
Neimark–Sacker Bifurcation of a Discrete-Time Predator–Prey Model with Prey Refuge Effect



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Preliminaries and Notation
3. Existence and Stability of Equilibria
- (i)
- The trivial equilibrium is a saddle point if and only if , and it is a source if and only if .
- (ii)
- The boundary equilibrium is a sink if and only if , it is a saddle point if and only if , and transcritical bifurcation occurs at if and only if .
- (i)
- First, the Jacobian matrix of (2) at point is given by:Then, eigenvalues of are and . Therefore, is a saddle point if and only if , and it is a source if and only if .
- (ii)
- Secondly, the Jacobian matrix of (2) at point is given by:Then, eigenvalues of are and . Therefore, is a sink if and only if , it is a saddle point if and only if , and transcritical bifurcation occurs at if and only if .
- (i)
- The positive equilibrium is a sink if
- (ii)
- The positive equilibrium is a saddle point if
- (iii)
- The positive equilibrium is a source if
- (iv)
- Transcritical bifurcation occurs at if
- (v)
- Flip bifurcation occurs at if
- (vi)
- Neimark–Sacker bifurcation occurs at if
- (i)
- is a sink with the eigenvalues and , which is equivalent to , , and .
- (ii)
- is a saddle point with the eigenvalues and ( and ), which is equivalent to and .
- (iii)
- is a source with the eigenvalues and , which is equivalent to , , and .
- (iv)
- Transcritical bifurcation occurs with the eigenvalues , or , , which is equivalent to and .
- (v)
- Flip bifurcation occurs with the eigenvalues , or , , which is equivalent to and .
- (vi)
- Neimark–Sacker bifurcation occurs with the eigenvalues and , which is equivalent to and .
4. Neimark–Sacker Bifurcation at
4.1. Existence Condition of Neimark–Sacker Bifurcation at
4.2. The Direction of Neimark–Sacker Bifurcation at
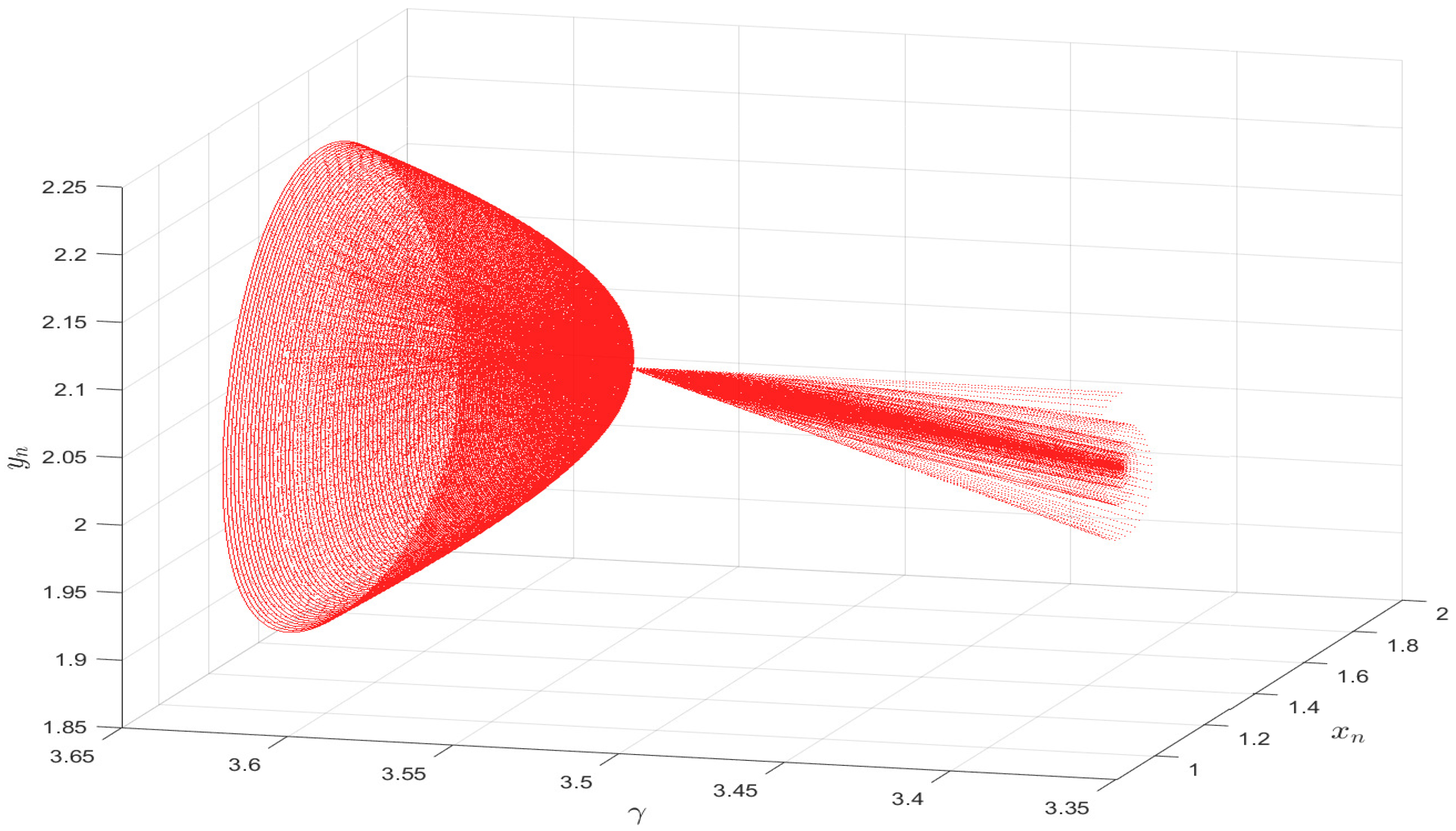
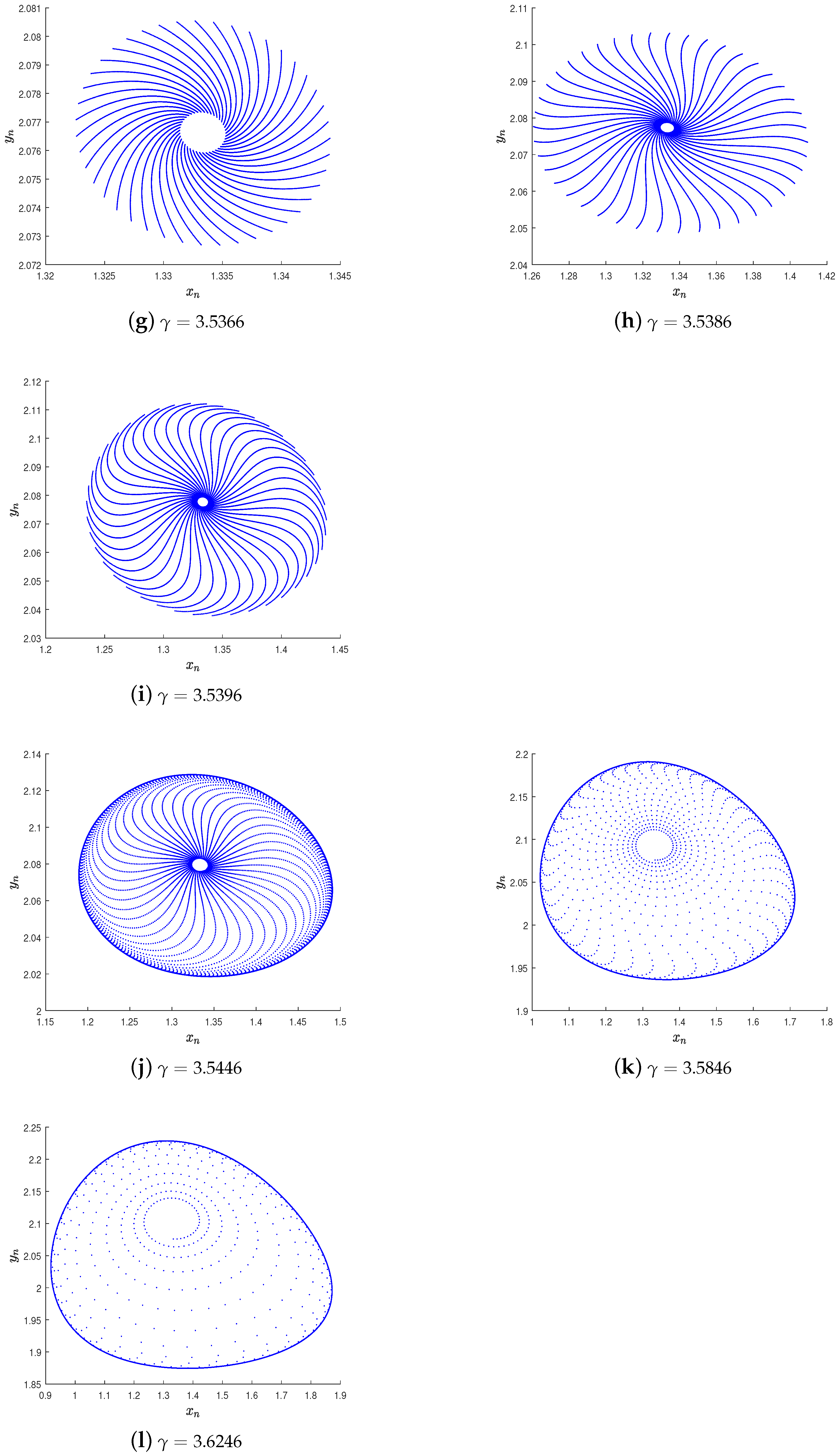
5. Numerical Simulation
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lotka, A.J. Analytical Note on Certain Rhythmic Relations in Organic Systems. Proc. Natl. Acad. Sci. USA 1920, 6, 410–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volterra, V. Variazioni e Fluttuazioni del Numero d’Individui in Specie Animali Conviventi. Mem. R. Accad. Naz. Lincei. Ser. VI 1926, 2, 31–113. [Google Scholar]
- Lotka, A.J. Contribution to the Mathematical Theory of Capture. Proc. Natl. Acad. Sci. USA 1932, 18, 172–178. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Zhang, Q.L.; Huang, J.; Tang, W.S. Dynamical Behavior of a Harvested Prey-Predator Model with Stage Structure and Discrete Time Delay. J. Biol. Syst. 2009, 17, 759–777. [Google Scholar] [CrossRef]
- Samuelson, P.A. Generalized Predator-Prey Oscillations in Ecological and Economic Equilibrium. Proc. Natl. Acad. Sci. USA 1971, 68, 980–983. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.X.; Zhang, C.R. Stability and Optimal Control of Tree-Insect Model under Forest Fire Disturbance. Mathematics 2022, 10, 2563. [Google Scholar] [CrossRef]
- Chen, H.Y.; Zhang, C.R. Dynamic analysis of a Leslie–Gower-type predator–prey system with the fear effect and ratio-dependent Holling III functional response. Nonlinear Anal. Model. Control. 2022, 27, 1–23. [Google Scholar] [CrossRef]
- Liu, X.L.; Xiao, D.M. Complex dynamic behaviors of a discrete-time predator–prey system. Chaos Solitons Fractals 2007, 32, 80–94. [Google Scholar] [CrossRef]
- Beretta, E.; Capasso, V.; Rinaldi, F. Global stability results for a generalized Lotka-Volterra system with distributed delays. J. Math. Biol. 1988, 26, 661–688. [Google Scholar] [CrossRef]
- Hofbauer, J.; Thus, J.W. Multiple limit cycles for predator-prey models. Math. Biosci. 1990, 99, 71–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brauer, F.; Sánchez, D.A. Constant rate population harvesting: Equilibrium and stability. Theor. Popul. Biol. 1975, 8, 12–30. [Google Scholar] [CrossRef]
- Cheng, L.F.; Cao, H.J. Bifurcation analysis of a discrete-time ratio-dependent predator–prey model with Allee Effect. Commun. Nonlinear Sci. Numer. Simul. 2016, 38, 288–302. [Google Scholar] [CrossRef]
- Lai, L.Y.; Zhu, Z.L.; Chen, F.D. Stability and Bifurcation in a Predator-Prey Model with the Additive Allee Effect and the Fear Effect dagger. Mathematics 2020, 8, 1280. [Google Scholar] [CrossRef]
- Sasmal, S.K. Population dynamics with multiple allee effects induced by fear factors—A mathematical study on prey-predator interactions. Appl. Math. Model. 2018, 64, 1–14. [Google Scholar] [CrossRef]
- Kuznetsov, Y.A.; Rinaldi, S. Remarks on food chain dynamics. Math. Biosci. 1996, 134, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Aziz-Alaoui, M.A.; Okiye, M.D. Boundedness and global stability for a predator-prey model with modi-fied Leslie-Gower and Holling-type II schemes. Appl. Math. Lett. 2003, 16, 1069–1075. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.D.; Chen, L.J.; Xie, X.D. On a Leslie-Gower predator-prey model incorporating a prey refuge. Nonlinear Anal. Real World Appl. 2009, 10, 2905–2908. [Google Scholar] [CrossRef]
- Hung, K.-C.; Wang, S.-H. A theorem on S-shaped bifurcation curve for a positone problem with convex–concave nonlinearity and its applications to the perturbed Gelfand problem. J. Differ. Equ. 2011, 251, 223–237. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.Y.; Teng, Z.D.; Zhang, L. Stability and bifurcation analysis of a discrete predator–prey model with nonmonotonic functional response. Nonlinear Anal. Real World Appl. 2011, 12, 2356–2377. [Google Scholar] [CrossRef]
- Zhang, C.R.; Zheng, B. Stability and bifurcation of a two-dimension discrete neural network model with multi-delays. Chaos Solitons Fractals 2007, 31, 1232–1242. [Google Scholar] [CrossRef]
- Abdelaziz, M.A.M.; Ismail, A.I.; Abdullah, F.A.; Mohd, M.H. Bifurcations and chaos in a discrete SI epi-demic model with fractional order. Adv. Differ. Equ. 2018, 2018, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, J.; Sahoo, B.; Poria, S. Prey-predator dynamics with prey refuge providing additional food to predator. Chaos Solitons Fractals 2017, 96, 110–119. [Google Scholar] [CrossRef]
- Gladkov, S.O. On the Question of Self-Organization of Population Dynamics on Earth. Biophysics 2021, 66, 858–866. [Google Scholar] [CrossRef]
- Li, W.; Li, X.Y. Neimark–Sacker Bifurcation of a Semi-Discrete Hematopoiesis Model. J. Appl. Anal. Comput. 2018, 8, 1679–1693. [Google Scholar] [CrossRef]
- Li, X.Y.; Shao, X.M. Flip bifurcation and Neimark–Sacker bifurcation in a discrete predator-prey model with Michaelis-Menten functional response. Electron. Res. Arch. 2023, 31, 37–57. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, B.; Zhang, C. Neimark–Sacker Bifurcation of a Discrete-Time Predator–Prey Model with Prey Refuge Effect. Mathematics 2023, 11, 1399. https://doi.org/10.3390/math11061399
Hong B, Zhang C. Neimark–Sacker Bifurcation of a Discrete-Time Predator–Prey Model with Prey Refuge Effect. Mathematics. 2023; 11(6):1399. https://doi.org/10.3390/math11061399
Chicago/Turabian StyleHong, Binhao, and Chunrui Zhang. 2023. "Neimark–Sacker Bifurcation of a Discrete-Time Predator–Prey Model with Prey Refuge Effect" Mathematics 11, no. 6: 1399. https://doi.org/10.3390/math11061399
APA StyleHong, B., & Zhang, C. (2023). Neimark–Sacker Bifurcation of a Discrete-Time Predator–Prey Model with Prey Refuge Effect. Mathematics, 11(6), 1399. https://doi.org/10.3390/math11061399