
Supplementation of Coenzyme Q10 among Patients with Type 2 Diabetes Mellitus

Abstract
:1. Introduction
2. Mitochondria Function and Coenzyme Q10
2.1. Mitochondrial Function
2.2. Reactive Oxygen Species
2.3. Coenzyme Q10
2.4. Role of Mitochondria in Pancreatic Beta Cells
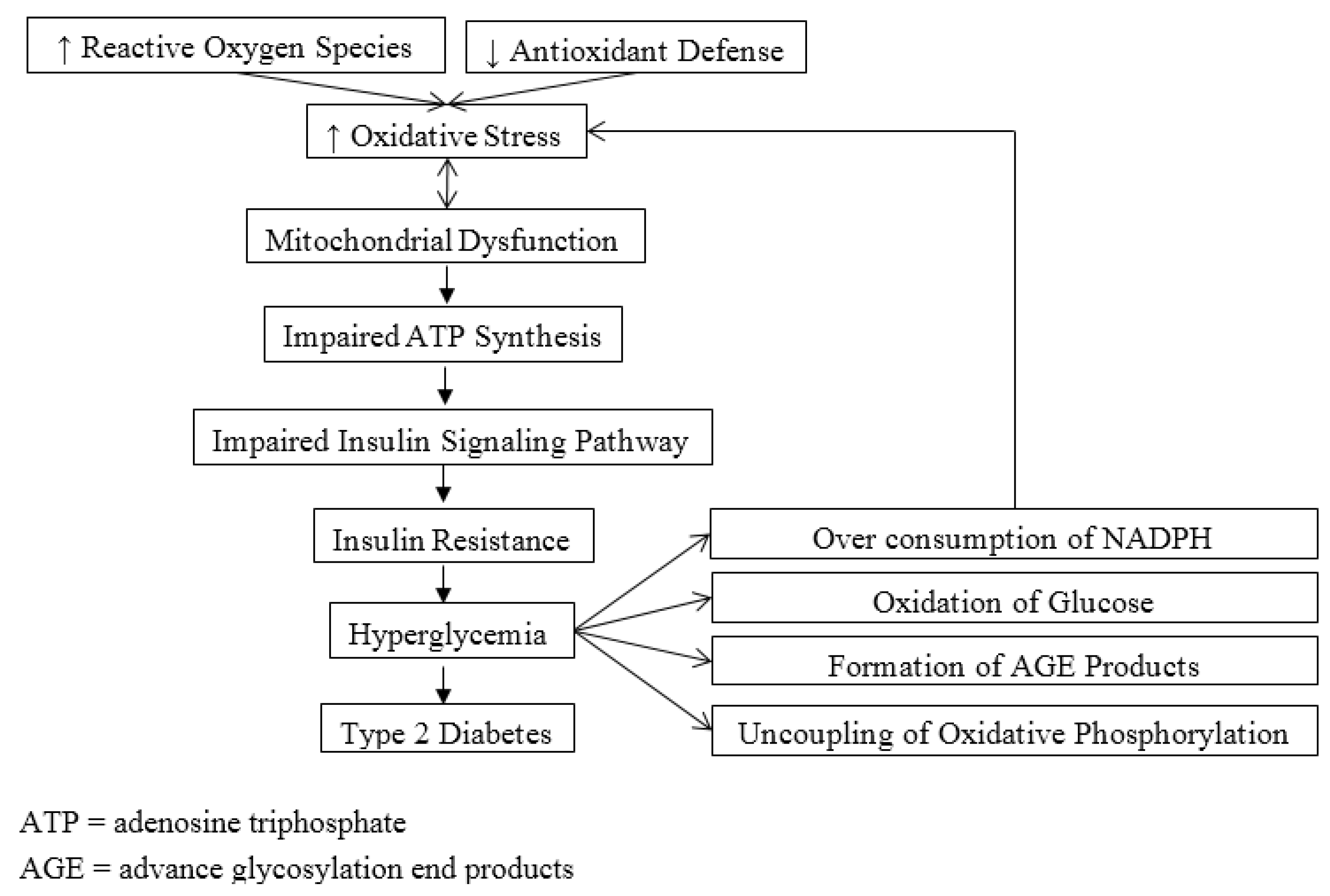
3. Mitochondrial Dysfunction, Oxidative Stress and Diabetes

4. Coenzyme Q10 Deficiency in Type 2 Diabetes
5. Supplement Use of Coenzyme Q10 in Type 2 Diabetes

{kind=link}
| Authors and Year of Publication | Study Design and Subjects | Form and Dosage of Coenzyme Q10 | Results |
|---|---|---|---|
| Shigeta et al. 1966 [38] | 39 patients with diabetes | CoQ7 120 mg for 2–18 weeks | There was significant decrease in glucose level (≥20%) in 67% of patients |
| Shimura et al. 1981 [39] | 15 patients with diabetes | CoQ10 60 mg daily for 12 weeks | There was significantly increased in insulin synthesis and secretion. Glycemic control was improved |
| Conget et al. 1996 [41] | Rat islets | 2, 4, and 8 µM CoQ10 | There was no significant increase in release of insulin.25% increase in insulin secretion was observed when a higher concentration (8 µM) was used |
| Eriksson et al. 1999 [30] | Double-blind placebo-controlled study 23 patients with T2DM | CoQ10 100 mg twice a day for 6 months | There was no significant improvement in blood glucose and HbA1c levels |
| Henriksen et al. 1999 [42] | Randomized, double-blind, placebo-controlled study 34 patients with T1DM | CoQ10 100 mg daily for 12 weeks | There was not significant decrease in HbA1c and blood glucose levels |
| Singh et al. 1999 [43] | Patients with hypertension and coronary artery disease | CoQ10 60 mg twice a day for 8 weeks | There was significantly decrease in blood pressures. Fasting and 2 h plasma glucose and insulin levels were significantly reduced. There was significant increase in other antioxidants such as vitamin A, E, and C and beta-carotene. Markers for oxidative stress (TBARS, malondialdehyde, and diene conjugates) were significantly reduced. |
| Hodgson et al. 2002 [44] | 74 patients with uncomplicated T2DM and dyslipidemia | CoQ10 200 mg/day for 12 weeks | HbA1c level was significantly decreased. No significant improvement in fasting blood glucose and insulin. No significant change in oxidative stress status measured by F2-isoprostane |
| Lim et al. 2008 [45] | 80 patients with T2DM | CoQ10 200 mg/day for 12 weeks | There was significant increase in plasma total CoQ10 level but no change in ubiquinol level. No significant improvement in HbA1c level. No significant improvement in microcirculatory endothelial function. |
| Sena et al. 2008 [46] | Type 2 diabetic GK rat model | CoQ10 20 mg/kg body weight and/or α-tocopherol for 8 weeks | There was significant decrease in HbA1c level. No significant improvement in fasting and 2 h blood glucose levels. |
| Mezawa et al. 2012 [10] | 9 patients with T2DM 5 healthy volunteers | Ubiquinol 200 mg daily for 12 weeks (T2DM patients) Ubiquinol 200 mg daily for 4 weeks | There was significant improvement in HbA1c (from 7.1 ± 0.4 to 6.8% ± 0.4%) among patients with T2DM. There was significant increase in insulin genic index (0.65 ± 0.29 to 1.23 ± 0.56) and decrease in proinsulin to insulin ratio (3.4 ± 1.8 to 2.1 ± 0.6) in healthy volunteers, indicating increased insulin secretion. |
| Sourris et al. 2012 [9] | Diabtic nephropathy rodent model | CoQ10 10 mg/kg/day for 10 weeks | There were significantly decreases in urinary albumin excretion in 24 h, albumin/creatinine ratio, and tubulointerstitial collagen deposition. |
| Kolahdouz Mohammadi et al. 2013 [47] | Randomized double-blind placebo-controlled trial 64 patients with T2DM | CoQ10 200 mg daily for 12 weeks | There was significant decrease in HbA1c in the CoQ10 group. |
6. Conclusions
Author Contributions
Conflicts of Interest
References
- Centers for Disease Control and Prevention. National Diabetes Statistics Report: Estimates of Diabetes and Its Burden in the United States, 2014; USA Department of Health and Human Services: Atlanta, GA, USA, 2014. [Google Scholar]
- International Diabetes Federation. Idf Diabetes Atlas; International Diabetes Federation: Brussels, Belgium, 2013. [Google Scholar]
- American Diabetes Association. Standards of medical care in diabetes—2014. Diabetes Care 2014, 37, S14–S80. [Google Scholar]
- White, M.F.; Myers, M.G.J. The molecular basis of insulin action. In Endocrinology; de Groot, L.J., Jameson, J.L., Eds.; WB Saunders Co.: Philadelphia, PA, USA, 2001; pp. 712–727. [Google Scholar]
- Lamson, D.W.; Plaza, S.M. Mitochondrial factors in the pathogenesis of diabetes: A hypothesis for treatment. Altern. Med. Rev. 2002, 7, 94–111. [Google Scholar] [PubMed]
- Ates, O.; Bilen, H.; Keles, S.; Alp, H.H.; Keles, M.S.; Yildirim, K.; Ondas, O.; Pinar, L.C.; Civelekler, M.; Baykal, O. Plasma coenzyme Q10 levels in type 2 diabetic patients with retinopathy. Int. J. Ophthalmol. 2013, 6, 675–679. [Google Scholar] [PubMed]
- El-ghoroury, E.A.; Raslan, H.M.; Badawy, E.A.; El-Saaid, G.S.; Agybi, M.H.; Siam, I.; Salem, S.I. Malondialdehyde and coenzyme Q10 in platelets and serum in type 2 diabetes mellitus: Correlation with glycemic control. Blood Coagul. Fibrinolysis 2009, 20, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, G.; Yamamoto, Y.; Zhi, J.G.; Tanino, Y.; Yamasaki, M.; Yano, M.; Nakajima, T.; Fukui, M.; Yoshikawa, T.; Nakamura, N. Daily profile of plasma %CoQ10 level, a biomarker of oxidative stress, in patients with diabetes manifesting postprandial hyperglycaemia. Acta Diabetol. 2005, 42, 179–181. [Google Scholar] [CrossRef] [PubMed]
- Sourris, K.C.; Harcourt, B.E.; Tang, P.H.; Morley, A.L.; Huynh, K.; Penfold, S.A.; Coughlan, M.T.; Cooper, M.E.; Nguyen, T.V.; Ritchie, R.H.; et al. Ubiquinone (coenzyme Q10) prevents renal mitochondrial dysfunction in an experimental model of type 2 diabetes. Free Radic. Biol. Med. 2012, 52, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Mezawa, M.; Takemoto, M.; Onishi, S.; Ishibashi, R.; Ishikawa, T.; Yamaga, M.; Fujimoto, M.; Okabe, E.; He, P.; Kobayashi, K.; et al. The reduced form of coenzyme Q10 improves glycemic control in patients with type 2 diabetes: An open label pilot study. Biofactors 2012, 38, 416–421. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A.; Rahman, M.M. Mitochondrial dysfunction in obesity: Potential benefit and mechanism of co-enzyme Q10 supplementation in metabolic syndrome. J. Diabetes Metab. Disord. 2014, 13, 60. [Google Scholar] [CrossRef] [PubMed]
- Svensson, O.L. Mitochondria: Structure, Functions and Dysfunctions, 1st ed.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2011. [Google Scholar]
- Yakes, F.M.; van Houten, B. Mitochondrial DNA damage is more extensive and persists longer than nuclear DNA damage in human cells following oxidative stress. Proc. Natl. Acad. Sci. USA 1997, 94, 514–519. [Google Scholar] [CrossRef] [PubMed]
- Genova, M.L.; Lenaz, G. New developments on the functions of coenzyme Q in mitochondria. Biofactors 2011, 37, 330–354. [Google Scholar] [CrossRef] [PubMed]
- Cotan, D.; Cordero, M.D.; Garrido-Maraver, J.; Oropesa-Avila, M.; Rodriguez-Hernandez, A.; Gomez Izquierdo, L.; de la Mata, M.; de Miguel, M.; Lorite, J.B.; Infante, E.R.; et al. Secondary coenzyme Q10 deficiency triggers mitochondria degradation by mitophagy in melas fibroblasts. FASEB J. 2011, 25, 2669–2687. [Google Scholar] [CrossRef] [PubMed]
- Yubero-Serrano, E.M.; Gonzalez-Guardia, L.; Rangel-Zuniga, O.; Delgado-Lista, J.; Gutierrez-Mariscal, F.M.; Perez-Martinez, P.; Delgado-Casado, N.; Cruz-Teno, C.; Tinahones, F.J.; Villalba, J.M.; et al. Mediterranean diet supplemented with coenzyme Q10 modifies the expression of proinflammatory and endoplasmic reticulum stress-related genes in elderly men and women. J. Gerontol. A Biol. Sci. Med. Sci. 2012, 67, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Mourier, A.; Motori, E.; Brandt, T.; Lagouge, M.; Atanassov, I.; Galinier, A.; Rappl, G.; Brodesser, S.; Hultenby, K.; Dieterich, C.; et al. Mitofusin 2 is required to maintain mitochondrial coenzyme Q levels. J. Cell Biol. 2015, 208, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Noh, Y.H.; Kim, K.Y.; Shim, M.S.; Choi, S.H.; Choi, S.; Ellisman, M.H.; Weinreb, R.N.; Perkins, G.A.; Ju, W.K. Inhibition of oxidative stress by coenzyme Q10 increases mitochondrial mass and improves bioenergetic function in optic nerve head astrocytes. Cell Death Dis. 2013, 4, e820. [Google Scholar] [CrossRef] [PubMed]
- Bournat, J.C.; Brown, C.W. Mitochondrial dysfunction in obesity. Curr. Opin. Endocrinol. Diabetes Obes. 2010, 17, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Brons, C.; Jacobsen, S.; Hiscock, N.; White, A.; Nilsson, E.; Dunger, D.; Astrup, A.; Quistorff, B.; Vaag, A. Effects of high-fat overfeeding on mitochondrial function, glucose and fat metabolism, and adipokine levels in low-birth-weight subjects. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E43–E51. [Google Scholar] [CrossRef] [PubMed]
- Chew, G.T.; Watts, G.F. Coenzyme Q10 and diabetic endotheliopathy: Oxidative stress and the “recoupling hypothesis”. QJM 2004, 97, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, M.T.; Mibus, A.L.; Forbes, J.M. Oxidative stress and advanced glycation in diabetic nephropathy. Ann. N.Y. Acad. Sci. 2008, 1126, 190–193. [Google Scholar] [CrossRef] [PubMed]
- Forbes, J.M.; Coughlan, M.T.; Cooper, M.E. Oxidative stress as a major culprit in kidney disease in diabetes. Diabetes 2008, 57, 1446–1454. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, T.; Edelstein, D.; Du, X.L.; Yamagishi, S.; Matsumura, T.; Kaneda, Y.; Yorek, M.A.; Beebe, D.; Oates, P.J.; Hammes, H.P.; et al. Normalizing mitochondrial superoxide production blocks three pathways of hyperglycaemic damage. Nature 2000, 404, 787–790. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.L.; Goldfine, I.D.; Maddux, B.A.; Grodsky, G.M. Oxidative stress and stress-activated signaling pathways: A unifying hypothesis of type 2 diabetes. Endocr. Rev. 2002, 23, 599–622. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.C.; Tan, H.H.; Goh, S.K.; Subramaniam, T.; Sum, C.F.; Tan, I.K.; Lee, B.L.; Ong, C.N. Oxidative burden in prediabetic and diabetic individuals: Evidence from plasma coenzyme Q(10). Diabet. Med. 2006, 23, 1344–1349. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, S.; Yamamoto, Y. Simultaneous detection of ubiquinol and ubiquinone in human plasma as a marker of oxidative stress. Anal. Biochem. 1997, 250, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Quinzii, C.M.; Lopez, L.C.; Naini, A.; DiMauro, S.; Hirano, M. Human CoQ10 deficiencies. Biofactors 2008, 32, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Mancino, R.; di Pierro, D.; Varesi, C.; Cerulli, A.; Feraco, A.; Cedrone, C.; Pinazo-Duran, M.D.; Coletta, M.; Nucci, C. Lipid peroxidation and total antioxidant capacity in vitreous, aqueous humor, and blood samples from patients with diabetic retinopathy. Mol. Vis. 2011, 17, 1298–1304. [Google Scholar] [PubMed]
- Eriksson, J.G.; Forsen, T.J.; Mortensen, S.A.; Rohde, M. The effect of coenzyme Q10 administration on metabolic control in patients with type 2 diabetes mellitus. Biofactors 1999, 9, 315–318. [Google Scholar] [CrossRef] [PubMed]
- Molyneux, S.L.; Young, J.M.; Florkowski, C.M.; Lever, M.; George, P.M. Coenzyme Q10: Is there a clinical role and a case for measurement? Clin. Biochem. Rev. 2008, 29, 71–82. [Google Scholar] [PubMed]
- Fernandez-Alvarez, J.; Conget, I.; Rasschaert, J.; Sener, A.; Gomis, R.; Malaisse, W.J. Enzymatic, metabolic and secretory patterns in human islets of type 2 (non-insulin-dependent) diabetic patients. Diabetologia 1994, 37, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Giroix, M.H.; Rasschaert, J.; Bailbe, D.; Leclercq-Meyer, V.; Sener, A.; Portha, B.; Malaisse, W.J. Impairment of glycerol phosphate shuttle in islets from rats with diabetes induced by neonatal streptozocin. Diabetes 1991, 40, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Ostenson, C.G.; Abdel-Halim, S.M.; Rasschaert, J.; Malaisse-Lagae, F.; Meuris, S.; Sener, A.; Efendic, S.; Malaisse, W.J. Deficient activity of FAD-linked glycerophosphate dehydrogenase in islets of GK rats. Diabetologia 1993, 36, 722–726. [Google Scholar] [CrossRef] [PubMed]
- Sener, A.; Herberg, L.; Malaisse, W.J. FAD-linked glycerophosphate dehydrogenase deficiency in pancreatic islets of mice with hereditary diabetes. FEBS Lett. 1993, 316, 224–227. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F. Can correction of sub-optimal coenzyme Q status improve beta-cell function in type II diabetics? Med. Hypotheses 1999, 52, 397–400. [Google Scholar] [CrossRef] [PubMed]
- Kishi, T.; Kishi, H.; Watanabe, T.; Folkers, K. Bioenergetics in clinical medicine. XI. Studies on coenzyme Q and diabetes mellitus. J. Med. 1976, 7, 307–321. [Google Scholar] [PubMed]
- Shigeta, Y.; LIzumi, K.; Abe, H. Effect of coenzyme Q7 tretment on blood sugar and ketone bodies of diabetes. J. Vitaminol. 1966, 12, 293–298. [Google Scholar] [CrossRef]
- Shimura, Y.; Hogimoto, S. Significance of coenzyme Q10 on the treatment of diabetes mellitus. Jpn. J. Clin. Exp. Med. 1981, 58, 1349–1532. [Google Scholar]
- Werbach, M.R. Nutritional Influences on Illness; Third Line Press: Tarzana, CA, USA, 1993. [Google Scholar]
- Conget, I.; Manzanares, J.M.; Barrientos, A.; Cardellach, F.; Gomis, R. Coenzyme Q10 and insulin secretion in vitro. Diabetes Res. Clin. Pract. 1996, 33, 135–136. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, J.E.; Andersen, C.B.; Hother-Nielsen, O.; Vaag, A.; Mortensen, S.A.; Beck-Nielsen, H. Impact of ubiquinone (coenzyme Q10) treatment on glycaemic control, insulin requirement and well-being in patients with type 1 diabetes mellitus. Diabet. Med. 1999, 16, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.B.; Niaz, M.A.; Rastogi, S.S.; Shukla, P.K.; Thakur, A.S. Effect of hydrosoluble coenzyme Q10 on blood pressures and insulin resistance in hypertensive patients with coronary artery disease. J. Hum. Hypertens. 1999, 13, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, J.M.; Watts, G.F.; Playford, D.A.; Burke, V.; Croft, K.D. Coenzyme Q10 improves blood pressure and glycaemic control: A controlled trial in subjects with type 2 diabetes. Eur. J. Clin. Nutr. 2002, 56, 1137–1142. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.C.; Lekshminarayanan, R.; Goh, S.K.; Ong, Y.Y.; Subramaniam, T.; Sum, C.F.; Ong, C.N.; Lee, B.L. The effect of coenzyme Q10 on microcirculatory endothelial function of subjects with type 2 diabetes mellitus. Atherosclerosis 2008, 196, 966–969. [Google Scholar] [CrossRef] [PubMed]
- Sena, C.M.; Nunes, E.; Gomes, A.; Santos, M.S.; Proenca, T.; Martins, M.I.; Seica, R.M. Supplementation of coenzyme Q10 and alpha-tocopherol lowers glycated hemoglobin level and lipid peroxidation in pancreas of diabetic rats. Nutr. Res. 2008, 28, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Kolahdouz Mohammadi, R.; Hosseinzadeh-Attar, M.J.; Eshraghian, M.R.; Nakhjavani, M.; Khorami, E.; Esteghamati, A. The effect of coenzyme Q10 supplementation on metabolic status of type 2 diabetic patients. Minerva Gastroenterol. Dietol. 2013, 59, 231–236. [Google Scholar] [PubMed]
- Yokoyama, H.; Lingle, D.M.; Crestanello, J.A.; Kamelgard, J.; Kott, B.R.; Momeni, R.; Millili, J.; Mortensen, S.A.; Whitman, G.J. Coenzyme Q10 protects coronary endothelial function from ischemia reperfusion injury via an antioxidant effect. Surgery 1996, 120, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, Y.; Liu, Y.; Sun, R.; Xia, M. Purified anthocyanin supplementation reduces dyslipidemia, enhances antioxidant capacity, and prevents insulin resistance in diabetic patients. J. Nutr. 2015, 145, 742–748. [Google Scholar] [CrossRef] [PubMed]
- Rafighi, Z.; Shiva, A.; Arab, S.; Mohd Yousof, R. Association of dietary vitamin C and E intake and antioxidant enzymes in type 2 diabetes mellitus patients. Glob. J. Health Sci. 2013, 5, 183–187. [Google Scholar] [PubMed]
- Bhatt, J.K.; Thomas, S.; Nanjan, M.J. Resveratrol supplementation improves glycemic control in type 2 diabetes mellitus. Nutr. Res. 2012, 32, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Movahed, A.; Nabipour, I.; Lieben Louis, X.; Thandapilly, S.J.; Yu, L.; Kalantarhormozi, M.; Rekabpour, S.J.; Netticadan, T. Antihyperglycemic effects of short term resveratrol supplementation in type 2 diabetic patients. Evid. Based Complement. Altern. Med. 2013. [Google Scholar] [CrossRef]
- Han, J.; Song, B.; Kim, J.; Kodali, V.K.; Pottekat, A.; Wang, M.; Hassler, J.; Wang, S.; Pennathur, S.; Back, S.H.; et al. Antioxidants complement the requirement for protein chaperone function to maintain beta cell function and glucose homeostasis. Diabetes 2015. [Google Scholar] [CrossRef]
- Crandall, J.P.; Oram, V.; Trandafirescu, G.; Reid, M.; Kishore, P.; Hawkins, M.; Cohen, H.W.; Barzilai, N. Pilot study of resveratrol in older adults with impaired glucose tolerance. J. Gerontol. A Biol. Sci. Med. Sci. 2012, 67, 1307–1312. [Google Scholar] [CrossRef] [PubMed]
- Kataja-Tuomola, M.K.; Kontto, J.P.; Mannisto, S.; Albanes, D.; Virtamo, J. Intake of antioxidants and risk of type 2 diabetes in a cohort of male smokers. Eur. J. Clin. Nutr. 2011, 65, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Parham, M.; Amini, M.; Aminorroaya, A.; Heidarian, E. Effect of zinc supplementation on microalbuminuria in patients with type 2 diabetes: A double blind, randomized, placebo-controlled, cross-over trial. Rev. Diabet. Stud. 2008, 5, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Seet, R.C.; Lee, C.Y.; Lim, E.C.; Quek, A.M.; Huang, H.; Huang, S.H.; Looi, W.F.; Long, L.H.; Halliwell, B. Oral zinc supplementation does not improve oxidative stress or vascular function in patients with type 2 diabetes with normal zinc levels. Atherosclerosis 2011, 219, 231–239. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, Q.; Pierce, J.D. Supplementation of Coenzyme Q10 among Patients with Type 2 Diabetes Mellitus. Healthcare 2015, 3, 296-309. https://doi.org/10.3390/healthcare3020296
Shen Q, Pierce JD. Supplementation of Coenzyme Q10 among Patients with Type 2 Diabetes Mellitus. Healthcare. 2015; 3(2):296-309. https://doi.org/10.3390/healthcare3020296
Chicago/Turabian StyleShen, Qiuhua, and Janet D. Pierce. 2015. "Supplementation of Coenzyme Q10 among Patients with Type 2 Diabetes Mellitus" Healthcare 3, no. 2: 296-309. https://doi.org/10.3390/healthcare3020296