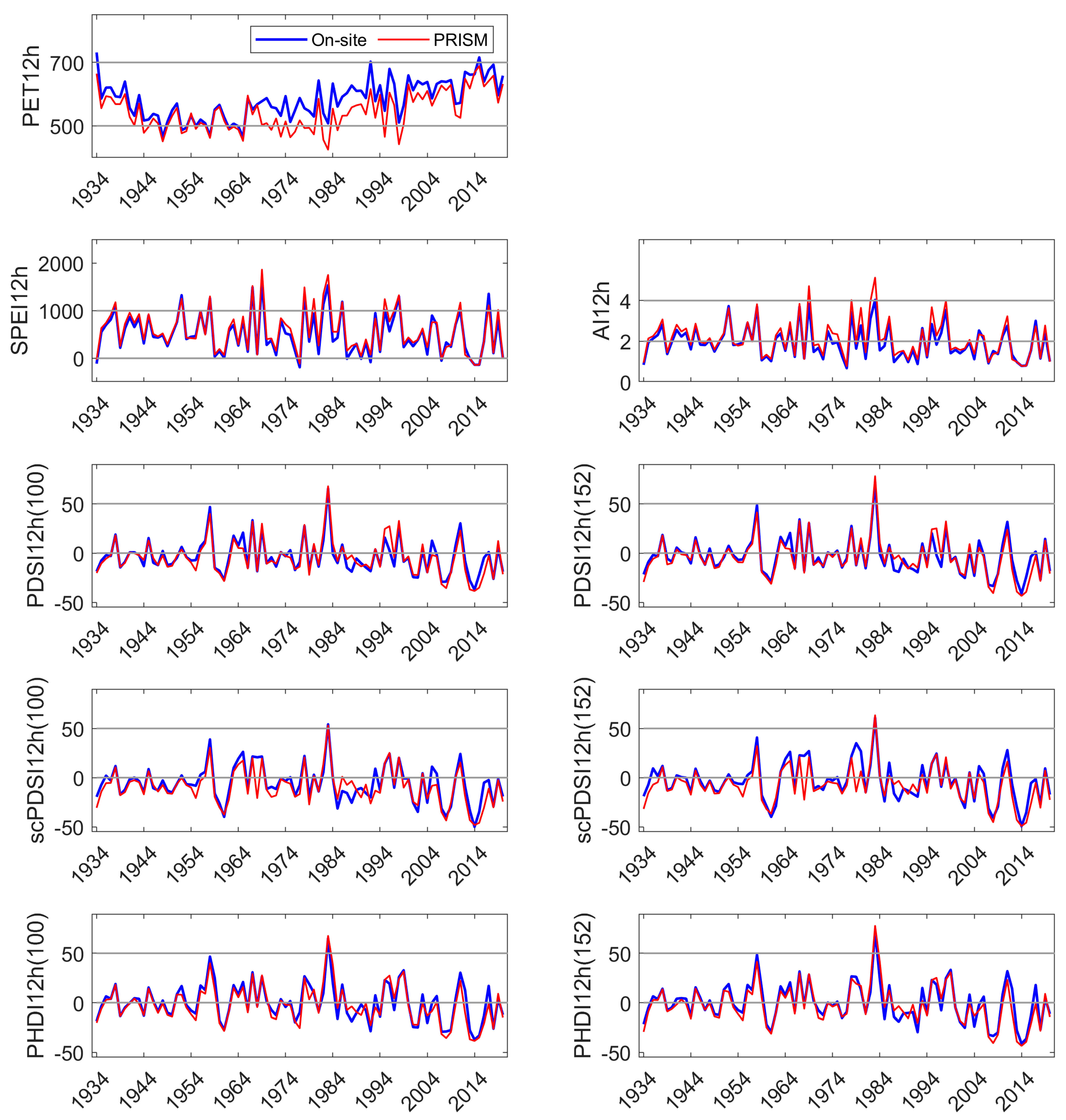
The Marble Fork Kaweah River record was short but included years of low precipitation (<60% of the long-term average) and two of the highest precipitation years of the 90 yr record (>150% of average). Current year DIs were significantly correlated to streamflow. The physics-based DIs, SPEI, and AI, had the highest correlation coefficients (>0.98). PDSI and derived indices had moderate correlation coefficients (0.72 to 0.87), with default AWC values of 152 mm performing better than 100 mm. Prior year DI, regardless of parameterization or type, was not correlated to current year streamflow perhaps because of the short distance between the headwaters and the measurement location, reducing residence and response time [
70]. In this environment with highly pulsed precipitation (90% of the annual precipitation is received between November and April) and unknown residence time, forecasts of annual streamflow can be expected to be elusive, although advances in seasonal forecasting have been made [
71].
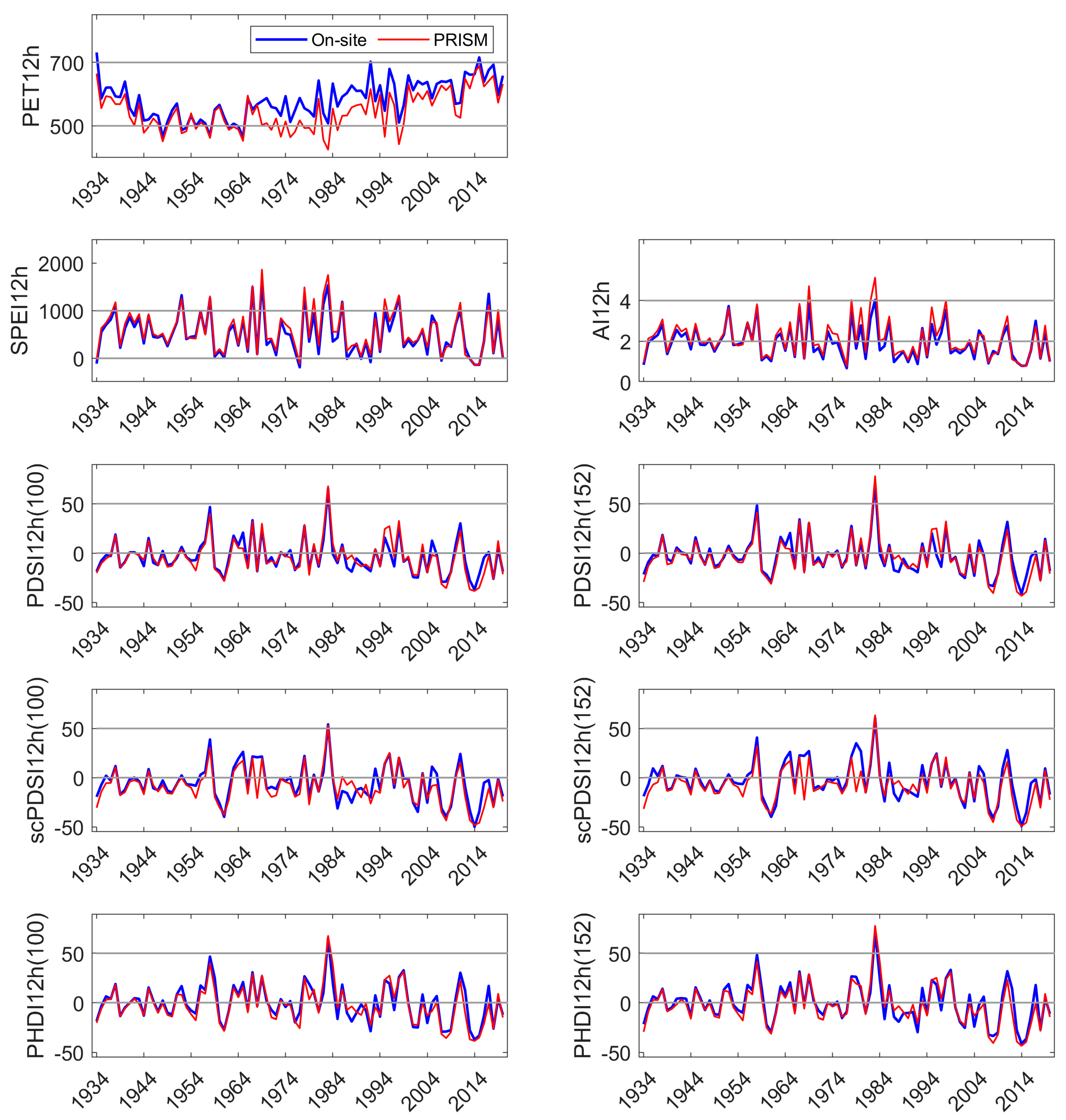
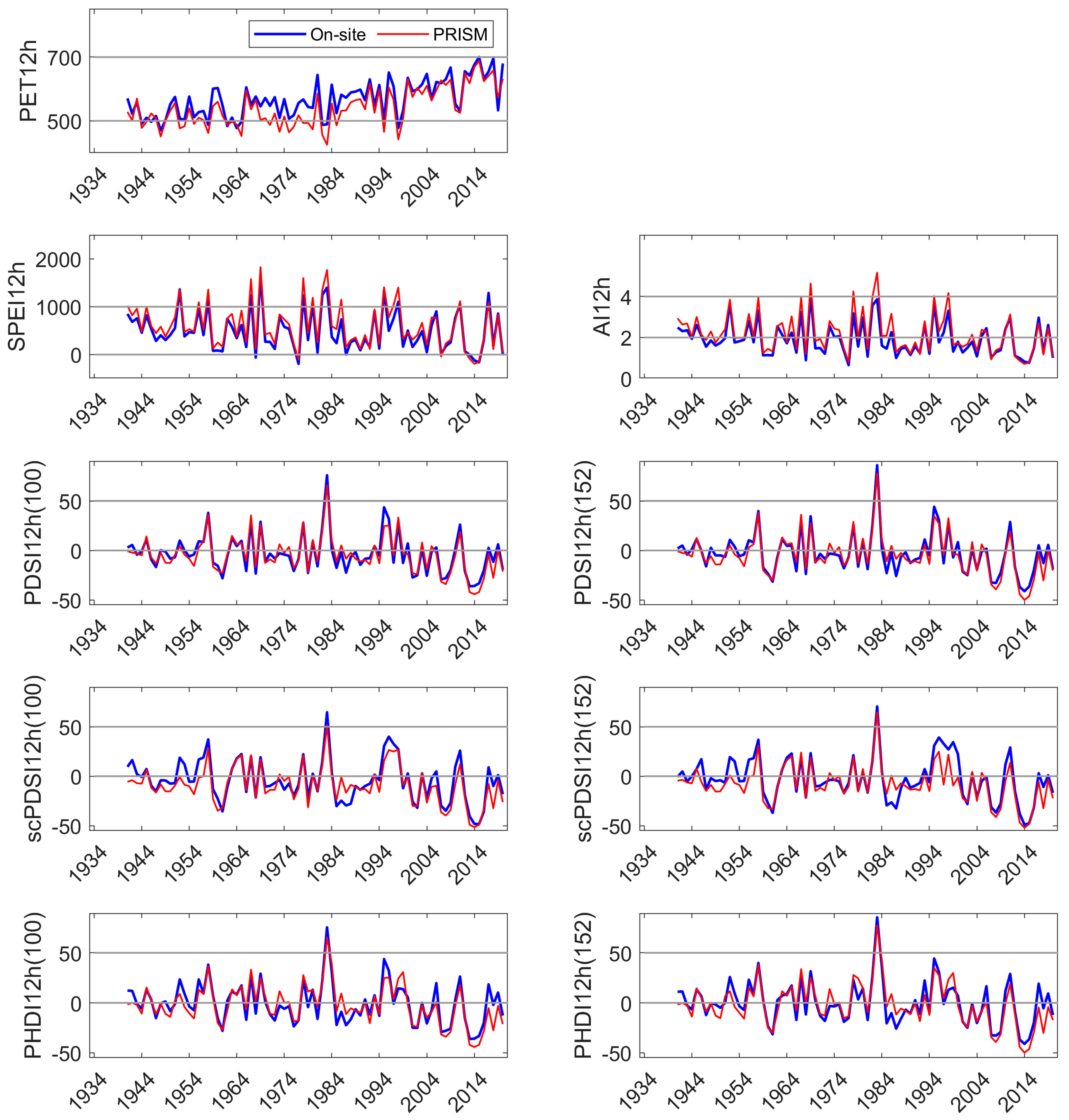
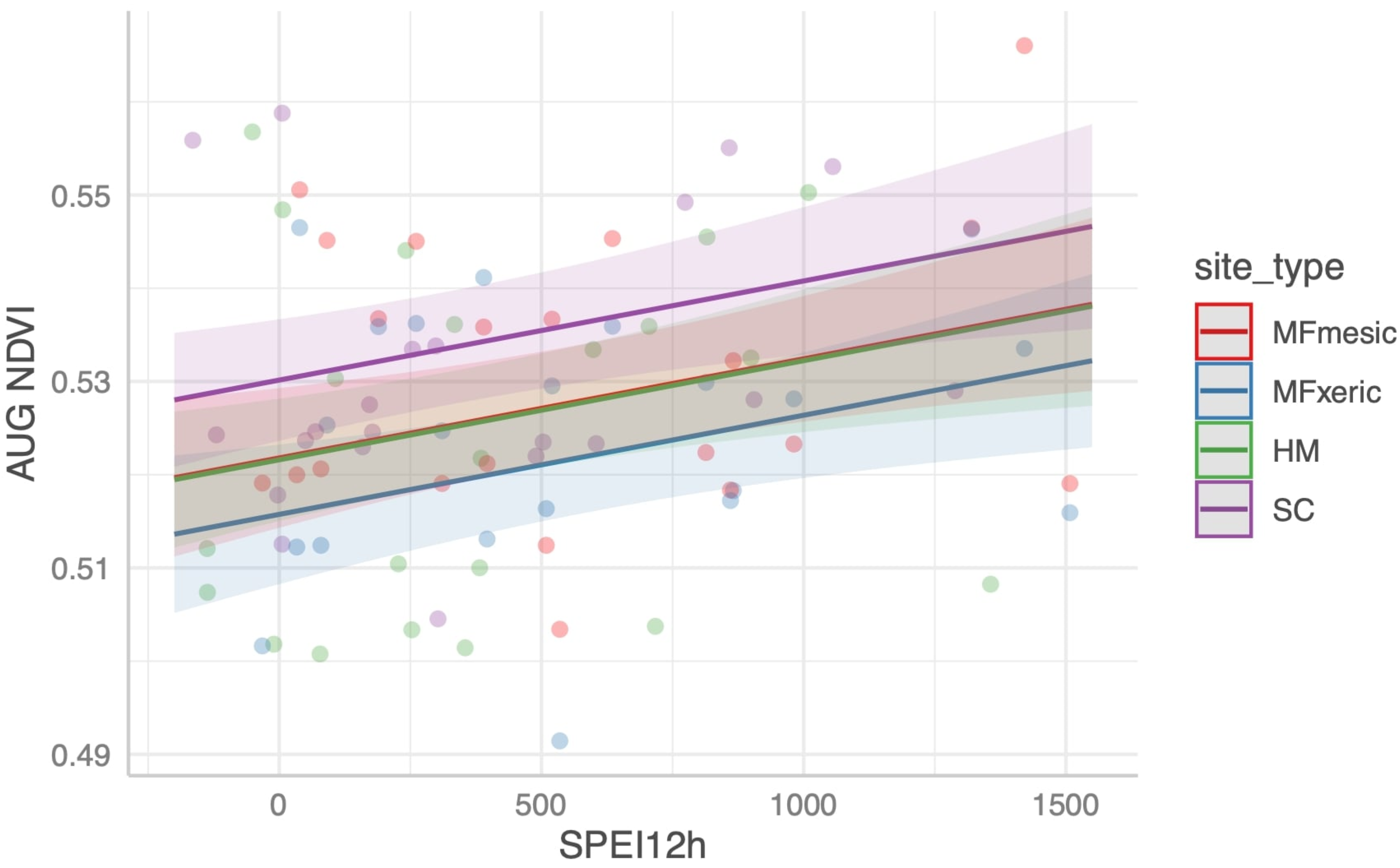
We developed a simple model using current and 1 yr lagged DIs to test their performance in predicting NDVI in the three sites. NDVI is commonly used as a proxy for vegetation productivity, a sensitive indicator of hydrologic drought [
72], and an indicator of vegetative degradation from other sources of disturbance [
73,
74]. NDVI may be particularly applicable to sites dominated by a single species, as investigated here. MODIS-derived NDVI was available for 20 yrs and, except for the 1977 drought year, contained the lowest and all but the highest precipitation year (1969) in the 86 yr precipitation record constructed [
49]. In our study, current and lagged year SPEI and PDSI
152 were significant explanatories (
p ≤ 0.01) of site NDVI, but the linear models had low R
2, whether NDVI was represented as a monthly average during period of greatest drawn-down of site water availability (August), or as an integration of site-specific, summed growing season NDVI. All indices and parameterizations performed similarly in tests of predicting NDVI (e.g., low R
2). Precipitation five months prior to the growing season (January, coinciding with the period of highest Sierra Nevada precipitation) was significantly correlated with NDVI (also MODIS-derived) [
75]. The authors suggested that a long growing season and greater carbon gain in the prior year could have also influenced gross net primary production. In another study, NDVI of individual tree crowns from a fixed-wing platform was one of a number of significant metrics in differentiating low vigor from ‘not low’ vigor crowns in a closely related pine [
76]. However, the standard deviation of crown NDVI explained more of the variation than mean crown NDVI (1st vs. 7th most important variable; Random Forest; [
76]).
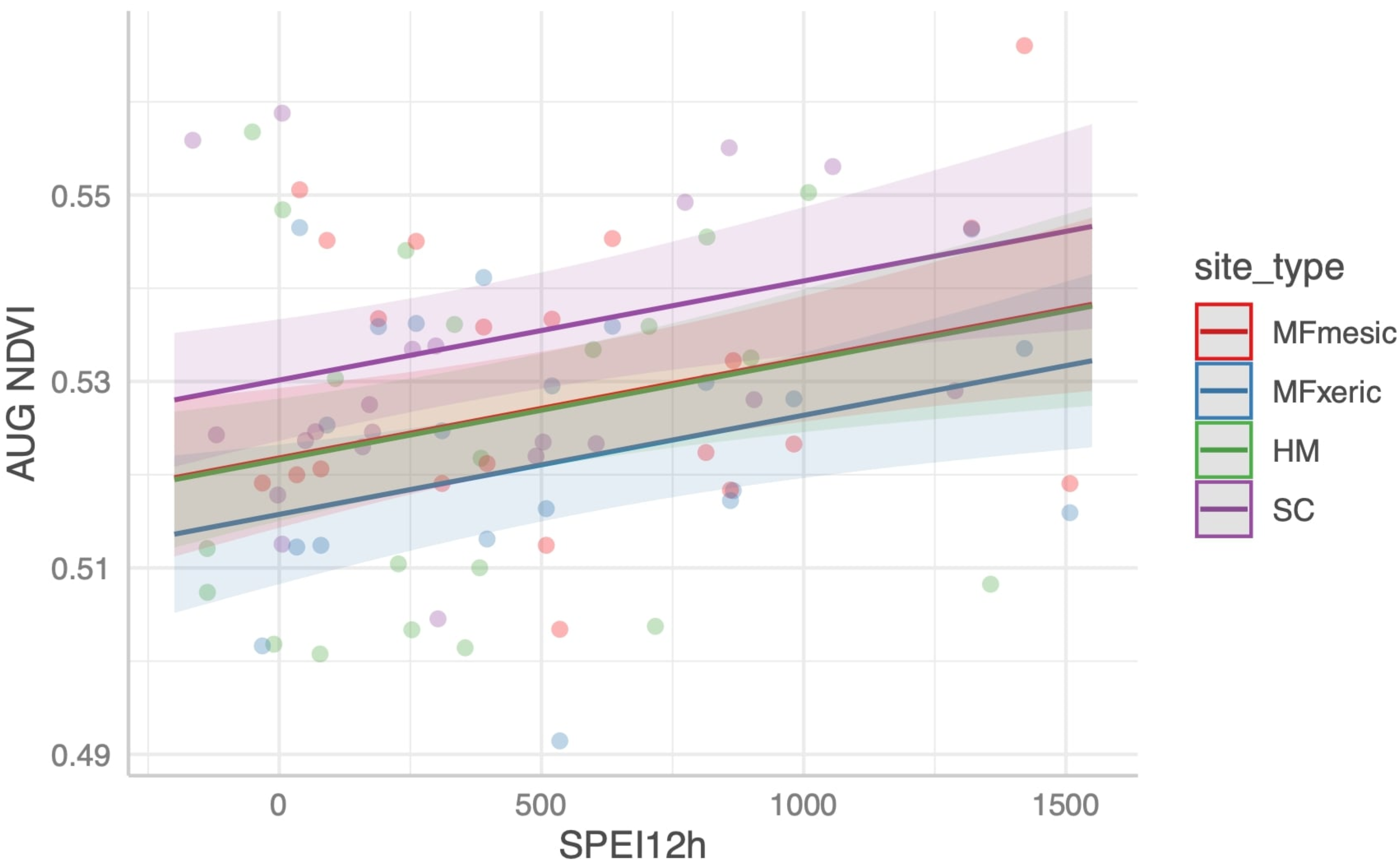
Jeffrey pine in mesic (MFm) and xeric microsites (MFx) could not be differentiated with MODIS-derived NDVI. Morphological crown response to drought (~30% loss of needles mid-crown in the 2007 drought; Grulke, unpubl. data at MF), physiological drought stress [
44], elevated foliar oxidative stress [
66], and as detected with thermal aerial imagery in 2013 ([
45]) clearly differentiated trees in the two subsites at MF. NDVI has been successfully used to validate the use of DIs in vastly different ecosystems (agricultural [
77]; chaparral, [
78]; forest [
79]; and the African continent [
80], but in the Californian mixed conifer zone with discontinuous woodlands and forest cover, neither mean August NDVI nor site-specific NDVI summed over the growing season could be used as a validation metric for DI parameterization (however, see [
77]).
We then evaluated the role of DIs and the source of their parameterizations in predicting Jeffrey pine BAI. The HM site had a shorter tree ring chronology than MF and SC and was analyzed separately. At HM, none of the five DIs, with either parameterization, at either of the AWC default values, were significant explanatories of BAI. The HM site was downslope (<0.5 km) from Crescent Meadow, which may have acted as a reservoir to buffer annual precipitation inputs, or lack thereof. It is possible that the 1987–1992 drought may not have been long or extreme enough, nor was the single drought year 2002 extreme enough, to reduce the putative supply from the meadow that might have been reflected in BAI. Proximity (distance) to a meadow accounted for 8% of the total variability in canopy water content (using Random Forest, [
81]) of giant sequoia (
Sequoiadendron giganteum Lind. & Buch.), a species that occurs adjacent to the HM site. In the same area, the underlying geology, and proximity to a stream were significant predictors of crown water content of both giant sequoia and Jeffrey pine in 2015 and 2016 [
82]. However, based on isotopic analysis [
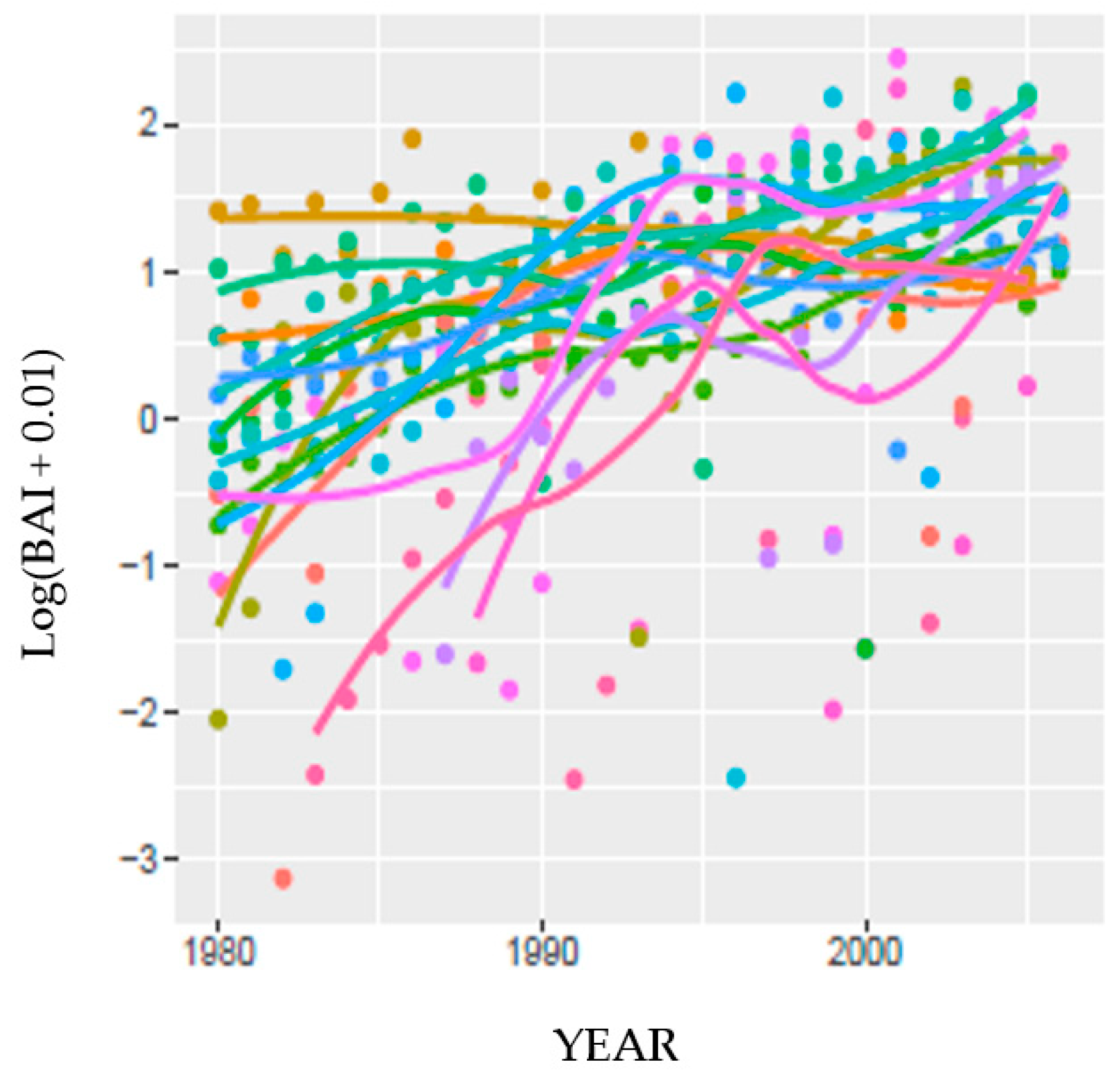
83], several deciduous desert tree species did not appear to utilize water from an adjacent stream. The trees sampled and presented here were all alive in 2019, 14 yrs after end of the tree ring record analyzed at this site. Patterns of tree BAI at HM were variable, but not more variable than at the other two sites. BAI of four HM trees had a different growth pattern suggesting reliance on a different water source than others in the stand.
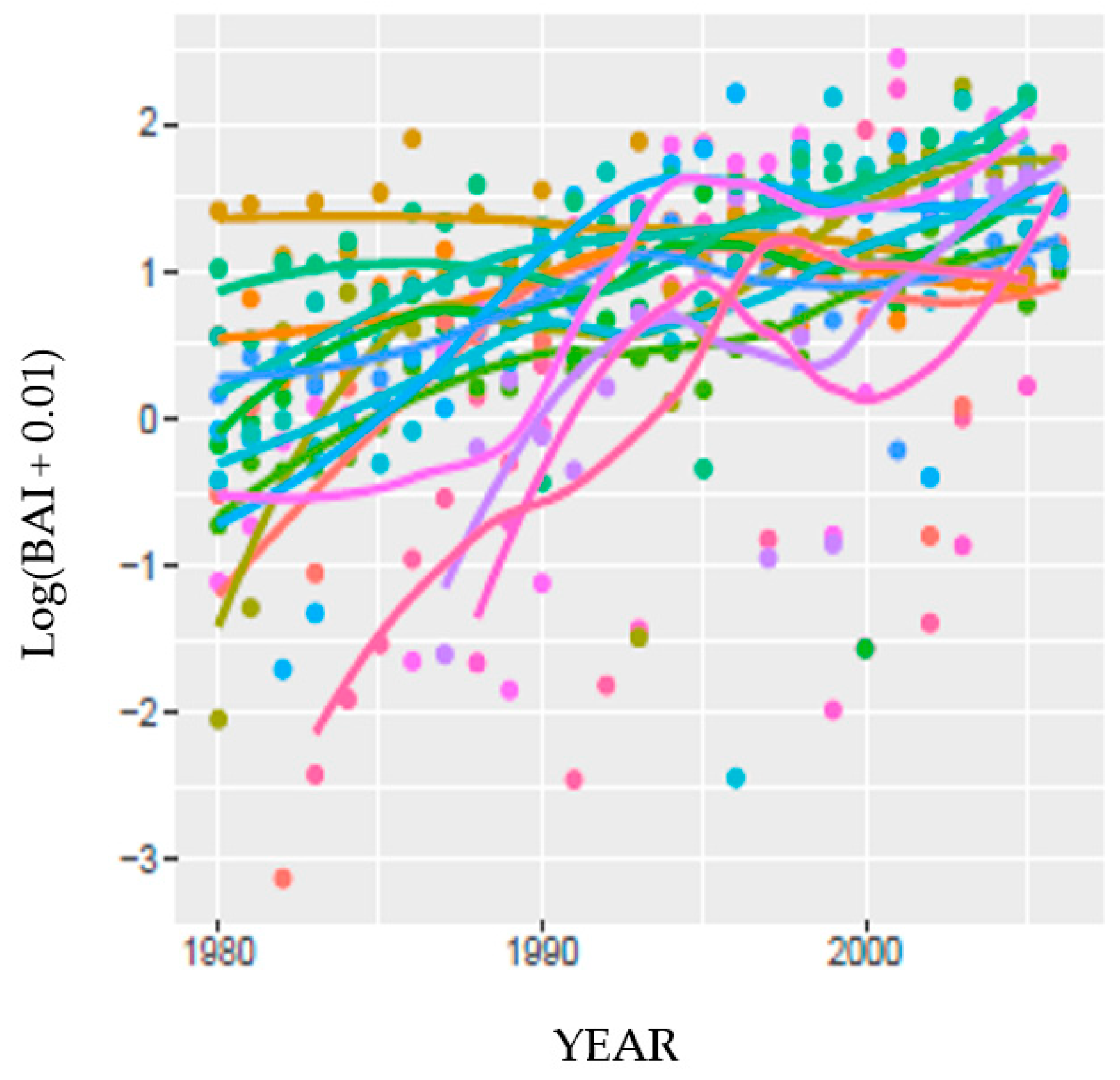
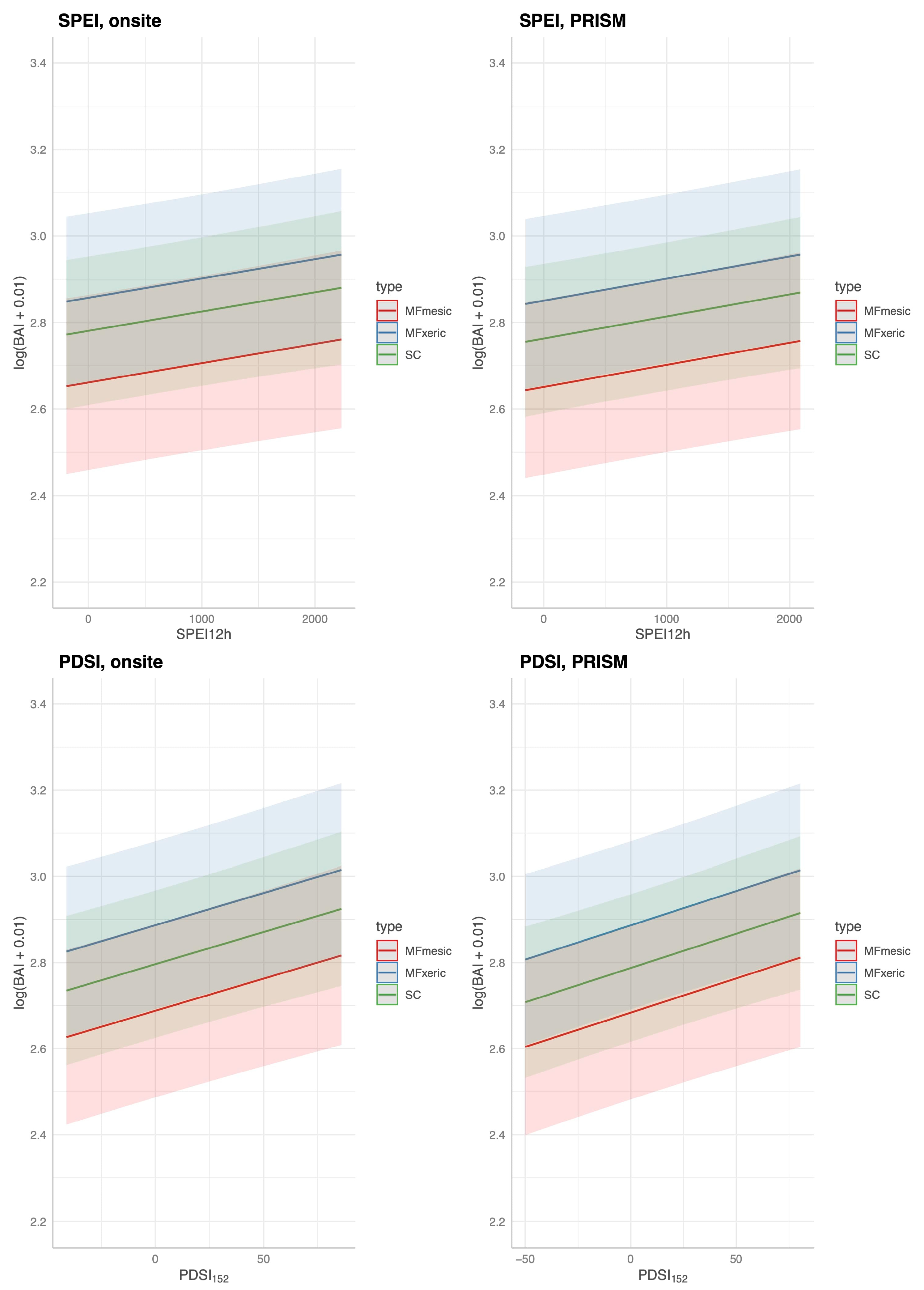
A more complex model was developed for MF and SC to predict BAI from current and temporally lagged DIs (SPEI and PDSI
152) and biological co-variates (tree crown vigor, bole growth vigor, and tree-to-tree competition). The co-variates tested for Jeffrey pine were selected from an exhaustive analysis of a closely related species, ponderosa pine [
60], and were all re-tested in this analysis of Jeffrey pine. Four of the five explanatories were significant using either DI, and the model R
2 was moderate (0.62). The fifth explanatory, tree crown vigor, was not significant with either DI or parameterization source, possibly explaining why although DIs were significantly correlated to NDVI, they explained little of the variation. Temporal lags in precipitation were also found to be significant explanatories in a model forecasting the likelihood of tree mortality due to drought and insects (bark beetle, wood borers) in the Sierra Nevada province [
84], as well as for temporally lagged SPEI for pine ring width in another summer-dry forest (for Scots pine, (
Pinus sylvestris L.), and black pine (
Pinus nigra Arn.), in Spain; [
85]). Black pine was sensitive to short term drought, but at higher elevation was less so [
86]. Jeffrey pine is known to be more drought tolerant than closely related ponderosa pine [
87], and perhaps is less sensitive to short term drought as in stone pine (
Pinus cembra L.) in the same study [
86]. Despite the possibility of long vs. short term drought sensitivities, within-growing season drought significantly affected ring width (index) in black pine. Wood properties, rather than the ring width analyzed in this study, have also been shown to be more responsive than ring widths to climate signals [
88], applicable to non-treeline locations.
The hydrology of mid-elevation sites on the western slope of the Sierra Nevada is complex. Jeffrey pine accesses moisture in shallow soil in the spring, but when near-surface horizons dry out, it likely relies on soil pockets in bedrock interstices, trapped moisture under cobbles and boulders, bedrock itself [
42,
43], surficial and underground seeps, streams, and rivers (however, see [
83]). Uphill meadows may also provide a water supply that buffers the impact of droughts [
81], as may be the case at our HM site. Individual trees in the same stand may access different water sources, which may vary with residence time, flow or percolation rates, and as well as a possibly unknowable amount of storage in soils and regolith (Cr) [
25]. Adjacent trees did not necessarily have the same pattern of growth. The converse was also true: trees in very different microsites (swales with deep soils vs. trees emerging from exposed, cracked bedrock) exhibited similar patterns of BAI. In the central Sierra Nevada, no relationship between Jeffrey pine basal area and soil thickness (A + C, nor Cr) was found [
88]. These observations suggest that trees access different sources of water and or there are temporal lags in tree response to similar sources of water.
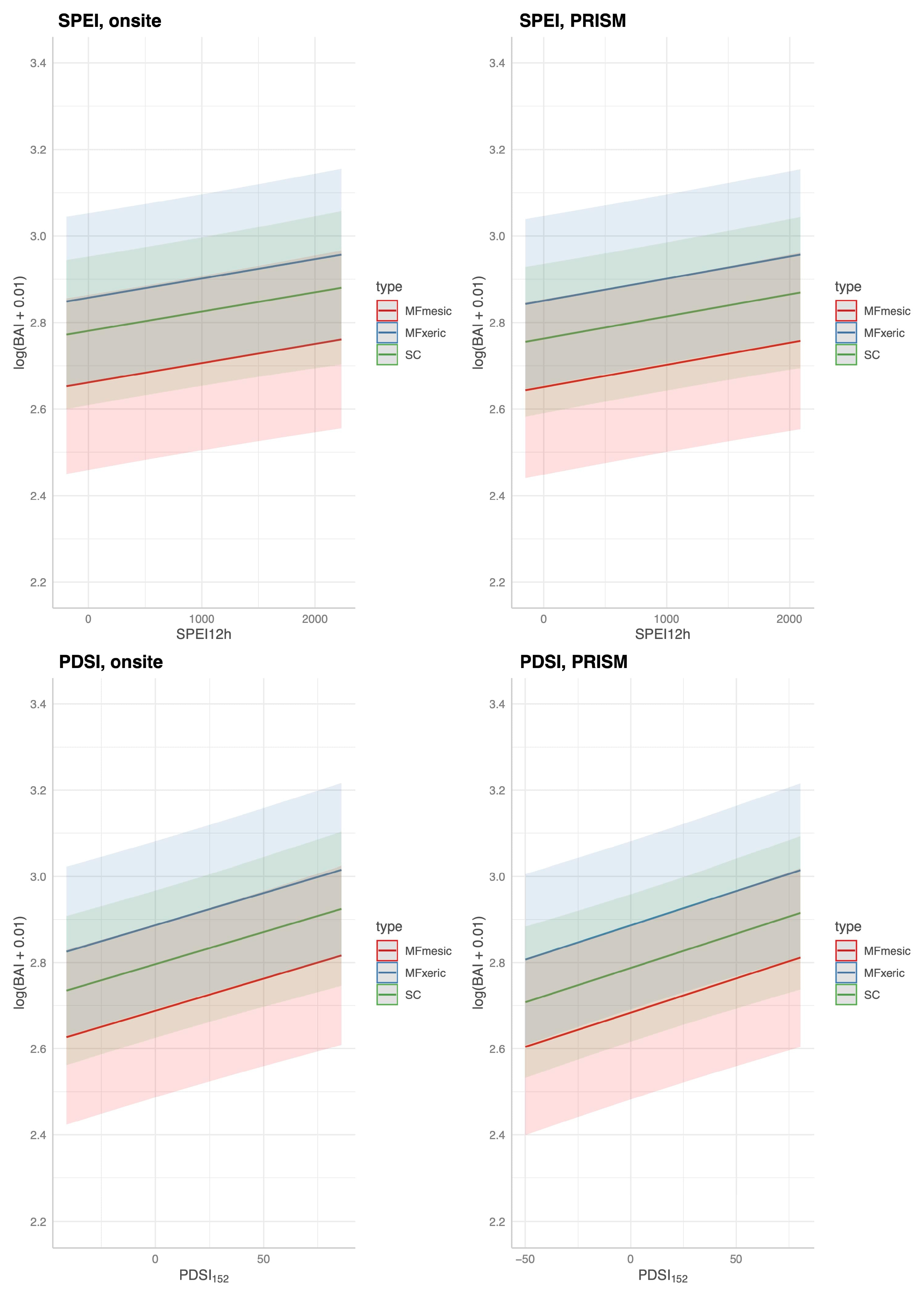
Considering the complex hydrology and largely unknown or unknowable pedology of these sites suggests that SPEI or AI would be more appropriate indices. However, for predicting BAI in our sites, there was no detectable difference between SPEI and PDSI
152 as they exhibited the same correlation coefficient with BAI. In this study, DIs alone (e.g., no biological co-variates), whether parameterized with on-site or modeled meteorological data, were not effective in predicting BAI (R
2 = 0.14), and thus in anticipating tree mortality. Surprisingly, despite the different scales of source meteorological data (microsite, point to landscape, and pixel), there was little to no statistical difference in model performance [
89].
The Sierra Nevada and Eastern Nevada Province [
84] had very high tree mortality in the 2012–2015 drought, an ‘unprecedented drought’ [
90]. Although giant sequoia grows in moist sites with deeper soils [
82,
91], and Jeffrey (and ponderosa) pine grows in thin soils overlying weathered bedrock [
24], not all trees sampled of either species responded to the drought [
81]. In giant sequoia, lower canopy water content occurred on the margins of groves, at lower elevations further from streams, or in areas of high stand density [
81]. Scots pine was more susceptible to drought-induced decline and mortality on forest edges [
92]. In our study, we observed high within-site variability BAI in the investigated sites. Adjacent trees in apparently similar microsites had differing BAI patterns, and trees with similar BAI growth patterns occurred in disparate microsites (streamside, dry convex slope). Similar to giant sequoia, some Jeffrey pine were not responsive to drought and others exhibited increased growth regardless of calculated drought severity (e.g., resilience to drought; [
93]). In this study, SPEI and PDSI
152 with on-site or modeled meteorological parameterizations, performed similarly (R
2 = 0.62, MF/SC combined) in predicting BAI only when biological covariates were included in the model. Point-in-time bole growth rates and tree-to-tree competition were significant covariates. Point-in-time bole growth rate, here barkless DBH as a function of tree age at a mid-point in the analysis, was a significant co-variate. In a much larger analysis, larger trees had a disproportionate competitive advantage in mixed age conifer forests [
39], and trees with higher bole growth rates were less likely to respond to drought defined by PDSI. The important role of stand- and tree-level competition in bole growth rates has long been established [
94,
95], and ‘neighborhood’ competition (CZD) was also a significant co-variate in our model.
Trees may succumb quickly or at length to a physical or biological assault. Manion’s [
35] ‘death spiral’ suggests mortality in 5 to 8 yrs with multiple insults in long-lived trees. In south central Oregon, ponderosa pine was found to take 40 yrs to succumb after an inciting drought, based on steadily declining BAI to nil (Grulke and Mershel, unpubl. data). Jeffrey pines died in the same year on the eastern edge of the Transverse Range in 1999, the first year of a multi-year drought (1999–2002) (Grulke, pers. obs.). In that range, 25% to 75% tree mortality occurred in stands with greater than 50% tree cover [
96]. In the Sierra Nevada, the ‘background’ tree mortality from 2010 to 2014 was 2.7 million trees annually, preceded by a decade of already lower precipitation and higher temperatures than long term records. The 2013, 2014, and 2015 mortality rates were 29, 62, and 27 million trees, respectively, dropping to 19 million trees in 2018 [
6], and still elevated 3 to 6 yrs after prolonged ‘hot’ drought as ground water deficits lag precipitation deficits [
36].
We can monitor declining canopy water content [
97] and pre-emptively anticipate physiological tree drought stress by gaging within-crown thermal differences [
45] or oxidative stress (PRI, xanthophyll cycle [
98]), but the analysis presented here suggests that biological co-variates, bole growth vigor and tree-to-tree competition, aid in predicting declining BAI. Only the latter co-variate can be assessed remotely. Continuity of mature tree canopies (e.g., greater cover) may be assessed with LiDAR, which is increasingly monitored and available over forested regions. At the California state-wide level [
79], a change metric was successfully identified using NDVI that was associated with increased tree mortality 6 to 19 months later [
99]. Crown vigor was correlated to ‘bole vigor’ in an analysis of ponderosa pine [
60] but was not a significant variant in our model predicting BAI, whether or not bole vigor was also included. The predictive capacity of our model dropped from moderate to poor when bole vigor was not concurrently considered.
Anticipating the meteorological conditions that will lead to extensive tree die-offs in mid- to upper elevations is needed, as loss of vegetation cover will lead to lower on-site water retention and ‘flashier’ downslope flows. DIs alone do not appear to be particularly effective in predicting tree bole growth, an attribute that declines as trees succumb. Conditions and thresholds have been defined that lead to hydraulic failure in trees [
100], but the approach may not be scalable in mid to upper elevation forests that rely on regolithic water in the last third to half of the growing season. Estimating the rate of decline, year over year in NDVI [
79,
100], oxidative stress [
99], and or crown or sub-crown thermal imagery [
45] could be used as a harbinger of tree decline and mortality, and perhaps with sufficient lead time, for mitigation. We can anticipate loss of water resources with DIs, but at present we only monitor the degradation of mid elevation, open forest Sierran ecosystems with lower water availability and higher temperatures.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}