

A Network-Level Stochastic Model for Pacemaker GABAergic Neurons in Substantia Nigra Pars Reticulata

Abstract
:1. Introduction
Substantia Nigra Pars Reticulata
2. Mathematical Formulation
Model Definition
3. Results
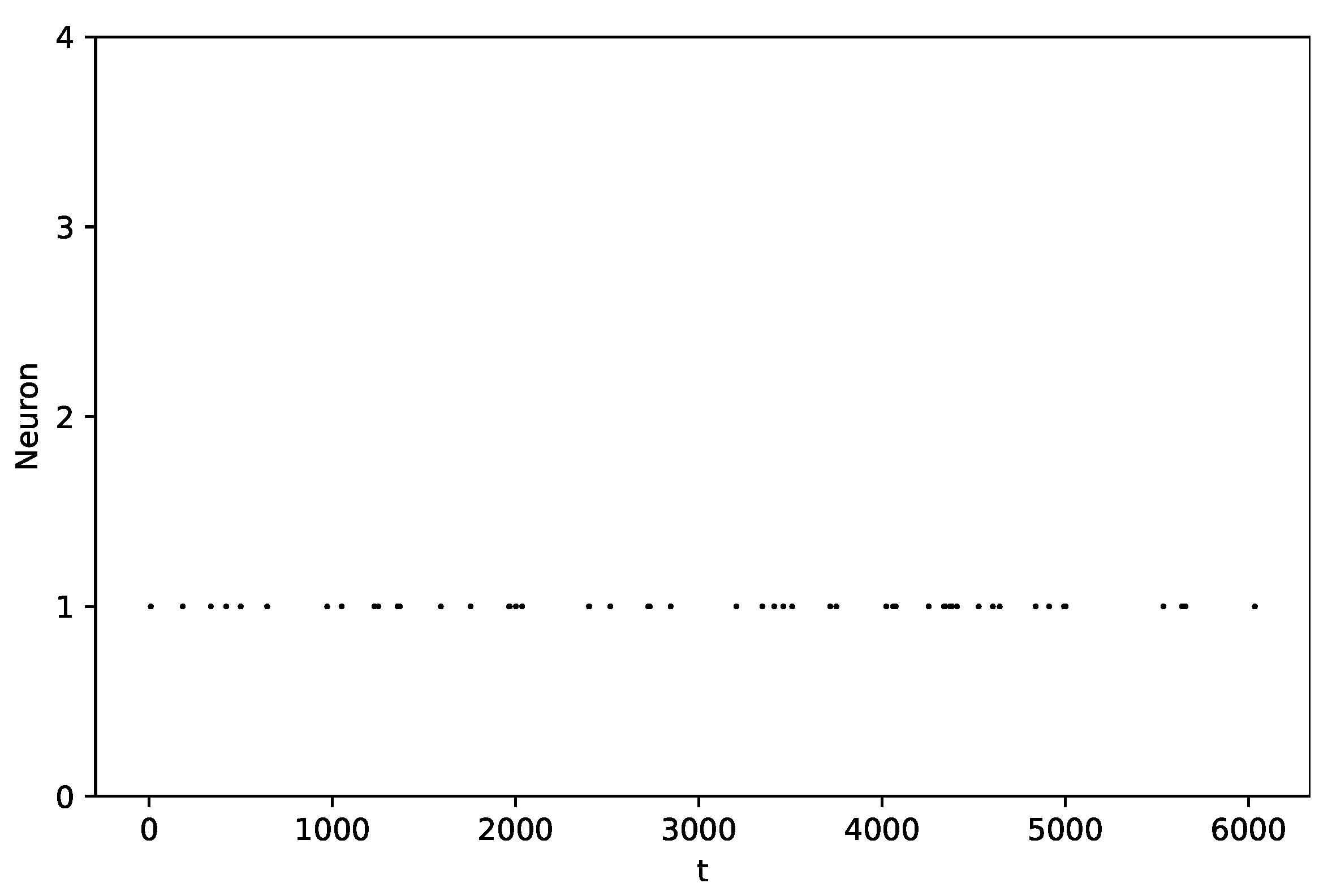
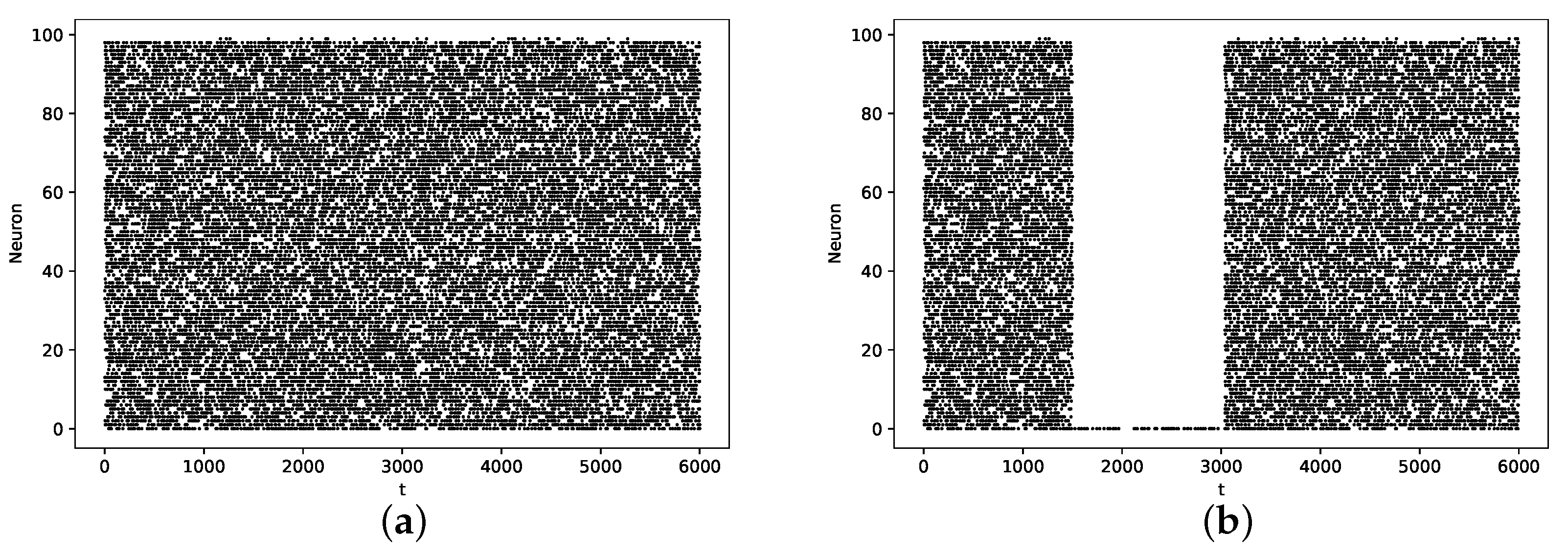
3.1. Baseline Case
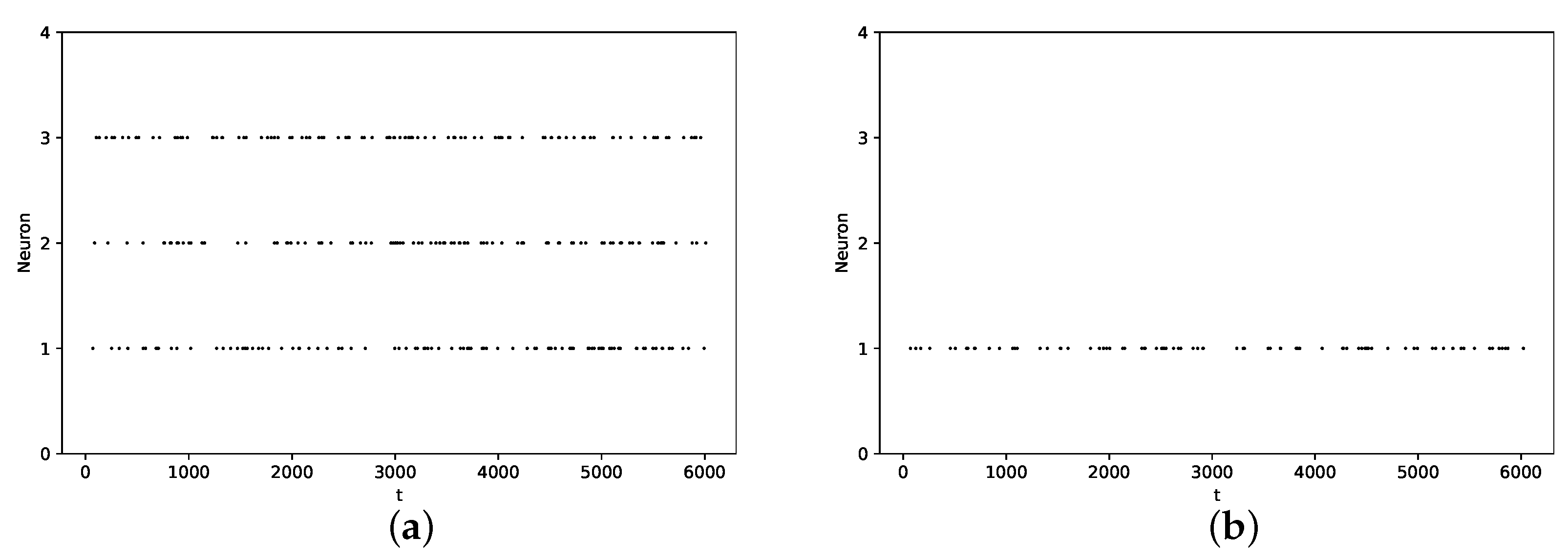
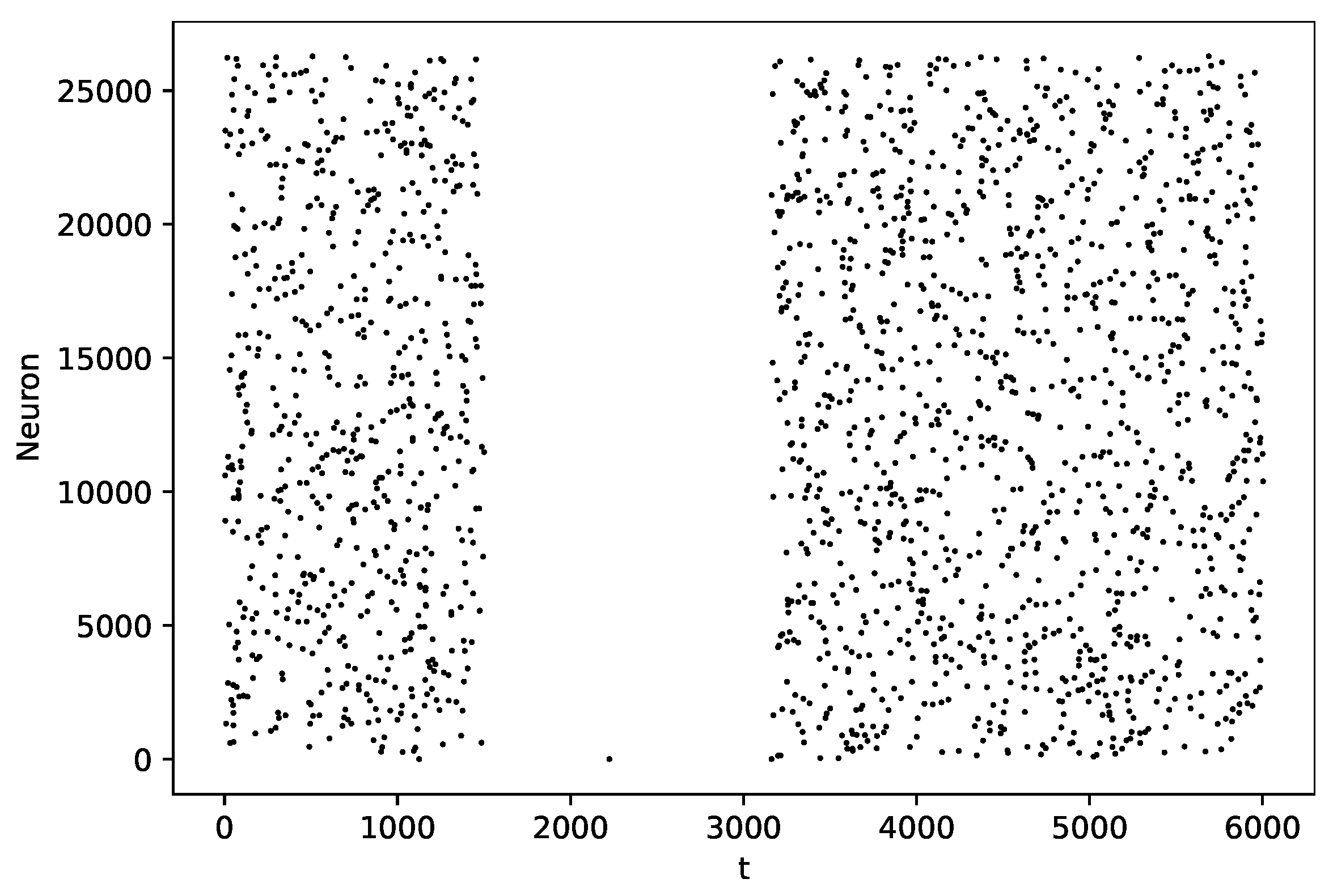
3.2. Pause Phenomenon
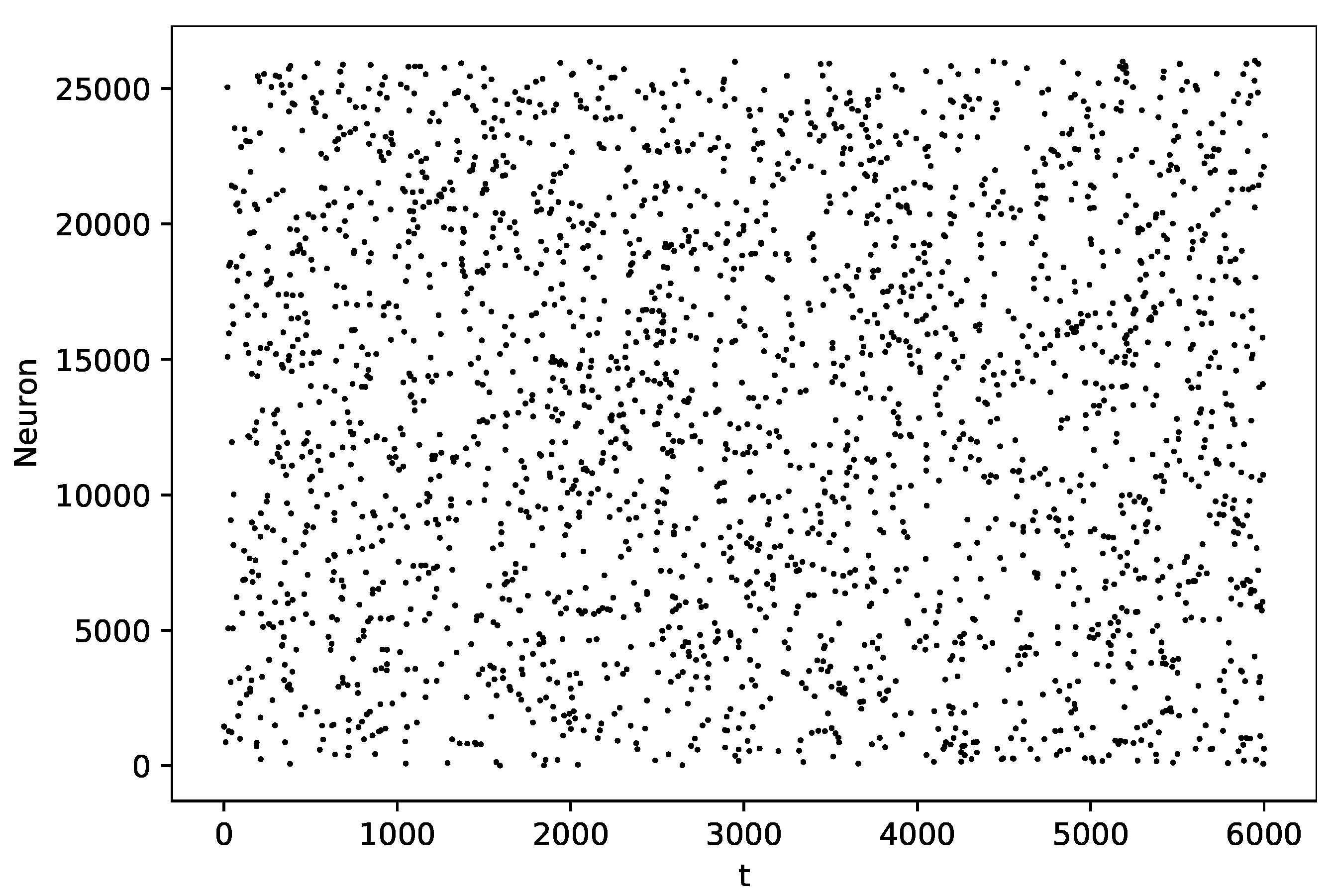
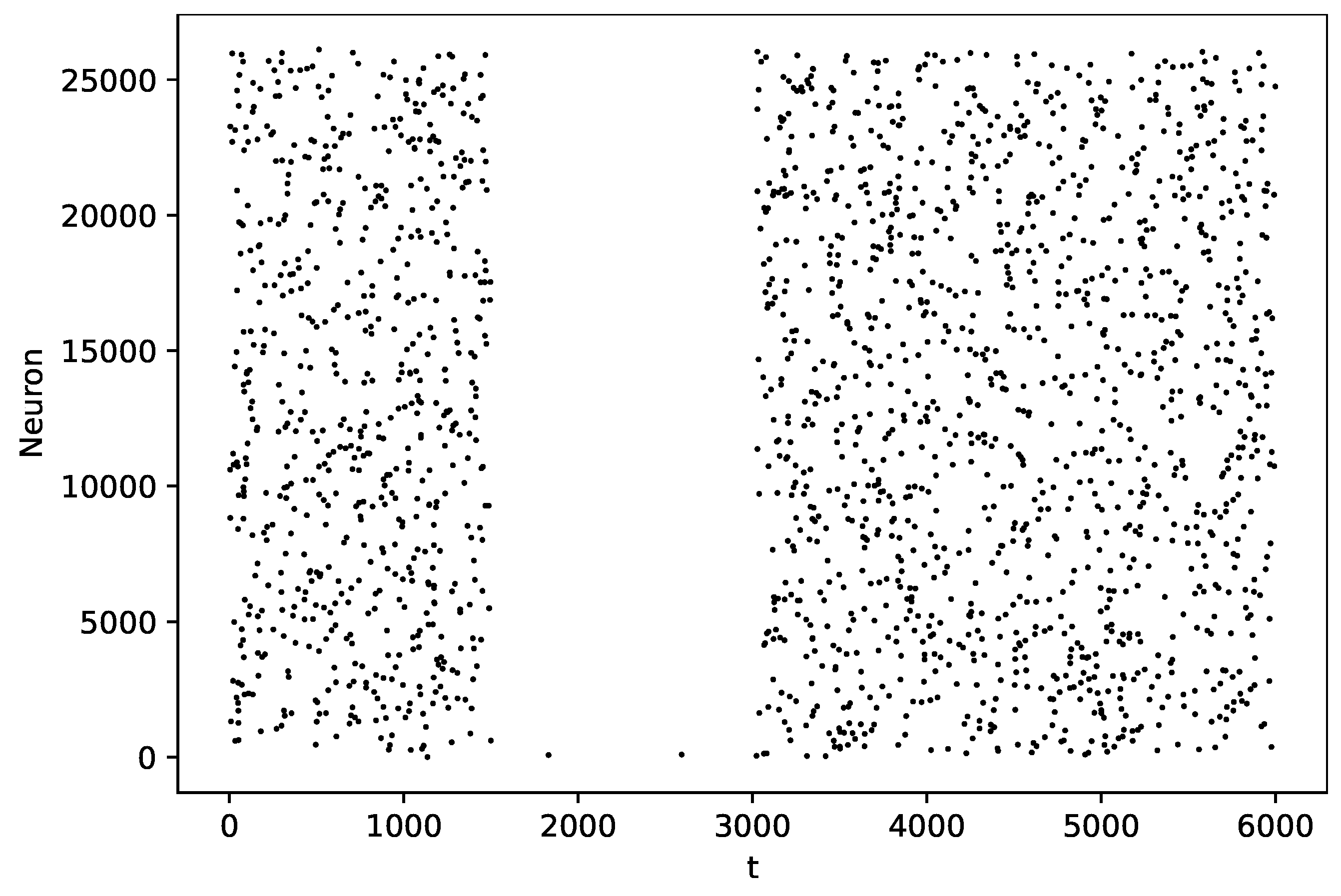
3.3. Random Graph
3.4. Sensitivity Analysis
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Description | Value |
|---|---|---|
| initial condition-uniform random variable | values in [−1, 0] | |
| resting potential | −30 | |
| potential loss to the extracellular medium | 0.020 | |
| w | synaptic weight | −0.9 |
References
- Faisal, A.A.; Laughlin, S.B. Stochastic simulations on the reliability of action potential propagation in this axons. PLoS Comp. Biol. 2007, 3, 783–795. [Google Scholar] [CrossRef] [PubMed]
- Goldwyn, J.H.; Imennov, N.S.; Famulare, M.; Shea-Brown, E. Stochastic differential equation models for ion channel noise in Hodgkin-Huxley neurons. Phys. Rev. Stat. Nonlinear Soft Matter Phys. 2011, 83 Pt 1, 041908. [Google Scholar] [CrossRef] [PubMed]
- Luo, L. Principles of Neurobiology, 1st ed.; Garland Science: New York, NY, USA, 2016. [Google Scholar]
- Mainen, Z.F.; Sejnowski, T.J. Reliability of spike timing in neocortical neurons. Science 1995, 268, 1503–1506. [Google Scholar] [CrossRef] [PubMed]
- Schneidman, E.; Freedman, B.; Segev, I. Ion channel stochasticity may be critical in determining the reliability and precision of spike timing. Neural Comput. 1998, 10, 1679–1703. [Google Scholar] [CrossRef] [PubMed]
- De Masi, A.; Galves, A.; Löcherbach, E.; Presutti, E. Hydrodynamic limit for interacting neurons. J. Stat. Phys. 2015, 158, 866–902. [Google Scholar] [CrossRef]
- Galves, A.; Löcherbach, E. Infinite Systems of Interacting Chains with Memory of Variable Length—A Stochastic Model for Biological Neural Nets. J. Stat. Phys. 2013, 151, 896–921. [Google Scholar] [CrossRef]
- Bolam, J.P.; Hanley, J.J.; Booth, P.A.C.; Bevan, M.D. Synaptic organization of the basal ganglia. J. Anat. 2000, 196, 527–542. [Google Scholar] [CrossRef]
- Deniau, J.M.; Mailly, P.; Maurice, N.; Charpier, S. The pars reticulata of the substantia nigra: A window to basal ganglia output. Prog. Brain Res. 2007, 160, 151–172. [Google Scholar]
- Guatteo, E.; Cucchiaroni, M.L.; Mercuri, N.B. Substantia nigra control of basal ganglia nuclei. J. Neural Transm. Suppl. 2009, 73, 91–101. [Google Scholar]
- Hardman, C.D.; McRitchie, D.A.; Halliday, G.M.; Cartwright, H.R.; Morris, J.G. Substantia nigra pars reticulata neurons in Parkinson’s disease. Neurodegeneration 1996, 5, 49–55. [Google Scholar] [CrossRef]
- Hu, Y.; Ma, T.C.; Alberico, S.L.; Ding, Y.; Jin, L.; Kang, U.J. Substantia Nigra Pars Reticulata Projections to the Pedunculopontine Nucleus Modulate Dyskinesia. Mov. Disord. 2023. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Wichmann, T.; DeLong, M.R. Pathophysiology of Parkinson’s disease: The MPTP primate model of the human disorder. Ann. N. Y. Acad. Sci. 2003, 991, 199–213. [Google Scholar] [CrossRef] [PubMed]
- Graybiel, A.M.; Rauch, S.L. Toward a neurobiology of obsessive-compulsive disorder. Neuron 2000, 28, 343–347. [Google Scholar] [CrossRef] [PubMed]
- Graybiel, A.M.; Canales, J.J. The neurobiology of repetitive behaviors: Clues to the neurobiology of Tourette syndrome. Adv. Neurol. 2001, 85, 123–131. [Google Scholar] [PubMed]
- Parent, A. Extrinsic connections of the basal ganglia. Trends Neurosci. 1990, 13, 254–258. [Google Scholar] [CrossRef]
- Hikosaka, O.; Tepper, J.M.; Abercrombie, E.D.; Bolam, J.P. GABAergic output of the basal ganglia. Prog. Brain Res. 2007, 160, 209–226. [Google Scholar]
- Freeze, B.S.; Kravitz, A.V.; Hammack, N.; Berke, J.D.; Kreitzer, A.C. Control of basal ganglia output by direct and indirect pathway projection neurons. J. Neurosci. 2013, 33, 18531–18539. [Google Scholar] [CrossRef]
- Naito, A.; Kita, H. The cortico-nigral projection in the rat: An anterograde tracing study with biotinylated dextran amine. Brain Res. 1994, 637, 317–322. [Google Scholar] [CrossRef]
- Windels, F.; Kiyatkin, E.A. GABA, not glutamate, controls the activity of substantia nigra reticulata neurons in awake, unrestrained rats. J. Neurosci. Off. J. Soc. Neurosci. 2004, 24, 6751–6754. [Google Scholar] [CrossRef]
- Deniau, J.M.; Kitai, S.T.; Donoghue, J.P.; Grofova, I. Neuronal interactions in the substantia nigra pars reticulata through axon collaterals of the projection neurons. Exp. Brain Res. 1982, 47, 105–113. [Google Scholar] [CrossRef]
- Mailly, P.; Charpier, S.; Menetrey, A.; Deniau, J.-M. Three-Dimensional Organization of the Recurrent Axon Collateral Network of the Substantia Nigra Pars Reticulata Neurons in the Rat. J. Neurosci. 2003, 23, 5247. [Google Scholar] [CrossRef] [PubMed]
- Surmeier, D.J.; Mercer, J.N.; Chan, S. Autonomous pacemakers in the basal ganglia: Who needs excitatory synapses anyway? Curr. Opin. Neurobiol. 2005, 15, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Seutin, V.; Engel, D. Differences in Na+ Conductance Density and Na+ Channel Functional Properties Between Dopamine and GABA Neurons of the Rat Substantia Nigra. J. Neurophysiol. 2010, 103, 3099–3114. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Zhou, F.-M. Molecular and functional differences in voltage-activated sodium currents between GABA projection neurons and dopamine neurons in the substantia nigra. Soc. Neurosci. Abstr. 2010, 339, 22. [Google Scholar] [CrossRef]
- Ding, S.; Wei, W.; Zhou, F.-M. Kv3-like potassium channels are required for sustained high-frequency firing in basal ganglia output neurons. J. Neurophysiol. 2011, 106, 3019–3034. [Google Scholar] [CrossRef]
- Zhou, F.M.; Lee, C.R. Intrinsic and integrative properties of substantia nigra pars reticulata neurons. Neuroscience 2011, 198, 69–94. [Google Scholar] [CrossRef]
- Higgs, M.H.; Wilson, C.J. Unitary synaptic connections among substantia nigra pars reticulata neurons. J. Neurophysiol. 2016, 115, 2814–2829. [Google Scholar] [CrossRef]
- Simmons, D.V.; Higgs, M.H.; Lebby, S.; Wilson, C.J. Predicting responses to inhibitory synaptic input in substantia nigra pars reticulate neurons. J. Neurophysiol. 2018, 120, 2679–2693. [Google Scholar] [CrossRef]
- Lanciego, J.L.; Luquin, N.; Obeso, J.A. Functional neuroanatomy of the basal ganglia. Cold Spring Harb. Perspect. Med. 2012, 2, a009621. [Google Scholar] [CrossRef]
- Wilson, C.J. Active decorrelation in the basal ganglia. Neuroscience 2013, 250, 467–482. [Google Scholar] [CrossRef]
- Masuda, N.; Rocha, L.E.C. A Gillespie algorithm for non-Markovian stochastic processes. Siam Rev. 2018, 60, 95–115. [Google Scholar] [CrossRef]
- Richards, C.D.; Shiroyama, T.; Kitai, S.T. Electrophysiological and immunocytochemical characterization of GABA and Dopamine neurons in the Substantia Nigra of the rat. Neuroscience 1997, 80, 545–577. [Google Scholar] [CrossRef] [PubMed]
- Brunel, N.; Hakim, V. Fast global oscillations in networks of integrate-and-fire neurons with low firing rates. Neural Comput. 1999, 11, 1621–1671. [Google Scholar] [CrossRef] [PubMed]
- Brunel, N. Dynamics of Sparsely Connected Networks of Excitatory and Inhibitory Spiking Neurons. J. Comput. Neurosci. 2000, 8, 183–208. [Google Scholar] [CrossRef]
- Saltelli, A.; Tarantola, S.; Campolongo, F. Sensitivity Analysis as an Ingredient of Modeling. Stat. Sci. 2000, 15, 377–395. [Google Scholar]
- Phillips, R.S.; Rosner, I.; Gittis, A.H.; Rubin, J.E. A computational model explains and predicts substantia nigra pars reticulata responses to pallidal and striatal inputs. bioRxiv 2020. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guimarães, K.; Duarte, A. A Network-Level Stochastic Model for Pacemaker GABAergic Neurons in Substantia Nigra Pars Reticulata. Mathematics 2023, 11, 3778. https://doi.org/10.3390/math11173778
Guimarães K, Duarte A. A Network-Level Stochastic Model for Pacemaker GABAergic Neurons in Substantia Nigra Pars Reticulata. Mathematics. 2023; 11(17):3778. https://doi.org/10.3390/math11173778
Chicago/Turabian StyleGuimarães, Karine, and Aline Duarte. 2023. "A Network-Level Stochastic Model for Pacemaker GABAergic Neurons in Substantia Nigra Pars Reticulata" Mathematics 11, no. 17: 3778. https://doi.org/10.3390/math11173778
APA StyleGuimarães, K., & Duarte, A. (2023). A Network-Level Stochastic Model for Pacemaker GABAergic Neurons in Substantia Nigra Pars Reticulata. Mathematics, 11(17), 3778. https://doi.org/10.3390/math11173778