
Bursting Dynamics of Spiking Neural Network Induced by Active Extracellular Medium



{kind=link}
{kind=link}
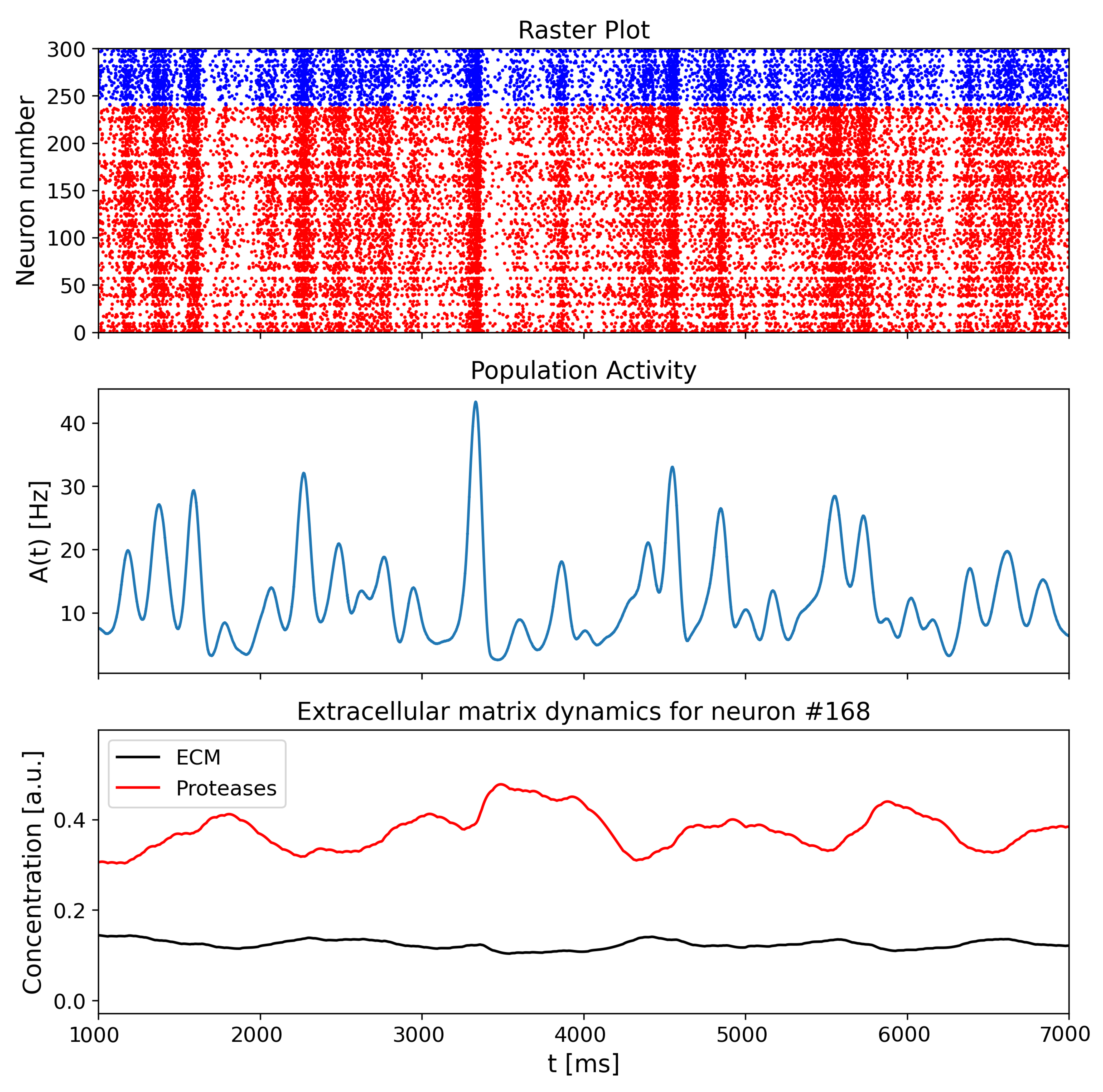
{kind=link}
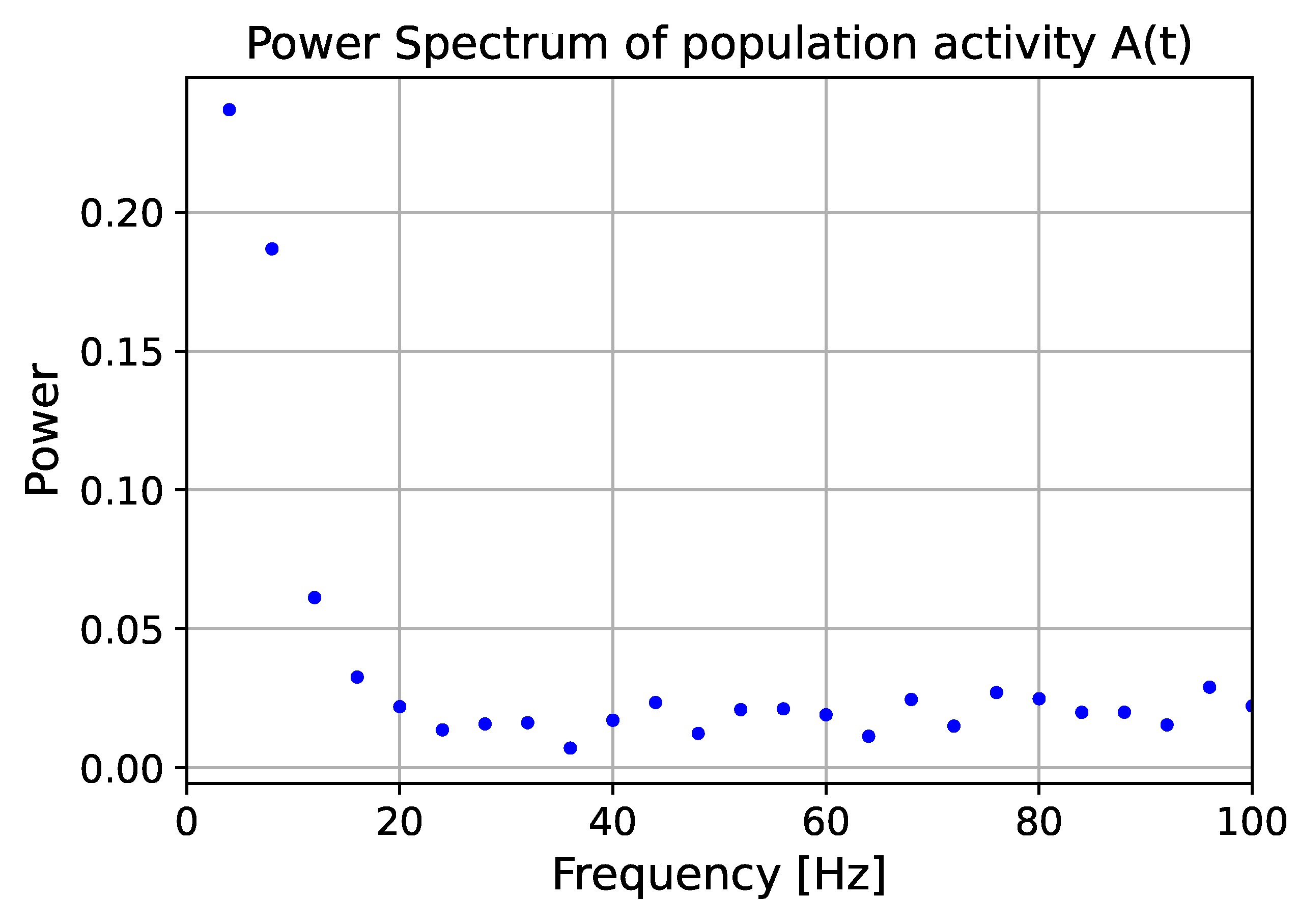
{kind=link}
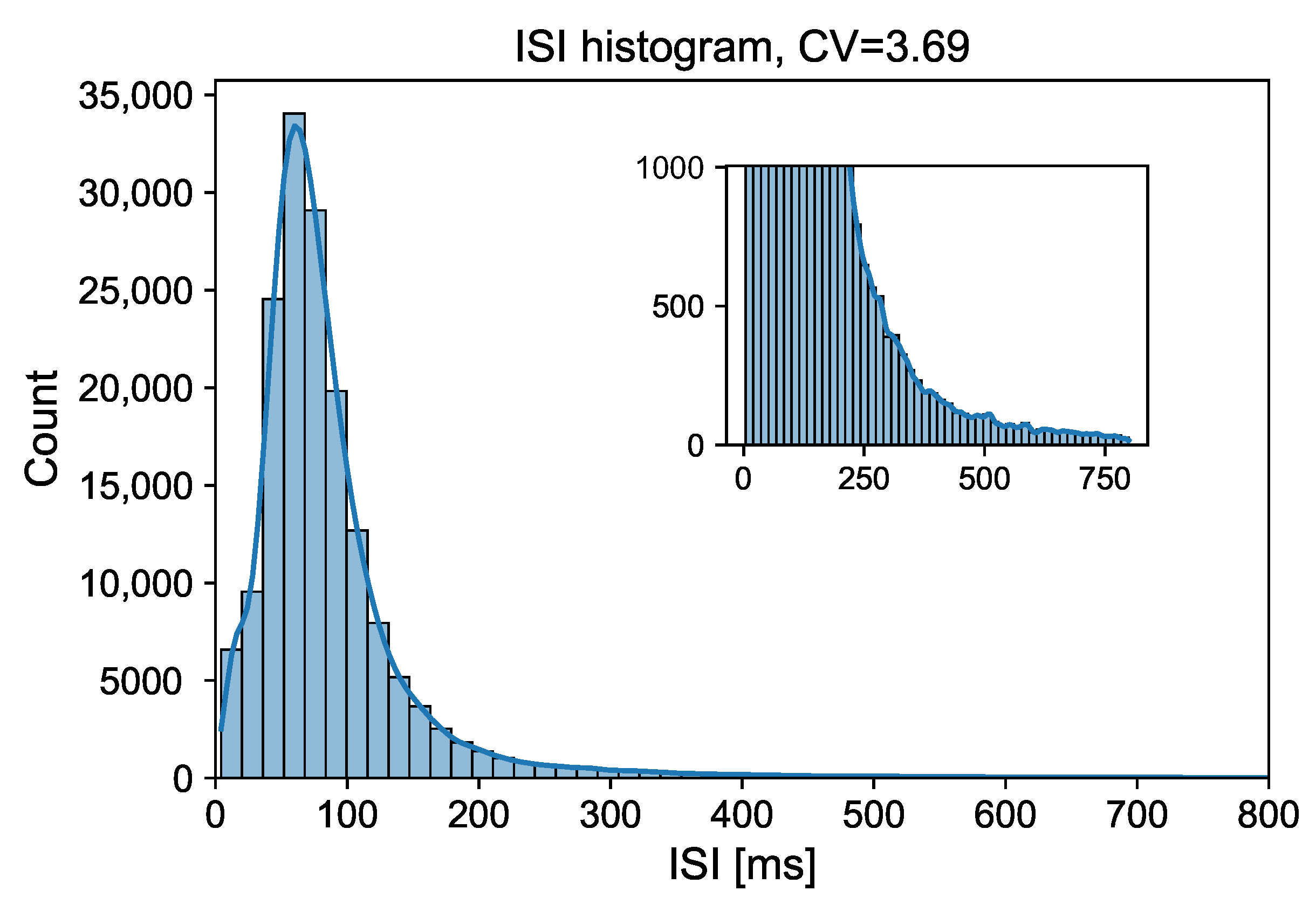
{kind=link}
{kind=link}
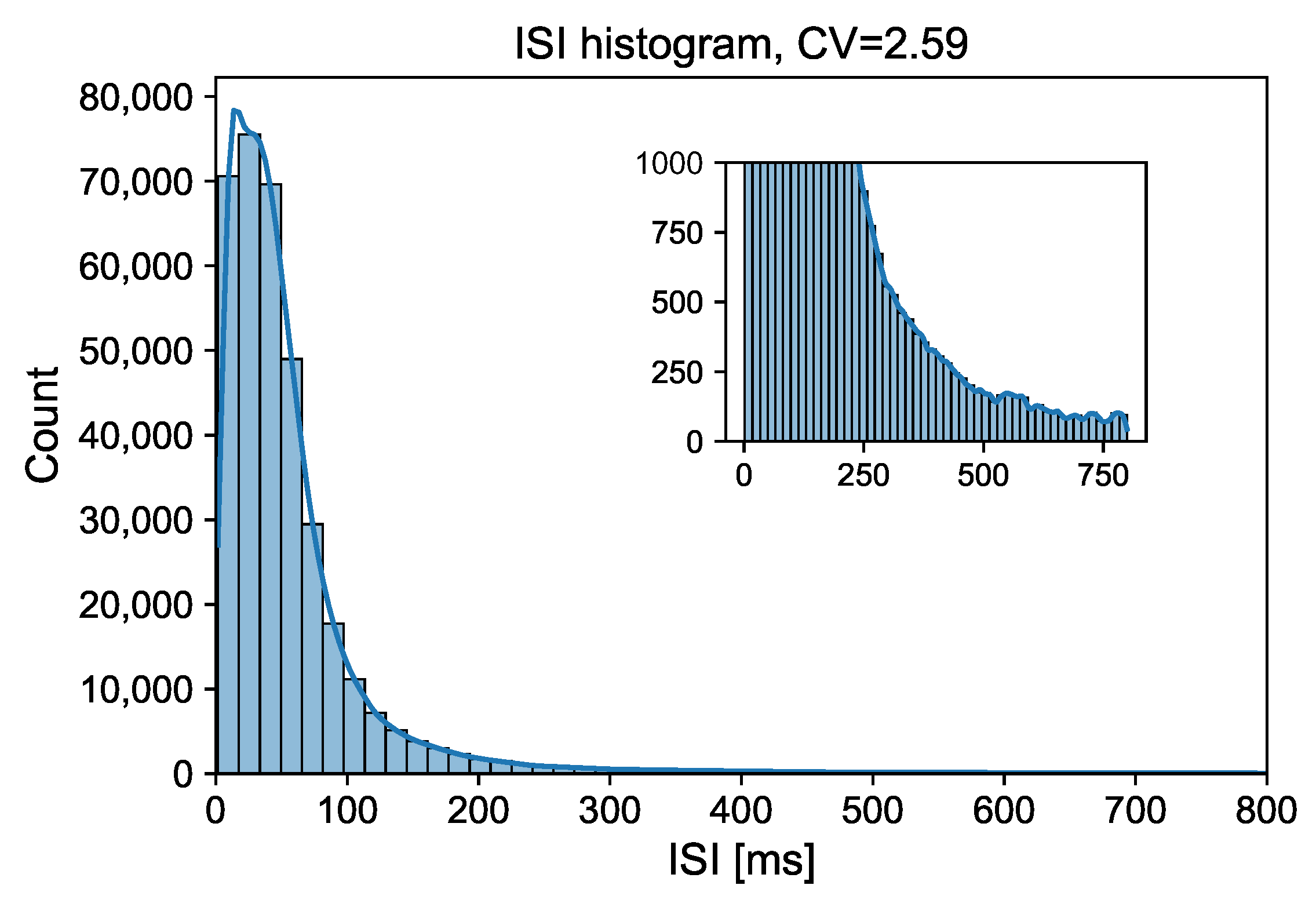{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Model
2.1. Average Activity Level
2.2. Extracellular Matrix Dynamics
2.3. Extracellular Matrix Modulation of Neural Activity
2.4. Neural Network
2.5. Numerical Simulation and Data Analysis Methods
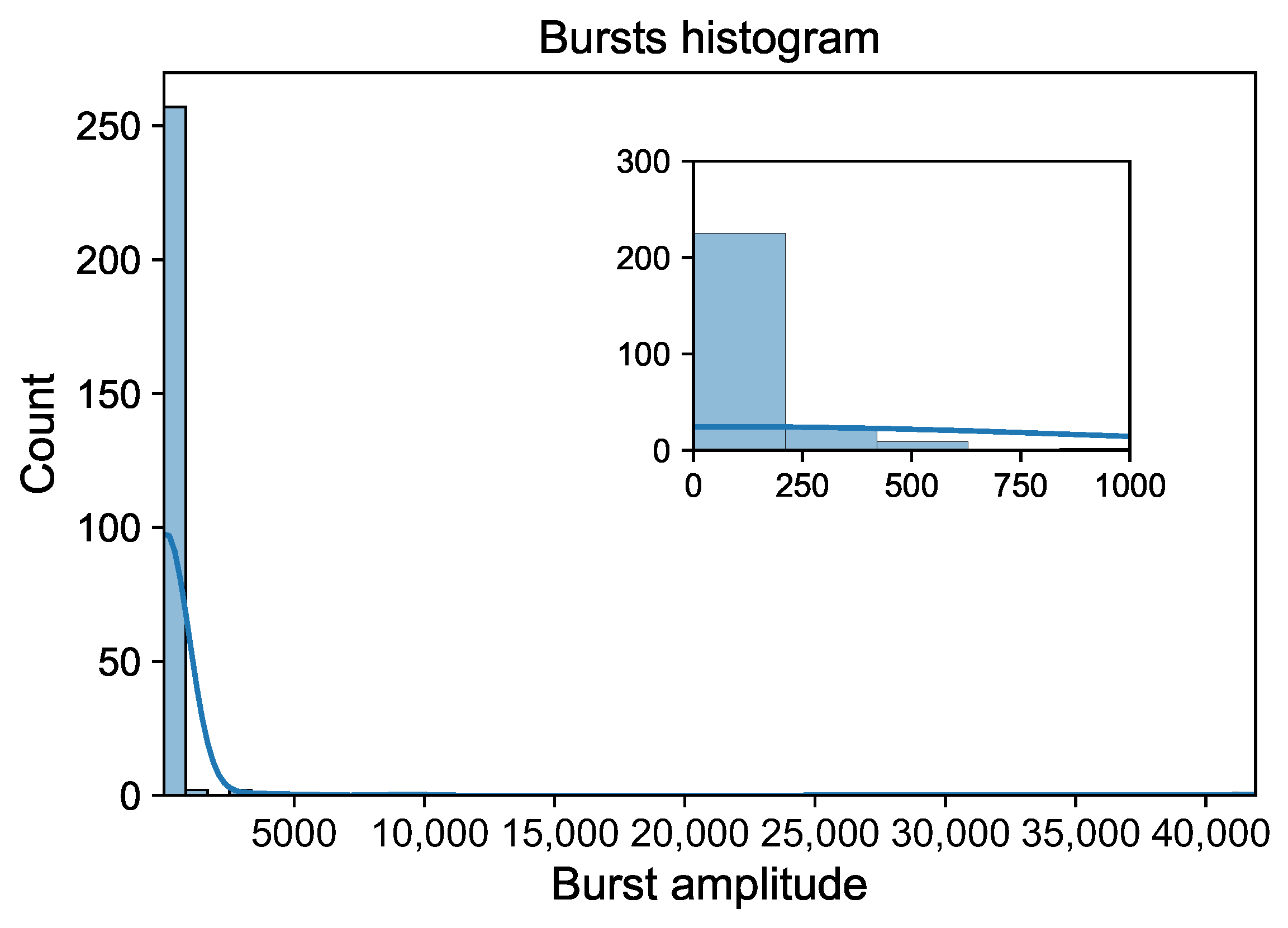
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fries, P. A mechanism for cognitive dynamics: Neuronal communication through neuronal coherence. Trends Cogn. Sci. 2005, 9, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Timofeev, I.; Bazhenov, M.; Seigneur, J.; Sejnowski, T. Neuronal synchronization and thalamocortical rhythms in sleep, wake and epilepsy. In Jasper’s Basic Mechanisms of the Epilepsies, 4th ed.; National Center for Biotechnology Information: Bethesda, MD, USA, 2012. [Google Scholar]
- Fell, J.; Axmacher, N. The role of phase synchronization in memory processes. Nat. Rev. Neurosci. 2011, 12, 105–118. [Google Scholar] [CrossRef]
- Baldauf, D.; Desimone, R. Neural mechanisms of object-based attention. Science 2014, 344, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Fries, P.; Reynolds, J.; Rorie, A.; Desimone, R. Modulation of oscillatory neuronal synchronization by selective visual attention. Science 2001, 291, 1560–1563. [Google Scholar] [CrossRef] [PubMed]
- Schnitzler, A.; Gross, J. Normal and pathological oscillatory communication in the brain. Nat. Rev. Neurosci. 2005, 6, 285–296. [Google Scholar] [CrossRef]
- Velazquez, J.; Cortez, M.; Snead, O., III; Wennberg, R. Dynamical regimes underlying epileptiform events: Role of instabilities and bifurcations in brain activity. Phys. D Nonlinear Phenom. 2003, 186, 205–220. [Google Scholar] [CrossRef]
- Wagenaar, D.; Pine, J.; Potter, S. An extremely rich repertoire of bursting patterns during the development of cortical cultures. BMC Neurosci. 2006, 7, 1–18. [Google Scholar] [CrossRef]
- Wagenaar, D.; Madhavan, R.; Pine, J.; Potter, S. Controlling bursting in cortical cultures with closed-loop multi-electrode stimulation. J. Neurosci. 2005, 25, 680–688. [Google Scholar] [CrossRef]
- Wagenaar, D.; Nadasdy, Z.; Potter, S. Persistent dynamic attractors in activity patterns of cultured neuronal networks. Phys. Rev. E 2006, 73, 051907. [Google Scholar] [CrossRef]
- Pasquale, V.; Massobrio, P.; Bologna, L.; Chiappalone, M.; Martinoia, S. Self-organization and neuronal avalanches in networks of dissociated cortical neurons. Neuroscience 2008, 153, 1354–1369. [Google Scholar] [CrossRef]
- Zeldenrust, F.; Wadman, W.; Englitz, B. Neural coding with bursts—Current state and future perspectives. Front. Comput. Neurosci. 2018, 12, 48. [Google Scholar] [CrossRef] [PubMed]
- Pimashkin, A.; Kastalskiy, I.; Simonov, A.; Koryagina, E.; Mukhina, I.; Kazantsev, V. Spiking signatures of spontaneous activity bursts in hippocampal cultures. Front. Comput. Neurosci. 2011, 5, 46. [Google Scholar] [CrossRef] [PubMed]
- Wang, X. Neurophysiological and computational principles of cortical rhythms in cognition. Physiol. Rev. 2010, 90, 1195–1268. [Google Scholar] [CrossRef] [PubMed]
- Zeitler, M.; Daffertshofer, A.; Gielen, C. Asymmetry in pulse-coupled oscillators with delay. Phys. Rev. E 2009, 79, 065203. [Google Scholar] [CrossRef] [PubMed]
- Pikovsky, A.; Rosenblum, M.; Kurths, J. Synchronization: A Universal Concept in Nonlinear Science; American Association of Physics Teachers; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Tsybina, Y.; Kastalskiy, I.; Kazantsev, V.; Gordleeva, S. Synchronization events in a spiking neural network. In Proceedings of the 2022 Fourth International Conference Neurotechnologies and Neurointerfaces (CNN), Kaliningrad, Russia, 14–16 September 2022; pp. 206–208. [Google Scholar]
- Simonov, A.; Gordleeva, S. Synchronization with an arbitrary phase shift in a pair of synaptically coupled neural oscillators. JETP Lett. 2014, 98, 632–637. [Google Scholar] [CrossRef]
- Muthukumaraswamy, S.; Edden, R.; Jones, D.; Swettenham, J.; Singh, K. Resting GABA concentration predicts peak gamma frequency and fMRI amplitude in response to visual stimulation in humans. Proc. Natl. Acad. Sci. USA 2009, 106, 8356–8361. [Google Scholar] [CrossRef]
- Barabash, N.; Levanova, T.; Stasenko, S. Rhythmogenesis in the mean field model of the neuron–glial network. Eur. Phys. J. Spec. Top. 2023. [Google Scholar] [CrossRef]
- Stasenko, S.; Kazantsev, V. 3D model of bursting activity generation. In Proceedings of the 2022 Fourth International Conference Neurotechnologies and Neurointerfaces (CNN), Kaliningrad, Russia, 14–16 September 2022; pp. 176–179. [Google Scholar]
- Stasenko, S.; Lazarevich, I.; Kazantsev, V. Quasi-synchronous neuronal activity of the network induced by astrocytes. Procedia Comput. Sci. 2020, 169, 704–709. [Google Scholar] [CrossRef]
- Pankratova, E.; Kalyakulina, A.; Stasenko, S.; Gordleeva, S.; Lazarevich, I.; Kazantsev, V. Neuronal synchronization enhanced by neuron–astrocyte interaction. Nonlinear Dyn. 2019, 97, 647–662. [Google Scholar] [CrossRef]
- Lazarevich, I.; Stasenko, S.; Kazantsev, V. Synaptic multistability and network synchronization induced by the neuron–glial interaction in the brain. JETP Lett. 2017, 105, 210–213. [Google Scholar] [CrossRef]
- Makovkin, S.; Kozinov, E.; Ivanchenko, M.; Gordleeva, S. Controlling synchronization of gamma oscillations by astrocytic modulation in a model hippocampal neural network. Sci. Rep. 2022, 12, 6970. [Google Scholar] [CrossRef]
- Stasenko, S.; Hramov, A.; Kazantsev, V. Loss of neuron network coherence induced by virus-infected astrocytes: A model study. Sci. Rep. 2023, 13, 6401. [Google Scholar] [CrossRef]
- Stasenko, S.; Kazantsev, V. Dynamic Image Representation in a Spiking Neural Network Supplied by Astrocytes. Mathematics 2023, 11, 561. [Google Scholar] [CrossRef]
- Dityatev, A.; Rusakov, D. Molecular signals of plasticity at the tetrapartite synapse. Curr. Opin. Neurobiol. 2011, 21, 353–359. [Google Scholar] [CrossRef]
- Kazantsev, V.; Gordleeva, S.; Stasenko, S.; Dityatev, A. A homeostatic model of neuronal firing governed by feedback signals from the extracellular matrix. PLoS ONE 2012, 7, e41646. [Google Scholar] [CrossRef]
- Rich, M.; Wenner, P. Sensing and expressing homeostatic synaptic plasticity. Trends Neurosci. 2007, 30, 119–125. [Google Scholar] [CrossRef]
- Turrigiano, G. Homeostatic signaling: The positive side of negative feedback. Curr. Opin. Neurobiol. 2007, 17, 318–324. [Google Scholar] [CrossRef]
- Kochlamazashvili, G.; Henneberger, C.; Bukalo, O.; Dvoretskova, E.; Senkov, O.; Lievens, P.; Westenbroek, R.; Engel, A.; Catterall, W.; Rusakov, D.; et al. The extracellular matrix molecule hyaluronic acid regulates hippocampal synaptic plasticity by modulating postsynaptic L-type Ca2+ channels. Neuron 2010, 67, 116–128. [Google Scholar] [CrossRef]
- Hodgkin, A.; Huxley, A. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 1952, 117, 500. [Google Scholar] [CrossRef]
- Fawcett, J.; Fyhn, M.; Jendelova, P.; Kwok, J.; Ruzicka, J.; Sorg, B. The extracellular matrix and perineuronal nets in memory. Mol. Psychiatry 2022, 27, 3192–3203. [Google Scholar] [CrossRef]
- Dityatev, A. Remodeling of extracellular matrix and epileptogenesis. Epilepsia 2010, 51, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Dityatev, A.; Fellin, T. Extracellular matrix in plasticity and epileptogenesis. Neuron Glia Biol. 2008, 4, 235–247. [Google Scholar] [CrossRef]
- Jong, J.; Broekaart, D.; Bongaarts, A.; Mühlebner, A.; Mills, J.; Vliet, E.; Aronica, E. Altered Extracellular Matrix as an Alternative Risk Factor for Epileptogenicity in Brain Tumors. Biomedicines 2022, 10, 2475. [Google Scholar] [CrossRef] [PubMed]
- Kepecs, A.; Lisman, J. Information encoding and computation with spikes and bursts. Netw. Comput. Neural Syst. 2003, 14, 103. [Google Scholar] [CrossRef]
- Lobov, S.; Zharinov, A.; Makarov, V.; Kazantsev, V. Spatial memory in a spiking neural network with robot embodiment. Sensors 2021, 21, 2678. [Google Scholar] [CrossRef]
- Kim, J.; Lee, H.; Choi, W.; Lee, K. Encoding information into autonomously bursting neural network with pairs of time-delayed pulses. Sci. Rep. 2019, 9, 1394. [Google Scholar] [CrossRef]
- Lundqvist, M.; Rose, J.; Herman, P.; Brincat, S.; Buschman, T.; Miller, E. Gamma and beta bursts underlie working memory. Neuron 2016, 90, 152–164. [Google Scholar] [CrossRef]
- Sokolov, I.; Azieva, A.; Burtsev, M. Patterns of spiking activity of neuronal networks in vitro as memory traces. In Biologically Inspired Cognitive Architectures (BICA) For Young Scientists, Proceedings of the First International Early Research Career Enhancement School (FIERCES 2016), Moscow, Russia, 21–24 April 2016; Springer: Cham, Switzerland, 2016; pp. 241–247. [Google Scholar]
- Prince, D. Neurophysiology of epilepsy. Annu. Rev. Neurosci. 1978, 1, 395–415. [Google Scholar] [CrossRef]
- Bikbaev, A.; Frischknecht, R.; Heine, M. Brain extracellular matrix retains connectivity in neuronal networks. Sci. Rep. 2015, 5, 14527. [Google Scholar] [CrossRef]
- Lam, D.; Enright, H.; Cadena, J.; Peters, S.; Sales, A.; Osburn, J.; Soscia, D.; Kulp, K.; Wheeler, E.; Fischer, N. Tissue-specific extracellular matrix accelerates the formation of neural networks and communities in a neuron-glia co-culture on a multi-electrode array. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Izhikevich, E. Dynamical Systems in Neuroscience: The Geometry of Excitability and Bursting. Dyn. Syst. 2007, 441. [Google Scholar]
- Lazarevich, I.; Stasenko, S.; Rozhnova, M.; Pankratova, E.; Dityatev, A.; Kazantsev, V. Activity-dependent switches between dynamic regimes of extracellular matrix expression. PLoS ONE 2020, 15, e0227917. [Google Scholar] [CrossRef]
- Rozhnova, M.; Pankratova, E.; Stasenko, S.; Kazantsev, V. Bifurcation analysis of multistability and oscillation emergence in a model of brain extracellular matrix. Chaos Solitons Fractals 2021, 151, 111253. [Google Scholar] [CrossRef]
- Sterratt, D.; Graham, B.; Gillies, A.; Willshaw, D. Principles of Computational Modelling in Neuroscience; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Frischknecht, R.; Gundelfinger, E. The brain’s extracellular matrix and its role in synaptic plasticity. Synaptic Plast. 2012, 970, 153–171. [Google Scholar]
- Van Rossum, G.; Drake, F., Jr. Python Tutorial; Centrum voor Wiskunde en Informatica: Amsterdam, The Netherlands, 1995. [Google Scholar]
- Nelli, F.; Nelli, F. The pandas library—An introduction. In Python Data Analytics: Data Analysis and Science Using Pandas, Matplotlib, and the Python Programming Language; Apress: Berkeley, CA, USA, 2015; pp. 63–101. [Google Scholar]
- Stimberg, M.; Brette, R.; Goodman, D. Brian 2, an intuitive and efficient neural simulator. eLife 2019, 8, e47314. [Google Scholar] [CrossRef]
- Bisong, E.; Bisong, E. Matplotlib and seaborn. In Building Machine Learning and Deep Learning Models on Google Cloud Platform: A Comprehensive Guide for Beginners; Apress: Berkeley, CA, USA, 2019; pp. 151–165. [Google Scholar]
- Virtanen, P.; Gommers, R.; Oliphant, T.; Haberl, M.; Reddy, T.; Cournapeau, D.; Burovski, E.; Peterson, P.; Weckesser, W.; Bright, J.; et al. SciPy 1.0: Fundamental algorithms for scientific computing in Python. Nat. Methods 2020, 17, 261–272. [Google Scholar] [CrossRef]
- Duarte, M. Detecta: A Python Module to Detect Events in Data. GitHub Repos. 2020. Available online: https://github.com/demotu/detecta (accessed on 2 March 2023).
- Gerstner, W.; Kistler, W.; Naud, R.; Paninski, L. Neuronal Dynamics: From Single Neurons to Networks and Models of Cognition; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Stasenko, S.; Kazantsev, V. Mean-field model of tetrapartite synapse. In Proceedings of the 2022 Fourth International Conference Neurotechnologies And Neurointerfaces (CNN), Kaliningrad, Russia, 14–16 September 2022; pp. 180–184. [Google Scholar]
- Broekaart, D.; Bertran, A.; Jia, S.; Korotkov, A.; Senkov, O.; Bongaarts, A.; Mills, J.; Anink, J.; Seco, J.; Baayen, J.; et al. The matrix metalloproteinase inhibitor IPR-179 has antiseizure and antiepileptogenic effects. J. Clin. Investig. 2021, 131, e138332. [Google Scholar] [CrossRef]
- Bonneh-Barkay, D.; Wiley, C. Brain extracellular matrix in neurodegeneration. Brain Pathol. 2009, 19, 573–585. [Google Scholar] [CrossRef]
- Khoshneviszadeh, M.; Jandke, S.; Kaushik, R.; Ulbrich, P.; Norman, O.; Jukkola, J.; Heikkinen, A.; Schreiber, S.; Dityatev, A. Microvascular Damage, Neuroinflammation and Extracellular Matrix Remodeling in Col18a1 Knockout Mice as a Model for Early Cerebral Small Vessel Disease. 2022. Available online: https://europepmc.org/article/ppr/ppr539658 (accessed on 29 March 2023).
- Ulbrich, P.; Khoshneviszadeh, M.; Jandke, S.; Schreiber, S.; Dityatev, A. Interplay between perivascular and perineuronal extracellular matrix remodelling in neurological and psychiatric diseases. Eur. J. Neurosci. 2021, 53, 3811–3830. [Google Scholar] [CrossRef]
















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stasenko, S.V.; Kazantsev, V.B. Bursting Dynamics of Spiking Neural Network Induced by Active Extracellular Medium. Mathematics 2023, 11, 2109. https://doi.org/10.3390/math11092109
Stasenko SV, Kazantsev VB. Bursting Dynamics of Spiking Neural Network Induced by Active Extracellular Medium. Mathematics. 2023; 11(9):2109. https://doi.org/10.3390/math11092109
Chicago/Turabian StyleStasenko, Sergey V., and Victor B. Kazantsev. 2023. "Bursting Dynamics of Spiking Neural Network Induced by Active Extracellular Medium" Mathematics 11, no. 9: 2109. https://doi.org/10.3390/math11092109
APA StyleStasenko, S. V., & Kazantsev, V. B. (2023). Bursting Dynamics of Spiking Neural Network Induced by Active Extracellular Medium. Mathematics, 11(9), 2109. https://doi.org/10.3390/math11092109