1. Introduction
The two primary objectives of studying systems ecology are to get an understanding of the dynamics of the ecological systems and the nature of the forces, which determine the community structure. Earlier, the classical ecological models focused on two species interactions. Continuous time models of two interacting species have been studied extensively in the literature (see [
1] and references therein). Mathematically, only two basic patterns are exhibited in these models—approach to a steady state or to a limit cycle [
2]—but the ecological models of nature exhibit very complex dynamical behavior. Price et al. [
3] proposed that community behavior must be based on three or more trophic levels. Three species continuous time models have observed more complex dynamical behavior [
4,
5,
6,
7]. All these studies depend upon the classical types of functional responses, which include Holling Types I, II and III and Holling-Tanner ratio dependent functional response.
The Crowley-Martin [
8] functional response is one of the predator dependent functional responses, i.e., the functional response is function of both prey and predator abundance because of predator interference. The basic assumption behind the formulation of Crowley-Martin functional response is that there is a decrease in predator feeding rate because of high predator density even if there is a high prey density. Hence, the effect of predator interference on the feeding rate is important, even when an individual predator is handling or searching for a prey at a given instant of time. Here,
v and
r are population density of prey and predator, respectively. The parameters
are positive parameters that describe the effects of capture rate, handling time and magnitude of interference among predators on the feeding rate, respectively. The per capita rate in this formulation is given by:
The Crowley-Martin functional response is used for datasets that indicate an asymptotic feeding rate that is affected by predator density. A continuous time model to analyze the dynamics of a three species food chain with Crowley-Martin functional response was studied by Upadhyay et al. [
9]. The model system in [
9] is based on the assumption that the species under consideration are spatially homogeneously distributed; however, in nature, the species distribution is always inhomogeneous. Therefore, to model a realistic food chain scenario, reaction diffusion mechanism should be considered, as employed in [
10,
11,
12]. A qualitative analysis of a two species model with Crowley-Martin functional response and diffusion is carried out in [
13]. In the above model, the movement of a species is determined solely by their own characteristics, i.e., the movements of the species in these models are physically affected by the population pressures due to the mutual interference among the individuals of the same species. Therefore, such models take into account only the self diffusion of the species in concern. However, in the case of prey-predator interacting systems, the movement of one species may affect the movement or motility of the other. The above models fail to describe such situation, which is more realistic. In many systems, the predators develop migratory strategies to take advantage over the prey. In the case of a three species prey-predator model, such a migratory behavior depends on the concentration of both predators (i.e., the mid level predator and the top level predator). This leads to
cross diffusive system, in addition to each species natural tendency to diffuse i.e., self diffusion; such models are studied in [
14,
15,
16,
17,
18]. The cross diffusive coefficient may have positive or negative values. The positive cross diffusivity coefficient represents that one species tends to move in the direction of lower concentration of another species. The negative cross diffusion term represents that the population flux of one species is in the direction of higher concentration of the other species.
In the present paper, we consider a tritrophic food chain model in which the species interact via Crowley-Martin and Holling type II functional response at different levels. We introduce a nonlinear cross diffusion among top level and mid level predators. The objective is to understand the effect of both types of diffusion self and cross on the stability of the system. An analytic approach is adopted to understand the formation of inhomogeneous steady state solutions. Numerical simulation depicting spatial pattern formation was performed for a wide range of parameters to understand the predation behavior due to the effect of cross and self diffusion.
In
Section 2, we formulate the temporal model using various assumptions and spatially extend it to formulate a nonlinear reaction diffusion system and with cross diffusivity (a strongly coupled parabolic system). In
Section 3, we perform a linear stability analysis of the temporal model and obtain the conditions under which the stationary uniform solution is locally asymptotically stable. In
Section 4, we provide analytical proof of stability of the stationary uniform solution in the presence of diffusion and in the absence of cross diffusion but unstable in the presence of cross diffusion. In
Section 5, we provide an analytical proof of the existence of inhomogeneous steady states.
Section 6 deals with the numerical simulation of the cross diffusive system and explains Turing pattern formation for different values of cross diffusive and self diffusive coefficients of the top level predator.
2. Model System
We first consider a temporal model governing the dynamics of tritrophic food chain, consisting of Holling type-II and Crowley Martin type functional responses, defined by a system of differential equations [
9]. Here,
is the population density of the lowest trophic level species (prey),
is the population density of the middle trophic level species (intermediate predator) and
is the population density of the highest trophic level species (top predator), at any time
T. We made the following basic assumptions for formulation of the proposed model system.
Assumption 1. The prey U grows with an intrinsic growth rate , and has a carrying capacity of K in the absence of predator V. D is a measure of the extent to which the environment can provide protection to U and w is the maximum value which per capita reduction rate of U can attain. In addition, the mid level predator V predates on U in accordance with Holling Type II functional response, . Therefore, considering the above assumptions, we formulate the first equation of the model system as: Assumption 2. The intermediate or mid level predator V has a natural death rate of . describes the maximum value which per capita reduction rate of V can attain, is similar to D of U and p represents the internal competition coefficient among the members of V. The predation of V over U is governed by Holling Type II functional response. The top level predator R predates on V in accordance with Crowley-Martin functional response. The constants , b and d are the saturating Crowley-Martin type functional response parameters, where describes capture rate effect, b represents handling time of prey and d is the magnitude of interference among predators. The second equation of the model system is given as: Assumption 3. The top level predator R dies at a natural death rate of c. As the predating behavior of R over V is described by the Crowley-Martin functional response. is the saturating Crowley-Martin type functional response parameter similar to . The third equation of the model system takes the following form: All parameters described above take only positive values and the model system is a three species food chain model involving a hybrid type of prey dependent and predator dependent functional response. Therefore, using the above assumptions, the model system takes the following form:
The model system described by Equation (
4) consists of thirteen parameters, which makes the analysis quite cumbersome, therefore the number of parameters are reduced by rescaling the model system. The model is rescaled, using the following new variables and parameters:
The rescaled system is as follows:
The model system proposed in Equation (
4) and the rescaled form of the model given in Equation (
6) are based on the assumption that the species under consideration are homogeneously distributed but in nature the species distribution is always inhomogeneous. Therefore, to model a realistic food chain scenario, we consider the model system with diffusion. Consider the spatially explicit three species predator prey food chain model system. At any location
and time
t, the interaction of three species populations
,
and
can be modeled with the reaction diffusion equations given in Equation (
7). Here,
denotes two dimensional Laplacian operator given by
, where,
and
.
is a bounded domain in
with a smooth boundary
.
The boundary conditions are directional derivative normal to the boundary and n is the outward normal to .
In the above system, the movement of a species is determined solely by their own characteristics, i.e., the movements of the species can be physically affected by the population pressures due to the mutual interference between the individuals of the same species. Therefore, we refer to the constants
,
and
as the
self diffusion rates of species
u,
v and
r, respectively. However, in the case of a prey-predator interacting system, the movement of one species may affect the movement or motility of the other. Hence, the above model cannot be used to describe such a situation. There are chances of development of migratory strategies among the predators to take advantage over the prey. This behavior can be described by taking into account the effect of
cross diffusion. Along with the self tendency to move, i.e., self diffusion, the species also migrate or move under the influence of other. Such behavior takes into account concentration of both predators—the mid and the top level predators—constituting a cross diffusive system. The coefficient of cross diffusion may be positive or negative. The positive cross diffusion coefficient represents that one species tends to move in the direction of lower concentration of another species. The negative cross diffusion term represents that the population flux of one species is in the direction of higher concentration of the other. Therefore, we propose the interaction of the above three species population model with cross diffusion as follows:
where
denotes two dimensional Laplacian operator given by
,
.
The nonlinear diffusion terms in the equation governing the dynamics of top level predator
r implies that the direction of dispersion of top predator contains a self diffusion term by which it moves from a region of its higher concentration to a region of its lower concentration and a cross diffusive term. The top predator
r diffuses with the flux
where the
, the
part of the flux
is directed towards the decreasing population density of mid level predator
v. This is because, in certain prey-predator systems, several prey form a huge group to protect themselves from the attacking predators. In addition, as in the model system the predation of top predator is impossible, to extensively study the role of cross diffusion on the model system, we induce it at
r, i.e., the top predator.
4. Analysis of Spatially Extended Model
In this section, we focus on the spatiotemporal models in Equation (
7) in presence of diffusion but absence of cross diffusion and Equation (
8) in presence of both self and cross diffusion. Our objective is to derive the conditions under which the positive equilibrium solution is stable with diffusion but without cross diffusion but unstable with diffusion and cross diffusion both. This phenomenon is called
cross diffusion driven instability [
16,
18].
4.1. Without Cross Diffusion
We now consider the system in Equation (
7). To discuss the local stability analysis of the system of parabolic Equations (
7) and (
8), we use the notation used in [
16].
Notation 1. Let be the eigenvalues of on Ω under no-flux boundary conditions, and be the space of eigenfunctions corresponding to . We define the following space decomposition
- (i)
where are orthonormal basis of for .
- (ii)
, and so , where .
Theorem 2. The positive equilibrium solution of the model system in Equation (7) is locally asymptotically stable if Proof. The spatially extended model system without cross diffusion in Equation (
7) has been linearized at
and expressed as [
16]:
where,
Notation 1 suggests that
is invariant under the operator
, and
is an eigenvalue of this operator on
if and only if it is an eigenvalue of the matrix in
. The characteristic polynomial of the matrix
is given by
where
□
By using the stated criteria in Theorem 2, it seems clear that
,
,
and
. Therefore, it follows from the Routh-Hurwitz criteria that, for each
, all three roots
of
have negative real parts. Hence, the positive equilibrium solution
of Equation (
7) is locally asymptotically stable under the stated condition in Equation (
12).
From Theorem 2, it is clear that addition of self diffusion terms to the temporal model system in Equation (
6) results in stable positive equilibrium solution under the stated condition. Therefore, diffusion driven instability has not yet occurred. Now, we now consider the effect of cross diffusion on the model system.
4.2. With Cross Diffusion
In this section, we study in detail the system given in Equation (
8).
Theorem 3. Provided that Theorem 2 holds, assuming and considering the following inequality holds,if , where is as explained in Notation 1 and is given by Equation (24), then there exists , such that, when , the positive equilibrium solution of the cross diffusive system in Equation (8) becomes unstable. We denote
. Linearizing the system in Equation (
8) at
, we have
where,
The characteristic polynomial of
is given by,
where
If
are the three roots of
, then
For at least one
Re, it suffices to show that
. In addition,
From the above expression, we clearly see that .
Let,
and let
be the three roots of
, such that
Re Re Re. Now,
As
due to condition specified in Equation (
12), we have,
As and , . Using the theory of equations if one of , , is real and negative and the product of the other two is positive.
To obtain the values of
, we calculate the following limits:
If
, then the following inequality holds,
We have, .
In addition,
As
, which means that the equation,
has one positive root and one negative root. From continuity, if
,
is real and negative. Since
,
and
are real and positive.
Therefore, there exists such that when then the specified criteria holds. In addition, when . Therefore, when , then , it follows that . Therefore, , and the proof is complete.
Therefore, from the above theorems, we conclude that cross diffusion destabilizes the stationary uniform solution.
5. Inhomogeneous Steady States
In this section, the justification for the cross diffusion driven instability phenomenon is explained. We now prove that model system in Equation (
8) admits the inhomogeneous steady state. We, consider the steady state model system in Equation (
8) in the following form,
The constants depend on the domain . As is fixed, this dependance is not mentioned explicitly. The parameters are collectively denoted by .
5.1. A Priori Estimates
We now attempt to give an a priori upper and lower bounds for the positive solutions to Equation (
26). We use Harnack Inequality and Maximum Principle (for details, refer to [
16,
18]).
Proposition 1. Harnack Inequality Let be a positive solution to , where , satisfying the homogeneous Neumann boundary condition. Then, there exists a positive constants that depends only on such that Proposition 2. Maximum principle Let and
- 1.
If satisfiesand , then . - 2.
If satisfiesand , then .
Theorem 4. (Upper Bound) Let be fixed positive constants. Then, there exists positive constants and such that when ; ; and , the positive equilibrium solution of Equation (26) satisfies, Proof. We apply the maximum principle to the first part of Equation (
26) and get
. Similarly, applying the maximum principle to the second part of Equation (
26) gives
.
Let
. Let
be such that
. Now, applying the maximum principle to the third part of Equation (
26) gives,
.
Defining
as
, we have,
Using boundedness of
, we apply the Harnack inequality to the first and second parts of Equation (
26), yielding
Let
and we have,
where
is bounded.
By Harnack inequality,
for some positive constant
Lemma 1. Let , be positive constants, and be the corresponding positive equilibrium solution of Equation (26) with . If as and is a constant vector, then , where is the nontrivial solution of . Proof. For all . If , then when m is large. However, since is positive, this is not possible. Similarly, is not possible. Therefore, . The same argument shows that . □
Theorem 5. Lower Bound: Let , , and be fixed positive constants. There exists positive constants such that, when , , and , the positive solution of Equation (26) satisfies, Proof. We integrate the second part of Equation (
26) in
and consider the inhomogeneous Neumann boundary condition, yielding
therefore, there exists
such that
. □
As
,
. By using Harnack inequality,
Now, integrating third part of Equation (
26) in
With reference to Equation (
27), to prove Theorem 5, it is sufficient to show that,
Let the inequality in Equation (
29) not hold. Then, there exists sequences
with
and the corresponding positive solutions
to Equation (
26) such that
where,
satisfies
Since , we may assume that and , where and are positive constants. In addition, we claim that and converge uniformly to positive constants, respectively. In fact, there are two cases of to be considered.
Case 1 is bounded with respect to
m. Set,
Using upper bound of
, we have
for all
. Since
satisfies
using
estimate and the Sobolev embedding theorems, we have
Similarly, the
norms of
is uniformly bounded with respect to
m. Thus, we assume that
in
. By the definition of
, for sufficiently large
m we have,
Since
and
, we have
Hence, ; otherwise, on that implies , which is a contradiction to , therefore .
Case 2 is unbounded with respect to
m. We may assume that
. Set
Therefore, proceeding as above
Hence, . Similarly, we can prove that constant . The above argument shows that there are positive constants , such that . This is contradiction to Lemma 5.1, thus the proof is completed.
5.2. Existence of Inhomogeneous Positive Steady States
We, now discuss the existence of inhomogeneous positive solutions. Let
be defined as in Notation 1. Define
Let
. Then, Equation (
26) is transformed as follows,
As the determinant of
is positive for all non-negative
,
exists and
is positive. Hence,
is a positive solution to Equation (
31) if and only if
where
is the inverse of
in
, where
is a compact perturbation of the identity operator, for any
. the Leray-Schauder degree
is well defined if
on
.
Furthermore, we note that
and as
is invertible, the index of
F at
is defined as
, where
is the number of negative eigenvalues of
.
For every integer
and for each integer
,
is invariant under
, and
is an eigenvalue of
on
if and only if it is an eigenvalue of the matrix,
Thus,
is invertible if and only if, for all
, the matrix
is non-singular. We have,
Further, if , then for each , the number of negative eigenvalues of on is odd if and only if . We have results as follows.
Proposition 3. Suppose that for all, , the matrix is nonsingular. Then, To compute
, we consider the sign of
. As we want to study the existence of positive solutions of Equation (
31) with respect to
, we focus on dependence of
on
, and consider
and
to be fixed. We denote,
As , we only need to consider . In fact , and by Theorem 3. Therefore, we make the following proposition.
Proposition 4. Assuming the condition specified in Theorem 3 holds, then there exists a positive number such that, for all , the roots of are all real and satisfy Equations (23)–(25). Moreover, for all . To discuss the effect of cross diffusion on the existence of inhomogeneous positive solution to Equation (
31), we first deduce a non-existence result when the cross diffusion term is absent.
Theorem 6. Suppose thatwhere is given previously. Then, there exists a positive constants when without cross diffusion term , Equation (26) has no non-constant positive solution. Furthermore, Proof. Assume
is a positive solution of Equation (
26) with
.
Multiplying the first part of Equation (
26) by
, the second part by
and the third part by
(taking
) and integrating by parts over
, we have
By using Cauchy inequality, we have
where
is any arbitrary positive constant.
By Poincare inequality we have,
By Theorem 5.1, we can choose a sufficiently small such that,
We conclude that
, which completes the proof. Therefore, we conclude that the model system has no non-constant positive solution. Now, we prove that in the presence of cross diffusion inhomogeneous solution is generated. The following theorem discusses the global existence of of inhomogeneous solution of Equation (
8) with respect to
as the other parameters
,
and
are fixed.
Theorem 7. Let be fixed and satisfy conditions of Theorems 2 and 3. Let be defined in Equations (23)–(25). If for some , and the sum is odd. Then, there exist a positive number such that, if , Equation (8) has at least one inhomogeneous positive steady state solution. Proof. Let
and
be positive constants and satisfy
,
,
and
. By Equations (
23)–(
25) and Proposition 4, there exists a positive constant
such that, when
, Equation (
32) holds and
□
Now, we prove that for,
, Equation (
26) has at least one inhomogeneous positive solution. The statement has been proved by contradiction and is based on homotopy invariance of topological degree. On the contrary, we assume that the assertion is not true. For
, define
where,
and consider
Then,
is a positive inhomogeneous solution of Equation (
26) if and only if it is a solution of Equation (
34) for
.
is the unique constant solution of Equation (
34) for any
. Now, we have for any
,
is a unique positive solution of Equation (
34) if and only if,
As
, by Theorem 4.2,
has only positive solution
in
, therefore we have,
where,
. Using Proposition 4 and Equation (
33), it follows,
Therefore, 0 is not an eigenvalue of the matrix
for all
, and
By Proposition 3,
By Theorems 3 and 4, there exists a positive constants
C such that, for all
, the positive solutions of Equation (
34) satisfying
. Therefore,
on
for all
. By the homotopy invariance of the topological degree,
However, by our deduction, both equations
and
have only the positive solution
in
, and hence by Equations (
35) and (
36)
which contradicts Equation (
37) and we complete the proof.
6. Numerical Simulation
The numerical simulation was carried out to understand the spatiotemporal dynamics of top predator
r under the influence of cross diffusion. For this purpose, in this section, we present a detailed investigation of the patterns in the top level predator for different diffusivity coefficients. The system of partial differential equations given in Equation (
8) was numerically solved using semi implicit finite difference technique. Forward difference scheme was used for the reaction terms and standard five point explicit finite difference scheme was used for diffusion term. Turing patterns were obtained from the effect of nonlinear diffusion term for the top level predator
r as in [
18,
19,
20].
To discretize the system, we considered Taylor’s series expansion about the non-trivial equilibrium point
of the cross diffusive term of the third equation describing the dynamics of
r for the model system in Equation (
8). We obtained a system of the following form:
The space step size and time step size were chosen appropriately to ensure the convergence of the scheme.
We used the standard five-point approximation for the two-dimensional Laplacian with the zero-flux boundary conditions. Initially, the entire system was at the stationary state
. The perturbation introduced in the initial condition was of the order
, as given in Equation (
39):
where
,
.
The set of parameter values at which the system would yield a locally asymptotically stable solution is:
To analyze the role of cross diffusion on
r, we considered the following set of parameters:
The parameter values of diffusivity coefficients are presented in
Table 1. We performed the simulation on a
grid with spatial step size
and time step size
. To investigate the role of cross diffusion and self diffusion in the pattern formation of the top predator
r, we performed simulations for a wide range of self diffusive coefficient
and cross diffusivity coefficient
. The different values of self and cross diffusive coefficients used in numerical experiments of top level predator
r are presented in
Table 1. We carried out all the numerical simulation at time level
t = 10,000 for the model system given in Equation (38).
Here, we performed numerical simulation to investigate the role of self and cross diffusion on pattern formation. In
Figure 1, we notice that, in the absence of cross diffusion, i.e.,
, no patterns were formed for the top level predator
r, of the system in Equation (38). The behavior persists even for the higher values of self diffusivity coefficients
. This numerical experiment shows that in the absence of cross diffusion there is no destabilizing effect on the system even when there is self diffusion in the system.
In our second numerical experiment, we introduced the cross diffusion by gradually increasing the cross diffusivity coefficient
and along with that we gradually decreased the self diffusivity coefficient
. The goal was to see the impact of cross diffusivity on the dynamics when there is very less role of self diffusion in the system. To this end, in
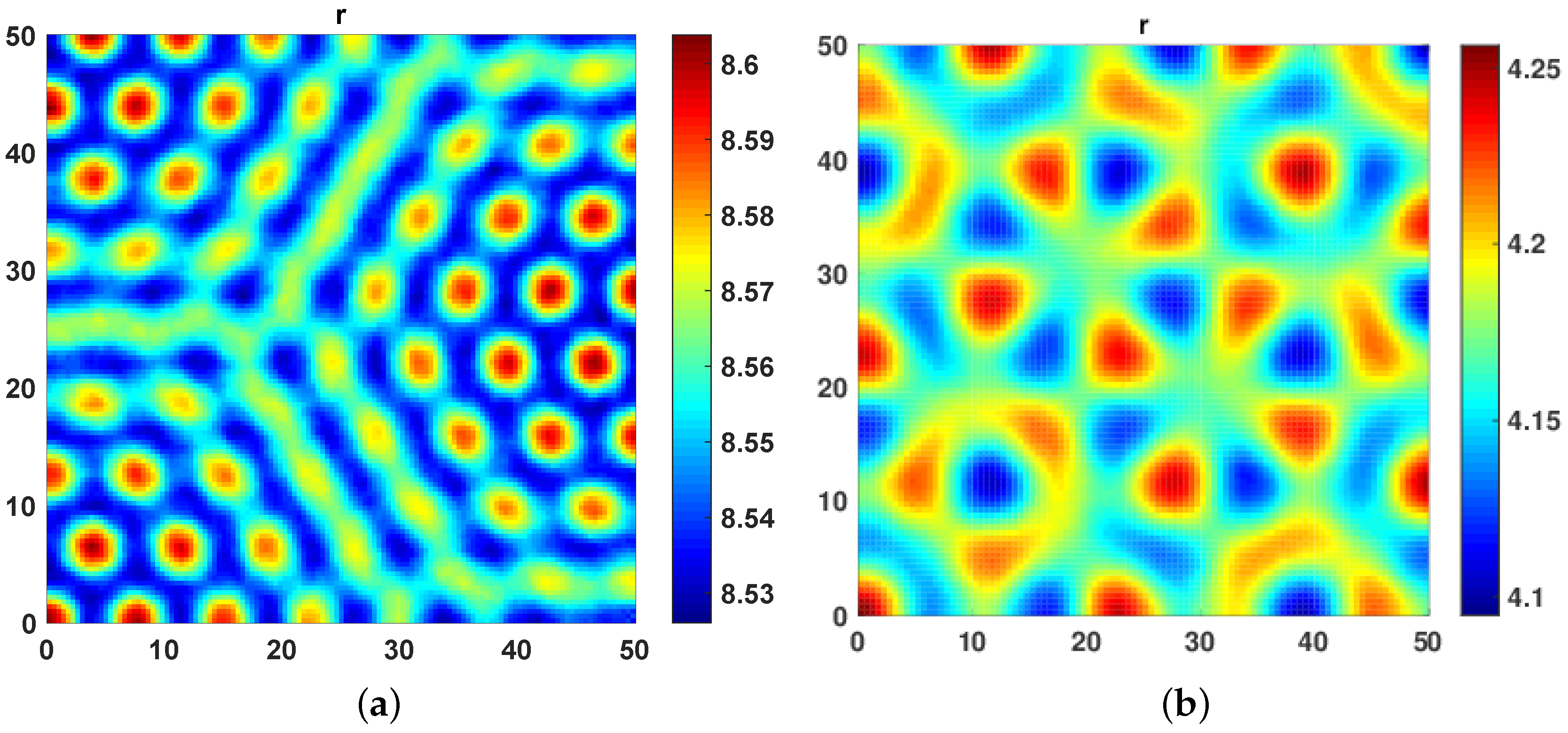
Figure 2a, we increase
from 0 to
and decrease
from 10 to
. An increase in cross diffusion coefficient results in destabilization in the top predator
r dynamics leading to a mixture of hot spots and labyrinth patterns, as seen in
Figure 2a. We further increased
to 5 and decreased
to 1 to observe floral turing patterns (
Figure 2b).
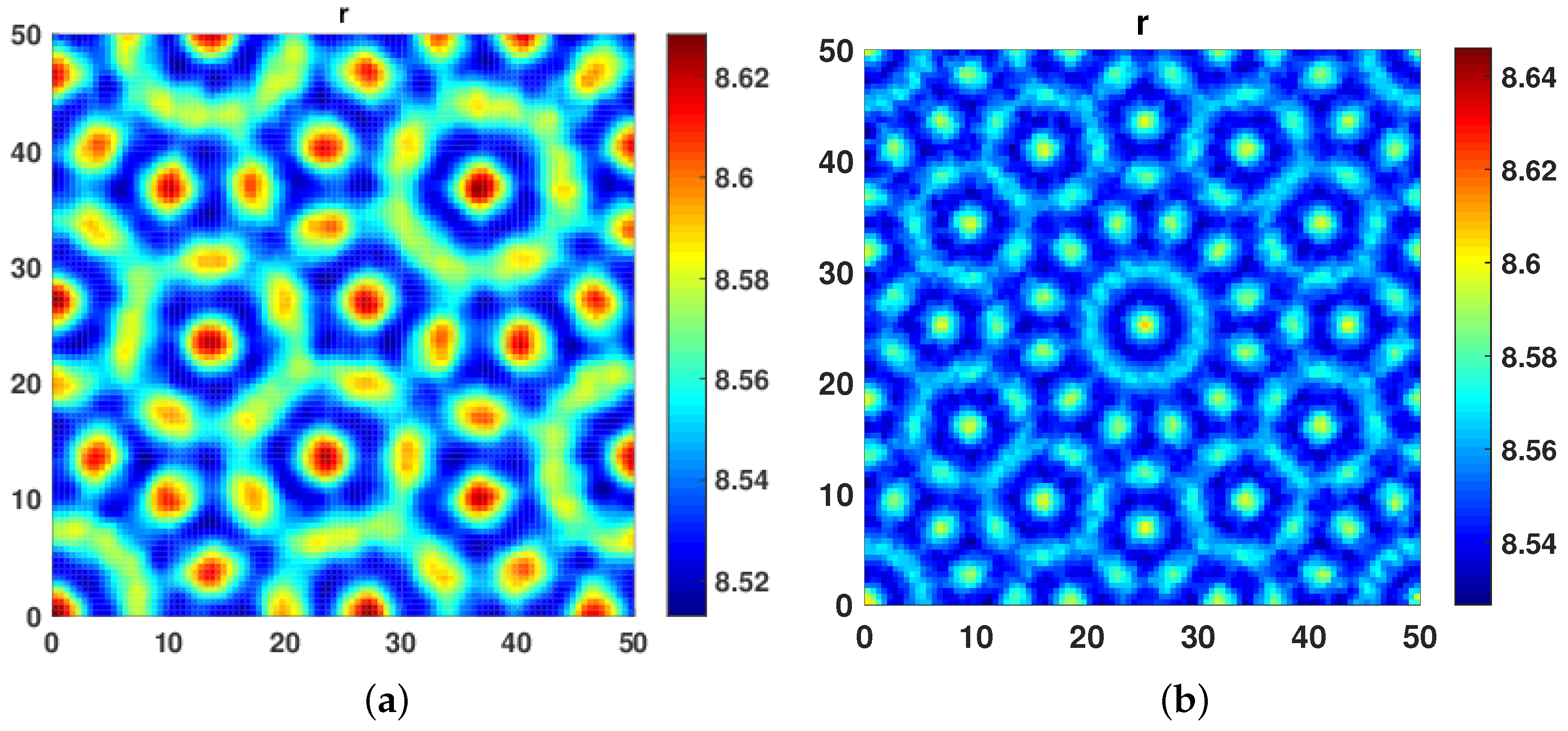
On further increasing the cross diffusivity coefficient at
and taking self diffusion coefficient at
, we observed pentagonal Turing patterns, as shown in
Figure 3a. Further, at
and
, we obtained Floral Turing Pattern.
Increasing the value of
to 10 and fixing
, we observed hot spothl turing patterns, as given in
Figure 4a at
. At
, a mix of hot spot and Labyrinth Turing patterns were observed, as shown in
Figure 4b.
As shown in
Figure 5a,b, we observed that, for a very high value of cross diffusion coefficients
and
, and very low values of
and
, hot spot and Hexagonal Spot Turing patterns are obtained, respectively.
From these experiments, we can conclude that the role of cross diffusion in destabilizing the dynamics is much more significant than self diffusion.
In our third numerical experiment, we started with equal values of both
and
fixed at
. The hot spot patterns began to appear. Further, when we increased diffusivity coefficient
and
to 1, a mix of hot spot and Labyrinth Turing Patterns were seen. We further increased both
and
with the restriction that both self and cross diffusion coefficient must remain the same. We observed that the patterns became more conspicuous, leading to a mix of hot spot and Labyrinth Turing Patterns (see
Figure 6a,b and
Figure 7a,b).
Through extensive simulations over a wide range of diffusivity coefficients, we observed that cross diffusion coefficient
plays an important role to realize the phenomenon of pattern formation. In the absence of cross diffusion, no patterns were formed, even at higher values of
, as shown in
Figure 1. Moreover we also determined the Turing patterns in the cases where cross diffusion coefficient equals to the self diffusion coefficient, i.e.,
.
Table 1 describes the various self and cross diffusivity coefficients used for obtaining the spatial patterns of the system in Equation (38).
Table 1 indicates a negative correlation between self diffusion coefficient and cross diffusion coefficients, which results in pattern formation. The above simulations results are coherent to our analytical results that non-constant positive solutions exist only in the presence of cross diffusion hence the pattern formation.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}