
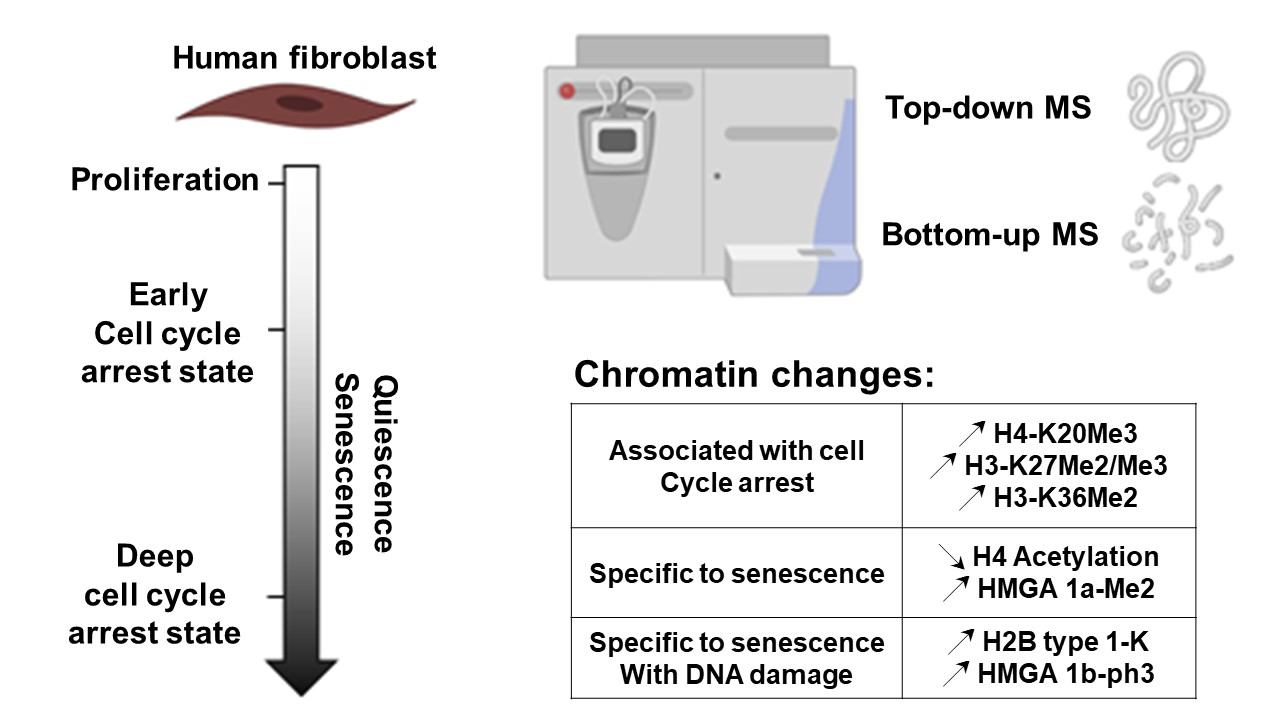
H2B Type 1-K Accumulates in Senescent Fibroblasts with Persistent DNA Damage along with Methylated and Phosphorylated Forms of HMGA1



Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
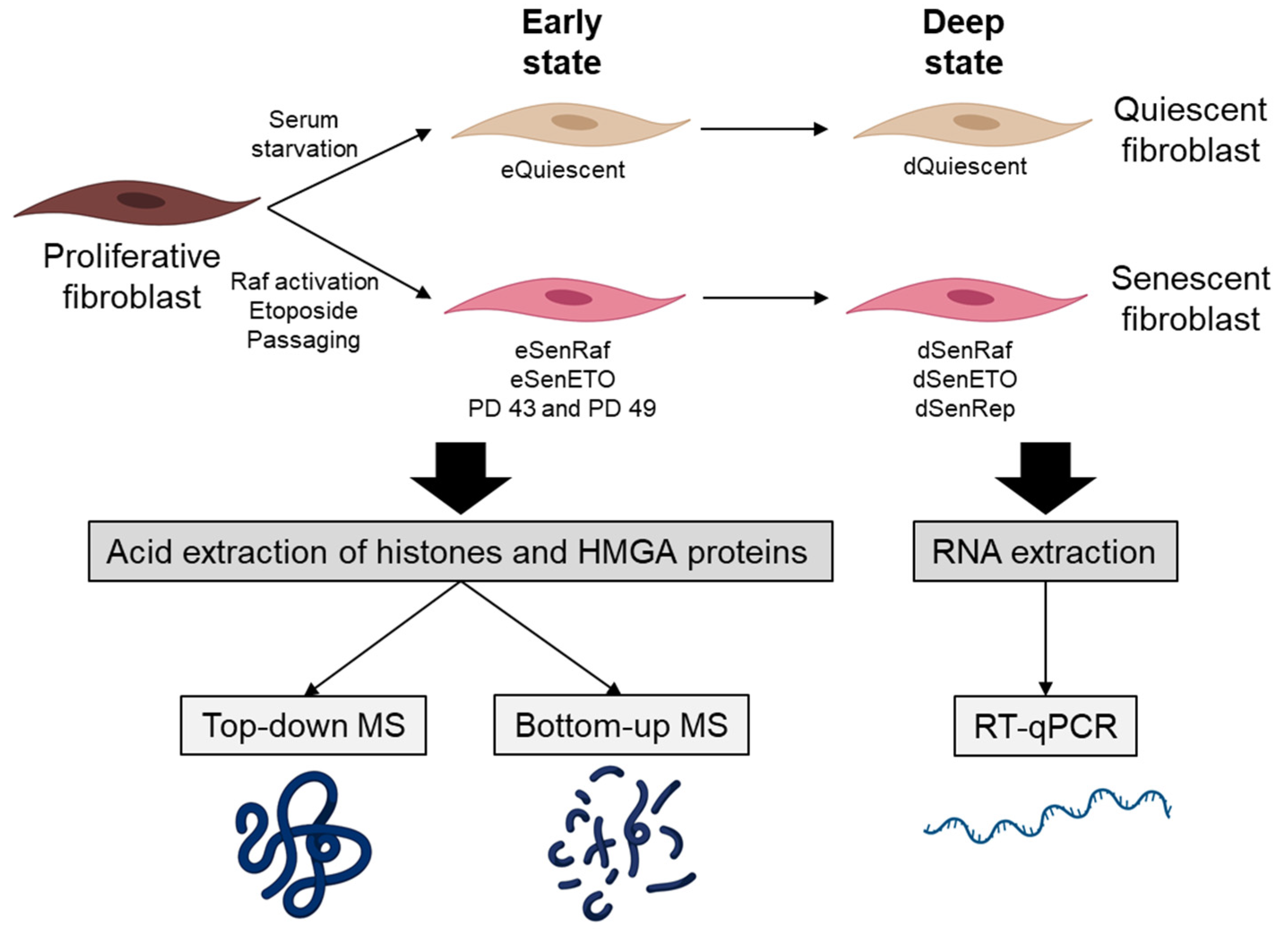
2. Materials and Methods
2.1. Cell Lines and Retroviruses
2.2. Chemicals
2.3. Preparation of Histones and HMGA Proteins, and Mass Spectrometry Analyses
2.4. Flow Cytometry Analyses of DNA Content
2.5. BrdU Incorporation and Immunostaining
2.6. qRT-PCR
2.7. Data Analysis and Statistics
3. Results
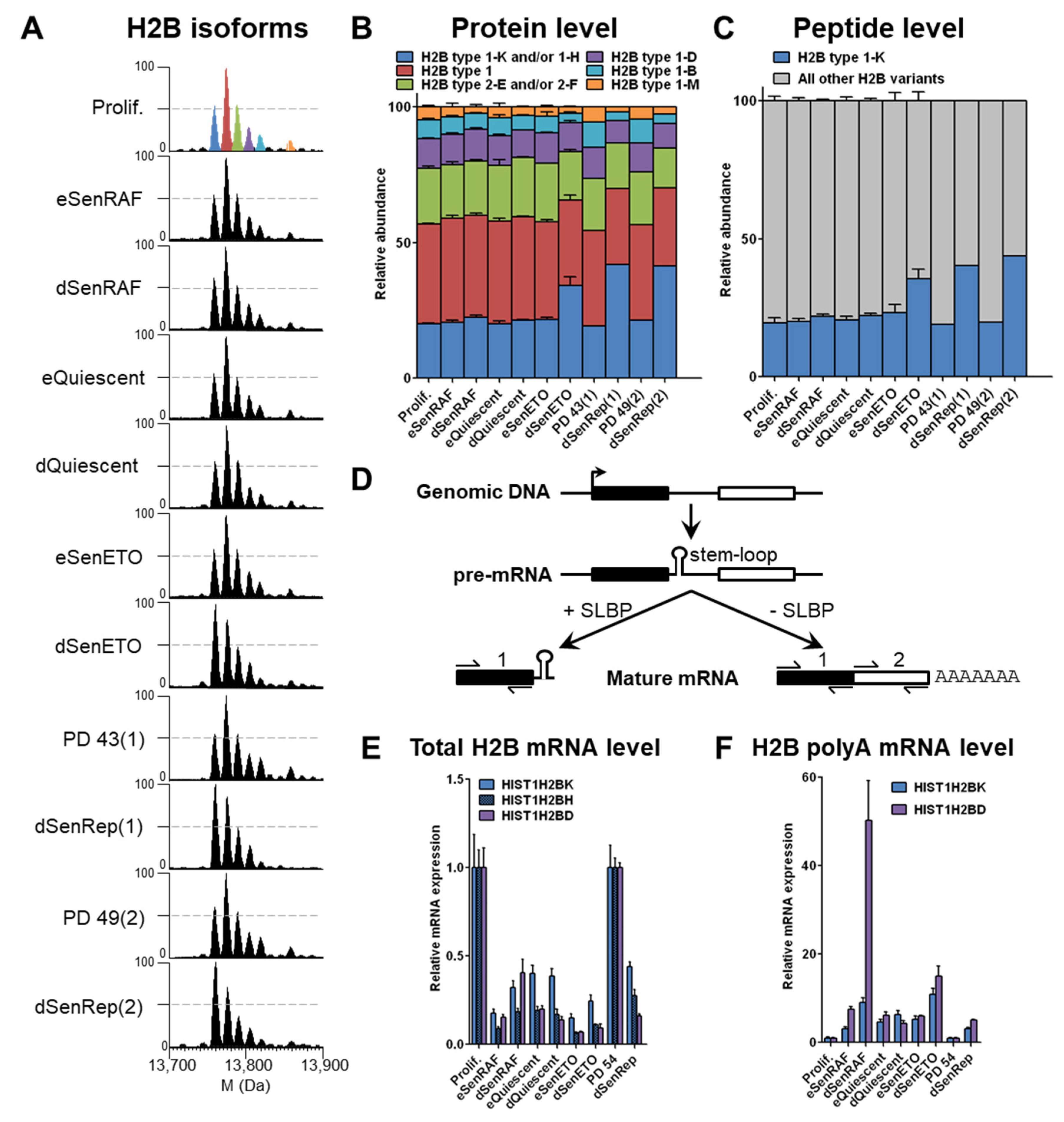
3.1. H2B type1-K Is Specifically Enriched in Deep Senescent Conditions with Persistent DNA Damage by an Active Post-Transcriptional Regulation
3.2. H4 Mono-Acetylation Remained Low in Deep Senescent States and H4-K20Me3 Increased Progressively with Time in Conditions of Cell Cycle Arrests
3.3. H3.1/2-K27Me2/Me3 and K36Me2 Accumulate with Time in Conditions of Cell Cycle Arrest
3.4. HMGA1a Di-Methylation and HMGA1b Tri-Phosphorylation Accumulated in Deep Senescent Conditions
4. Discussion
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kuilman, T.; Michaloglou, C.; Mooi, W.J.; Peeper, D.S. The essence of senescence. Genes Dev. 2010, 24, 2463–2479. [Google Scholar] [CrossRef] [Green Version]
- Rodier, F.; Campisi, J. Four faces of cellular senescence. J. Cell Biol. 2011, 192, 547–556. [Google Scholar] [CrossRef]
- Berger, S.L. The complex language of chromatin regulation during transcription. Nature 2007, 447, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Groth, A.; Corpet, A.; Cook, A.J.L.; Roche, D.; Bartek, J.; Lukas, J.; Almouzni, G. Regulation of replication fork progression through histone supply and demand. Science 2007, 318, 1928–1931. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Carey, M.; Workman, J.L. The role of chromatin during transcription. Cell 2007, 128, 707–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, V.W.; Goren, A.; Bernstein, B.E. Charting histone modifications and the functional organization of mammalian genomes. Nat. Rev. Genet. 2011, 12, 7–18. [Google Scholar] [CrossRef]
- Boyne, M.T.; Pesavento, J.J.; Mizzen, C.A.; Kelleher, N.L. Precise characterization of human histones in the H2A gene family by top down mass spectrometry. J. Proteome Res. 2006, 5, 248–253. [Google Scholar] [CrossRef]
- Siuti, N.; Roth, M.J.; Mizzen, C.A.; Kelleher, N.L.; Pesavento, J.J. Gene-specific characterization of human histone H2B by electron capture dissociation. J. Proteome Res. 2006, 5, 233–239. [Google Scholar] [CrossRef]
- Shah, S.G.; Mandloi, T.; Kunte, P.; Natu, A.; Rashid, M.; Reddy, D.; Gadewal, N.; Gupta, S. HISTome2: A database of histone proteins, modifiers for multiple organisms and epidrugs. Epigenet. Chromatin 2020, 13, 31. [Google Scholar] [CrossRef]
- Long, M.; Sun, X.; Shi, W.; Yanru, A.; Leung, S.T.C.; Ding, D.; Cheema, M.S.; MacPherson, N.; Nelson, C.J.; Ausio, J.; et al. A novel histone H4 variant H4G regulates rDNA transcription in breast cancer. Nucleic Acids Res. 2019, 47, 8399–8409. [Google Scholar] [CrossRef] [Green Version]
- Talbert, P.B.; Henikoff, S. Histone variants—Ancient wrap artists of the epigenome. Nat. Rev. Mol. Cell Biol. 2010, 11, 264–275. [Google Scholar] [CrossRef]
- Marzluff, W.F.; Wagner, E.J.; Duronio, R.J. Metabolism and regulation of canonical histone mRNAs: Life without a poly(A) tail. Nat. Rev. Genet. 2008, 9, 843–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kari, V.; Karpiuk, O.; Tieg, B.; Kriegs, M.; Dikomey, E.; Krebber, H.; Begus-Nahrmann, Y.; Johnsen, S.A. A subset of histone H2B genes produces polyadenylated mRNAs under a variety of cellular conditions. PLoS ONE 2013, 8, e63745. [Google Scholar] [CrossRef] [PubMed]
- Rai, T.S.; Cole, J.J.; Nelson, D.M.; Dikovskaya, D.; Faller, W.J.; Vizioli, M.G.; Hewitt, R.N.; Anannya, O.; McBryan, T.; Manoharan, I.; et al. HIRA orchestrates a dynamic chromatin landscape in senescence and is required for suppression of neoplasia. Genes Dev. 2014, 28, 2712–2725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molden, R.C.; Bhanu, N.V.; LeRoy, G.; Arnaudo, A.M.; Garcia, B.A. Multi-faceted quantitative proteomics analysis of histone H2B isoforms and their modifications. Epigenet. Chromatin 2015, 8, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyons, S.M.; Cunningham, C.H.; Welch, J.D.; Groh, B.; Guo, A.Y.; Wei, B.; Whitfield, M.L.; Xiong, Y.; Marzluff, W.F. A subset of replication-dependent histone mRNAs are expressed as polyadenylated RNAs in terminally differentiated tissues. Nucleic Acids Res. 2016, 44, 9190–9205. [Google Scholar] [CrossRef] [Green Version]
- De Koning, L.; Corpet, A.; Haber, J.E.; Almouzni, G. Histone chaperones: An escort network regulating histone traffic. Nat. Struct. Mol. Biol. 2007, 14, 997–1007. [Google Scholar] [CrossRef]
- Zhang, R.; Poustovoitov, M.V.; Ye, X.; Santos, H.A.; Chen, W.; Daganzo, S.M.; Erzberger, J.P.; Serebriiskii, I.G.; Canutescu, A.A.; Dunbrack, R.L.; et al. Formation of MacroH2A-containing senescence-associated heterochromatin foci and senescence driven by ASF1a and HIRA. Dev. Cell 2005, 8, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Lewis, P.W.; Elsaesser, S.J.; Noh, K.-M.; Stadler, S.C.; Allis, C.D. Daxx is an H3.3-specific histone chaperone and cooperates with ATRX in replication-independent chromatin assembly at telomeres. Proc. Natl. Acad. Sci. USA 2010, 107, 14075–14080. [Google Scholar] [CrossRef] [Green Version]
- Kappes, F.; Waldmann, T.; Mathew, V.; Yu, J.; Zhang, L.; Khodadoust, M.S.; Chinnaiyan, A.M.; Luger, K.; Erhardt, S.; Schneider, R.; et al. The DEK oncoprotein is a Su(var) that is essential to heterochromatin integrity. Genes Dev. 2011, 25, 673–678. [Google Scholar] [CrossRef] [Green Version]
- Contrepois, K.; Ezan, E.; Mann, C.; Fenaille, F. Ultra-high performance liquid chromatography-mass spectrometry for the fast profiling of histone post-translational modifications. J. Proteome Res. 2010, 9, 5501–5509. [Google Scholar] [CrossRef]
- Contrepois, K.; Coudereau, C.; Benayoun, B.A.; Schuler, N.; Roux, P.-F.; Bischof, O.; Courbeyrette, R.; Carvalho, C.; Thuret, J.-Y.; Ma, Z.; et al. Histone variant H2A.J accumulates in senescent cells and promotes inflammatory gene expression. Nat. Commun. 2017, 8, 14995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chicas, A.; Kapoor, A.; Wang, X.; Aksoy, O.; Evertts, A.G.; Zhang, M.Q.; Garcia, B.A.; Bernstein, E.; Lowe, S.W. H3K4 demethylation by Jarid1a and Jarid1b contributes to retinoblastoma-mediated gene silencing during cellular senescence. Proc. Natl. Acad. Sci. USA 2012, 109, 8971–8976. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.M.; Jaber-Hijazi, F.; Cole, J.J.; Robertson, N.A.; Pawlikowski, J.S.; Norris, K.T.; Criscione, S.W.; Pchelintsev, N.A.; Piscitello, D.; Stong, N.; et al. Mapping H4K20me3 onto the chromatin landscape of senescent cells indicates a function in control of cell senescence and tumor suppression through preservation of genetic and epigenetic stability. Genome Biol. 2016, 17, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piunti, A.; Shilatifard, A. The roles of Polycomb repressive complexes in mammalian development and cancer. Nat. Rev. Mol. Cell Biol. 2021, 22, 326–345. [Google Scholar] [CrossRef]
- Li, J.; Ahn, J.H.; Wang, G.G. Understanding histone H3 lysine 36 methylation and its deregulation in disease. Cell. Mol. Life Sci. CMLS 2019, 76, 2899–2916. [Google Scholar] [CrossRef] [PubMed]
- Narita, M.; Narita, M.; Krizhanovsky, V.; Nuñez, S.; Chicas, A.; Hearn, S.A.; Myers, M.P.; Lowe, S.W. A novel role for high-mobility group a proteins in cellular senescence and heterochromatin formation. Cell 2006, 126, 503–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contrepois, K.; Thuret, J.-Y.; Courbeyrette, R.; Fenaille, F.; Mann, C. Deacetylation of H4-K16Ac and heterochromatin assembly in senescence. Epigenet. Chromatin 2012, 5, 15. [Google Scholar] [CrossRef] [Green Version]
- Jeanblanc, M.; Ragu, S.; Gey, C.; Contrepois, K.; Courbeyrette, R.; Thuret, J.-Y.; Mann, C. Parallel pathways in RAF-induced senescence and conditions for its reversion. Oncogene 2012, 31, 3072–3085. [Google Scholar] [CrossRef] [Green Version]
- Smogorzewska, A.; de Lange, T. Different telomere damage signaling pathways in human and mouse cells. EMBO J. 2002, 21, 4338–4348. [Google Scholar] [CrossRef] [Green Version]
- Garcia, B.A.; Mollah, S.; Ueberheide, B.M.; Busby, S.A.; Muratore, T.L.; Shabanowitz, J.; Hunt, D.F. Chemical derivatization of histones for facilitated analysis by mass spectrometry. Nat. Protoc. 2007, 2, 933–938. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Garcia, B.A. Examining histone posttranslational modification patterns by high-resolution mass spectrometry. Methods Enzymol. 2012, 512, 3–28. [Google Scholar] [CrossRef] [Green Version]
- Sidoli, S.; Bhanu, N.V.; Karch, K.R.; Wang, X.; Garcia, B.A. Complete workflow for analysis of histone post-translational modifications using bottom-up mass spectrometry: From histone extraction to data analysis. J. Vis. Exp. JoVE 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evertts, A.G.; Manning, A.L.; Wang, X.; Dyson, N.J.; Garcia, B.A.; Coller, H.A. H4K20 methylation regulates quiescence and chromatin compaction. Mol. Biol. Cell 2013, 24, 3025–3037. [Google Scholar] [CrossRef] [PubMed]
- Garcia, B.A.; Thomas, C.E.; Kelleher, N.L.; Mizzen, C.A. Tissue-specific expression and post-translational modification of histone H3 variants. J. Proteome Res. 2008, 7, 4225–4236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edberg, D.D.; Adkins, J.N.; Springer, D.L.; Reeves, R. Dynamic and differential in vivo modifications of the isoform HMGA1a and HMGA1b chromatin proteins. J. Biol. Chem. 2005, 280, 8961–8973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, J.C.; Zamdborg, L.; Ahlf, D.R.; Lee, J.E.; Catherman, A.D.; Durbin, K.R.; Tipton, J.D.; Vellaichamy, A.; Kellie, J.F.; Li, M.; et al. Mapping intact protein isoforms in discovery mode using top-down proteomics. Nature 2011, 480, 254–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sgarra, R.; Diana, F.; Bellarosa, C.; Dekleva, V.; Rustighi, A.; Toller, M.; Manfioletti, G.; Giancotti, V. During apoptosis of tumor cells HMGA1a protein undergoes methylation: Identification of the modification site by mass spectrometry. Biochemistry 2003, 42, 3575–3585. [Google Scholar] [CrossRef]
- Zou, Y.; Wang, Y. Tandem mass spectrometry for the examination of the posttranslational modifications of high-mobility group A1 proteins: Symmetric and asymmetric dimethylation of Arg25 in HMGA1a protein. Biochemistry 2005, 44, 6293–6301. [Google Scholar] [CrossRef]
- Young, N.L.; Plazas-Mayorca, M.D.; DiMaggio, P.A.; Flaniken, I.Z.; Beltran, A.J.; Mishra, N.; LeRoy, G.; Floudas, C.A.; Garcia, B.A. Collective mass spectrometry approaches reveal broad and combinatorial modification of high mobility group protein A1a. J. Am. Soc. Mass Spectrom. 2010, 21, 960–970. [Google Scholar] [CrossRef] [Green Version]
- Lorain, S.; Quivy, J.P.; Monier-Gavelle, F.; Scamps, C.; Lécluse, Y.; Almouzni, G.; Lipinski, M. Core histones and HIRIP3, a novel histone-binding protein, directly interact with WD repeat protein HIRA. Mol. Cell. Biol. 1998, 18, 5546–5556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardal, A.J.; Fernandes-Duarte, F.; Bowman, A.J. The histone chaperoning pathway: From ribosome to nucleosome. Essays Biochem. 2019, 63, 29–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarg, B.; Koutzamani, E.; Helliger, W.; Rundquist, I.; Lindner, H.H. Postsynthetic trimethylation of histone H4 at lysine 20 in mammalian tissues is associated with aging. J. Biol. Chem. 2002, 277, 39195–39201. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Cheung, T.H.; Charville, G.W.; Hurgo, B.M.C.; Leavitt, T.; Shih, J.; Brunet, A.; Rando, T.A. Chromatin modifications as determinants of muscle stem cell quiescence and chronological aging. Cell Rep. 2013, 4, 189–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tvardovskiy, A.; Schwämmle, V.; Kempf, S.J.; Rogowska-Wrzesinska, A.; Jensen, O.N. Accumulation of histone variant H3.3 with age is associated with profound changes in the histone methylation landscape. Nucleic Acids Res. 2017, 45, 9272–9289. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Contrepois, K.; Mann, C.; Fenaille, F. H2B Type 1-K Accumulates in Senescent Fibroblasts with Persistent DNA Damage along with Methylated and Phosphorylated Forms of HMGA1. Proteomes 2021, 9, 30. https://doi.org/10.3390/proteomes9020030
Contrepois K, Mann C, Fenaille F. H2B Type 1-K Accumulates in Senescent Fibroblasts with Persistent DNA Damage along with Methylated and Phosphorylated Forms of HMGA1. Proteomes. 2021; 9(2):30. https://doi.org/10.3390/proteomes9020030
Chicago/Turabian StyleContrepois, Kévin, Carl Mann, and François Fenaille. 2021. "H2B Type 1-K Accumulates in Senescent Fibroblasts with Persistent DNA Damage along with Methylated and Phosphorylated Forms of HMGA1" Proteomes 9, no. 2: 30. https://doi.org/10.3390/proteomes9020030
APA StyleContrepois, K., Mann, C., & Fenaille, F. (2021). H2B Type 1-K Accumulates in Senescent Fibroblasts with Persistent DNA Damage along with Methylated and Phosphorylated Forms of HMGA1. Proteomes, 9(2), 30. https://doi.org/10.3390/proteomes9020030