
Comparison of Activation in the Prefrontal Cortex of Native Speakers of Mandarin by Ability of Japanese as a Second Language Using a Novel Speaking Task



Abstract
:1. Introduction
2. Methods
2.1. Subjects
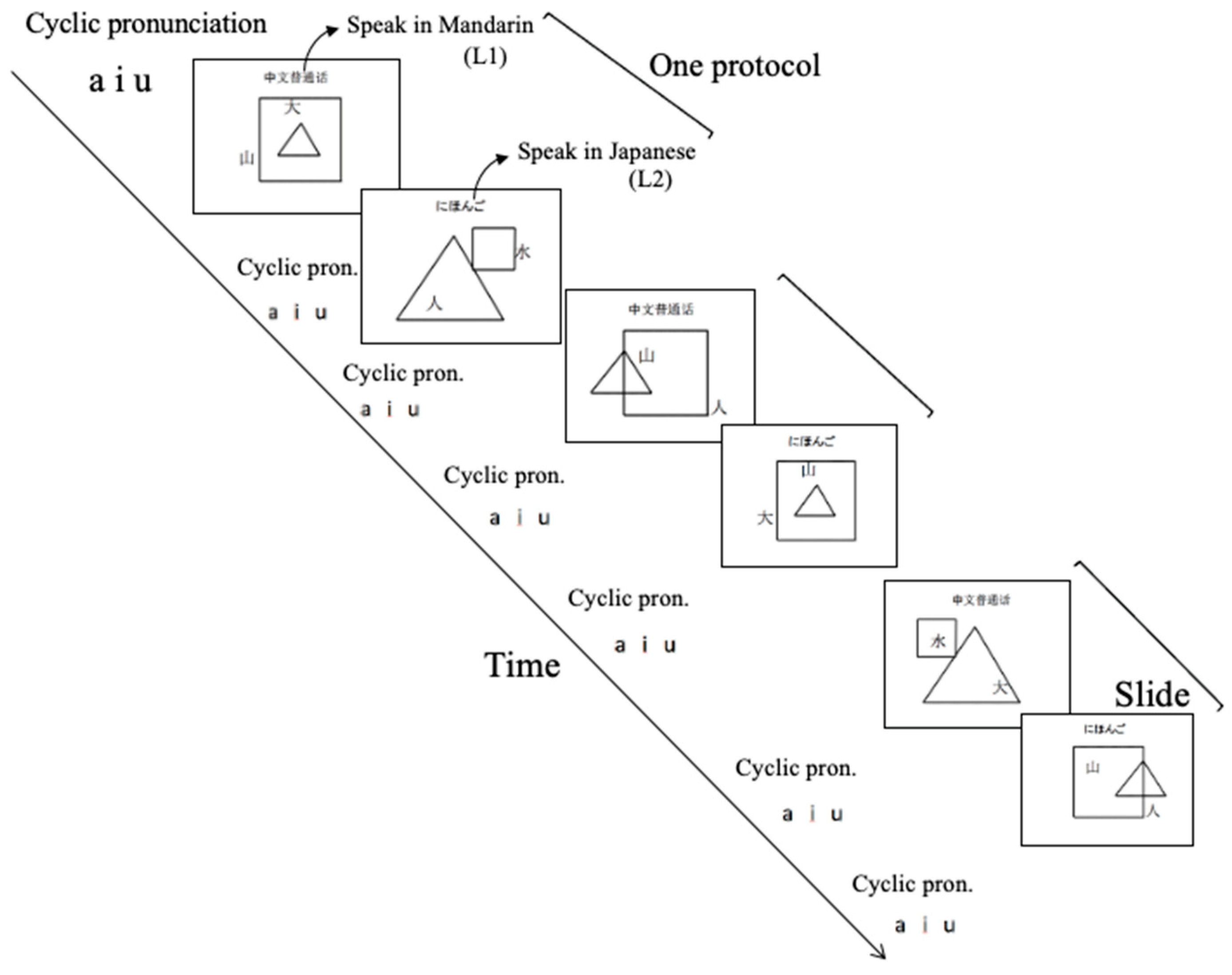
2.2. Speaking Task
2.3. Measurement Environment
2.4. Measuring Positions
2.5. Data Analysis
2.5.1. Approximate Integrals of Cerebral Blood Flow Change
2.5.2. Common Activation Regions
3. Results
3.1. Language Proficiency
3.2. Cerebral Blood Flow Change in the Prefrontal Cortex
3.3. Cerebral Blood Flow Change in the Left and Right Prefrontal Cortices
3.4. Cerebral Blood Flow Change at Each of the 22 Channels
3.5. Correlations between Cerebral Blood Flow Change and Language Proficiency
3.6. Common Activation Area Obtained from NIRS-SPM Analysis
4. Discussion
4.1. Subjects Selection and Cognitive Reserve Unification
4.2. Validity of Experimental Conditions and Analysis Methods
4.3. Comparison of Brain Activation
4.4. Mutual Influence of Language Distance
4.5. Study Limitations and Prospects
5. Conclusions
- People with low L2 ability showed much more brain activation when speaking L2 than when speaking L1. People with high L2 ability showed high-level brain activation when speaking either L2 or L1. Almost the same high-level brain activation was observed in both ability groups when speaking L2.
- The high level of activation in people with high L2 ability when speaking either L2 or L1 suggested strong inhibition of the non-spoken language. A wider area of brain activation in people with low compared with high L2 ability when speaking L2 is considered to be attributed to the cognitive load involved in code-switching L1 to L2 with strong inhibition of L1 and the cognitive load involved in using L2.
- The above results suggest that learning a second language of Japanese would be helpful for Chinese speakers of Mandarin to delay the onset of dementia by changing brain activation pattern. This effect should also be furtherly confirmed through an analysis of a wider area of the brain of more subjects using the fNIRS measurement as well as other techniques. Furthermore, implications for the fields of neurolinguistics and language education are also expected. An effective method for language education in enhancing the cognitive function might be important.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sakai, K.L. Language acquisition and brain development. Science 2005, 310, 815–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoniou, M.; Gunasekera, G.M.; Wong, P.C. Foreign language training as cognitive therapy for age-related cognitive decline: A hypothesis for future research. Neurosci. Biobehav. Rev. 2013, 37 Pt 2, 2689–2698. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Fornells, A.; Cunillera, T.; Mestres-Misse, A.; de Diego-Balaguer, R. Neurophysiological mechanisms involved in language learning in adults. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 3711–3735. [Google Scholar] [CrossRef] [Green Version]
- Hosoda, C.; Tanaka, K.; Nariai, T.; Honda, M.; Hanakawa, T. Dynamic neural network reorganization associated with second language vocabulary acquisition: A multimodal imaging study. J. Neurosci. 2013, 33, 13663–13672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legault, J.; Fang, S.Y.; Lan, Y.J.; Li, P. Structural brain changes as a function of second language vocabulary training: Effects of learning context. Brain Cogn. 2018, 134, 90–102. [Google Scholar] [CrossRef]
- Del Maschio, N.; Sulpizio, S.; Gallo, F.; Fedeli, D.; Weekes, B.S.; Abutalebi, J. Neuroplasticity across the lifespan and aging effects in bilinguals and monolinguals. Brain Cogn. 2018, 125, 118–126. [Google Scholar] [CrossRef]
- Li, P.; Legault, J.; Litcofsky, K.A. Neuroplasticity as a function of second language learning: Anatomical changes in the human brain. Cortex 2014, 58, 301–324. [Google Scholar] [CrossRef]
- Alladi, S.; Bak, T.H.; Duggirala, V.; Surampudi, B.; Shailaja, M.; Shukla, A.K.; Chaudhuri, J.R.; Kaul, S. Bilingualism delays age at onset of dementia, independent of education and immigration status. Neurology 2013, 81, 1938–1944. [Google Scholar] [CrossRef] [PubMed]
- Rose, N.S.; Luo, L.; Bialystok, E.; Hering, A.; Lau, K.; Craik, F.I. Cognitive processes in the Breakfast Task: Planning and monitoring. Can. J. Exp. Psychol. 2015, 69, 252–263. [Google Scholar] [CrossRef] [Green Version]
- Schweizer, T.A.; Ware, J.; Fischer, C.E.; Craik, F.I.; Bialystok, E. Bilingualism as a contributor to cognitive reserve: Evidence from brain atrophy in Alzheimer’s disease. Cortex 2012, 48, 991–996. [Google Scholar] [CrossRef] [PubMed]
- Ardila, A.; Bernal, B.; Rosselli, M. Connectivity of BA46 involvement in the executive control of language. Psicothema 2016, 28, 26–31. [Google Scholar] [CrossRef]
- De Baene, W.; Duyck, W.; Brass, M.; Carreiras, M. Brain Circuit for Cognitive Control is Shared by Task and Language Switching. J. Cogn. Neurosci. 2015, 27, 1752–1765. [Google Scholar] [CrossRef] [Green Version]
- Craik, F.I.M. Cognitive Problems in Older Adults: Can Bilingualism Help? Chapter 2; John Benjamins Publishing Company: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Green, D.W.; Abutalebi, J. Language control in bilinguals: The adaptive control hypothesis. J. Cogn. Psychol. 2013, 25, 515–530. [Google Scholar] [CrossRef]
- Nickels, L.; Hameau, S.; Nair, V.K.K.; Barr, P.; Biedermann, B. Ageing with bilingualism: Benefits and challenges. Speech Lang. Hear. 2019, 22, 32–50. [Google Scholar] [CrossRef]
- Li, P.; Sepanski, S.; Zhao, X. Language history questionnaire: A web-based interface for bilingual research. Behav. Res. Methods 2006, 38, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Calabria, M.; Branzi, F.M.; Marne, P.; HernÁNdez, M.; Costa, A. Age-related effects over bilingual language control and executive control. Biling. Lang. Cogn. 2013, 18, 65–78. [Google Scholar] [CrossRef]
- Borragan, M.; Martin, C.D.; de Bruin, A.; Dunabeitia, J.A. Exploring Different Types of Inhibition During Bilingual Language Production. Front. Psychol. 2018, 9, 2256. [Google Scholar] [CrossRef]
- Nucci, M.; Mapelli, D.; Mondini, S. Cognitive Reserve Index questionnaire (CRIq): A new instrument for measuring cognitive reserve. Aging Clin. Exp. Res. 2012, 24, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Endo, K.; Liang, N.; Idesako, M.; Ishii, K.; Matsukawa, K. Incremental rate of prefrontal oxygenation determines performance speed during cognitive Stroop test: The effect of ageing. J. Physiol. Sci. 2018, 68, 807–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamashiro, H.; Kinoshita, S.; Okamoto, T.; Urushidani, N.; Abo, M. Effect of baseline brain activity on response to low-frequency rTMS/intensive occupational therapy in poststroke patients with upper limb hemiparesis: A near-infrared spectroscopy study. Int. J. Neurosci. 2018, 129, 337–343. [Google Scholar] [CrossRef]
- Kondo, A.; Shoji, Y.; Morita, K.; Sato, M.; Ishii, Y.; Yanagimoto, H.; Nakano, S.; Uchimura, N. Characteristics of oxygenated hemoglobin concentration change during pleasant and unpleasant image-recall tasks in patients with depression: Comparison with healthy subjects. Psychiatry Clin. Neurosci. 2018, 72, 611–622. [Google Scholar] [CrossRef]
- Ye, J.C.; Tak, S.; Jang, K.E.; Jung, J.; Jang, J. NIRS-SPM: Statistical Parametric Mapping for Near-infrared Spectroscopy. Neuroimage 2011, 44, 428–447. [Google Scholar] [CrossRef] [PubMed]
- García, A.M.; Muñoz, E.; Kogan, B. Taxing the bilingual mind: Effects of simultaneous interpreting experience on verbal and executive mechanisms. Biling. Lang. Cogn. 2019, 23, 729–739. [Google Scholar] [CrossRef]
- Klein, D.; Mok, K.; Chen, J.K.; Watkins, K.E. Age of language learning shapes brain structure: A cortical thickness study of bilingual and monolingual individuals. Brain Lang. 2014, 131, 20–24. [Google Scholar] [CrossRef]
- Mohades, S.G.; Struys, E.; Van Schuerbeek, P.; Mondt, K.; Van De Craen, P.; Luypaert, R. DTI reveals structural differences in white matter tracts between bilingual and monolingual children. Brain Res. 2012, 1435, 72–80. [Google Scholar] [CrossRef]
- Kakimoto, Y.; Nishimura, Y.; Hara, N.; Okada, M.; Tanii, H.; Okazaki, Y. Intrasubject reproducibility of prefrontal cortex activities during a verbal fluency task over two repeated sessions using multi-channel near-infrared spectroscopy. Psychiatry Clin. Neurosci. 2009, 63, 491–499. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, A.W.; Cohen, J.D.; Stenger, V.A.; Carter, C.S. Dissociating the Role of the Dorsolateral Prefrontal and Anterior Cingulate Cortex in Cognitive Control. Science 2000, 288, 1835–1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klaus, J.; Schutter, D. The Role of Left Dorsolateral Prefrontal Cortex in Language Processing. Neuroscience 2018, 377, 197–205. [Google Scholar] [CrossRef]
- Grundy, J.G.; Timmer, K. Bilingualism and working memory capacity: A comprehensive meta-analysis. Second Lang. Res. 2016, 33, 325–340. [Google Scholar] [CrossRef]
- Alvarez, J.A.; Emory, E. Executive function and the frontal lobes: A meta-analytic review. Neuropsychol. Rev. 2006, 16, 17–42. [Google Scholar] [CrossRef]
- Godefroy, O.; Jeannerod, M.; Allain, P.; Le Gall, D. Lobe frontal, fonctions exécutives et controle cognitif Frontal lobe, executive functions and cognitive contro. Rev. Neuroloque 2018, 164, 119–127. [Google Scholar] [CrossRef]
- Forstmann, B.U.; Jahfari, S.; Scholte, H.S.; Wolfensteller, U.; van den Wildenberg, W.P.; Ridderinkhof, K.R. Function and structure of the right inferior frontal cortex predict individual differences in response inhibition: A model-based approach. J. Neurosci. 2008, 28, 9790–9796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Ettinger-Veenstra, H.; Ragnehed, M.; McAllister, A.; Lundberg, P.; Engstrom, M. Right-hemispheric cortical contributions to language ability in healthy adults. Brain Lang. 2012, 120, 395–400. [Google Scholar] [CrossRef] [Green Version]
- Lei, M.; Akama, H.; Murphy, B. Neural basis of language switching in the brain: fMRI evidence from Korean-Chinese early bilinguals. Brain Lang. 2014, 138, 12–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita, S.; Tamashiro, H.; Okamoto, T.; Urushidani, N.; Abo, M. Association between imbalance of cortical brain activity and successful motor recovery in sub-acute stroke patients with upper limb hemiparesis: A functional near-infrared spectroscopy study. Neuroreport 2019, 30, 822–827. [Google Scholar] [CrossRef] [PubMed]
- Chiswick, B.R.; Miller, P.W. Linguistic Distance: A Quantitative Measure of the Distance between English and Other Languages. J. Multiling. Multicult. Dev. 2005, 26, 1–11. [Google Scholar] [CrossRef]
- Ghazi-Saidi, L.; Ansaldo, A.I. Second Language Word Learning through Repetition and Imitation: Functional Networks as a Function of Learning Phase and Language Distance. Front. Hum. Neurosci. 2017, 11, 463. [Google Scholar] [CrossRef] [Green Version]
- Costa, A.; Santesteban, M. Lexical access in bilingual speech production: Evidence from language switching in highly proficient bilinguals and L2 learners. J. Mem. Lang. 2004, 50, 491–511. [Google Scholar] [CrossRef]
- Linck, J.A.; Kroll, J.F.; Sunderman, G. Losing access to the native language while immersed in a second language: Evidence for the role of inhibition in second-language learning. Psychol. Sci. 2009, 20, 1507–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, M.D.; Janus, M.; Moreno, S.; Astheimer, L.; Bialystok, E. Early stage second-language learning improves executive control: Evidence from ERP. Brain Lang. 2014, 139, 84–98. [Google Scholar] [CrossRef]
- Jasinska, K.K.; Petitto, L.A. Development of neural systems for reading in the monolingual and bilingual brain: New insights from functional near infrared spectroscopy neuroimaging. Dev. Neuropsychol. 2014, 39, 421–439. [Google Scholar] [CrossRef]
- Peter Bright, R.F. Perspectives on the ‘Bilingual Advantage’: Challenges and Opportunities. Front. Psychol. 2019, 10, 1346. [Google Scholar] [CrossRef] [PubMed]
- Dash, T.; Ghazi-Saidi, L.; Berroir, P.; Adrover-Roig, D.; Benali, H.; Ansaldo, A.I. Is the bilingual brain better equipped for aging. OLBI J. 2017, 8. [Google Scholar] [CrossRef]
- Stern, Y. What is cognitive reserve? Theory and research application of the reserve concept. J. Int. Neuropsychol. Soc. 2002, 8, 448–460. [Google Scholar] [CrossRef] [PubMed]
- Stern, Y. Cognitive reserve. Neuropsychologia 2009, 47, 2015–2028. [Google Scholar] [CrossRef]
- Craik, F.I.; Bialystok, E.; Freedman, M. Delaying the onset of Alzheimer disease: Bilingualism as a form of cognitive reserve. Neurology 2010, 75, 1726–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzman-Velez, E.; Tranel, D. Does bilingualism contribute to cognitive reserve? Cognitive and neural perspectives. Neuropsychology 2015, 29, 139–150. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| All Subjects Are Native Speakers of Mandarin (L1) with Japanese as a Second Language (L2) | ||||
|---|---|---|---|---|
| Characteristics | Group 1 (n = 12) | Group 2 (n = 12) | p-Value | |
| Age (years: mean ± SD) | 51.1 ± 5.0 | 24.9 ± 1.4 | <0.0001 | |
| Sex (female/male) | 5/7 | 6/6 | ||
| Living years in Japan (years: mean ± SD) | 22.3 ± 3.5 | 2.75 ± 1.1 | <0.0001 | |
| AOA * (years: mean ± SD) | 27.3 ± 2.5 | 22.4 ± 0.5 | <0.0001 | |
| Japanese (L2) | Reading (mean score ± SD) | 9.2 ± 0.4 | 4.2 ± 1.8 | <0.0001 |
| Listening (mean score ± SD) | 9.2 ± 0.7 | 4.0 ± 1.8 | <0.0001 | |
| Writing (mean score ± SD) | 8.1 ± 1.2 | 2.8 ± 1.8 | <0.0001 | |
| Speaking (mean score ± SD) | 8.5 ± 1.2 | 3.4 ± 1.8 | <0.0001 | |
| total-Japanese (mean score ± SD) | 8.8 ± 0.7 | 3.6 ± 1.7 | <0.0001 | |
| Mandarin (L1) | Reading (mean score ± SD) | 9.4 ± 0.9 | 9.3 ± 0.9 | =0.8215 |
| Listening (mean score ± SD) | 9.5 ± 0.7 | 9.8 ± 0.4 | =0.150 | |
| Writing (mean score ± SD) | 9.0 ± 0.6 | 9.0 ± 1.1 | =1.0000 | |
| Speaking (mean score ± SD) | 9.5 ± 0.5 | 9.7 ± 0.5 | =0.3140 | |
| total-Japanese (mean score ± SD) | 9.4 ± 0.5 | 9.5 ± 0.6 | =0.6553 | |
| Criq ** | CRiq-E *** (mean score ± SD) | 132 ± 2 | 102 ± 5 | <0.0001 |
| CRiq-W **** (mean score ± SD) | 108 ± 15 | 91 ± 1 | =0.0006 | |
| CRiq-L ***** (mean score ± SD) | 107 ± 8 | 89 ± 1 | <0.0001 | |
| total-CRiq (mean score ± SD) | 120 ± 8 | 92 ± 3 | <0.0001 | |
| group 1 = High L2 ability group 2 = Low L2 ability | ||||
| L1 Task | L2 Task | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| High-L2-Ability Group | Low-L2-Ability Group | High-L2-Ability Group | Low-L2-Ability Group | |||||||
| Channel | Mean | SD | Mean | SD | p-Value | Mean | SD | Mean | SD | p-Value |
| ch1 | 0.739 | 0.621 | −0.0570 | 0.705 | 0.0077 | 0.641 | 0.863 | 0.837 | 0.855 | 0.5819 |
| ch2 | 0.606 | 0.636 | −0.1490 | 0.660 | 0.0093 | 0.532 | 0.946 | 0.414 | 0.843 | 0.7508 |
| ch3 | 0.611 | 0.619 | 0.1130 | 0.569 | 0.0517 | 0.585 | 0.806 | 0.537 | 0.879 | 0.8892 |
| ch4 | 0.840 | 1.065 | 0.1810 | 0.719 | 0.0894 | 0.748 | 1.370 | 0.936 | 1.150 | 0.719 |
| ch5 | 0.876 | 0.659 | −0.2160 | 0.818 | 0.0016 | 0.699 | 0.833 | 0.698 | 1.180 | 0.9984 |
| ch6 | 0.516 | 0.653 | −0.4860 | 0.772 | 0.0024 | 0.318 | 0.100 | 0.350 | 1.004 | 0.9376 |
| ch7 | 0.487 | 0.507 | −0.0930 | 0.581 | 0.0162 | 0.327 | 0.801 | 0.460 | 0.775 | 0.6841 |
| ch8 | 0.663 | 0.725 | 0.2160 | 0.649 | 0.1258 | 0.508 | 1.081 | 0.827 | 0.820 | 0.4248 |
| ch9 | 1.094 | 0.885 | 0.4080 | 0.588 | 0.0358 | 0.729 | 1.246 | 1.252 | 0.976 | 0.265 |
| ch10 | 0.621 | 0.664 | −0.5850 | 0.732 | 0.0003 | 0.364 | 0.653 | 0.442 | 1.192 | 0.8451 |
| ch11 | 0.618 | 0.459 | −0.2250 | 0.533 | 0.0004 | 0.422 | 0.598 | 0.680 | 0.936 | 0.4287 |
| ch12 | 0.672 | 0.520 | 0.0580 | 0.529 | 0.009 | 0.630 | 0.664 | 0.692 | 0.725 | 0.829 |
| ch13 | 1.132 | 0.674 | 0.3490 | 0.685 | 0.0099 | 0.894 | 0.290 | 1.420 | 0.290 | 0.2126 |
| ch14 | 0.851 | 0.804 | 0.0500 | 0.427 | 0.0059 | 0.528 | 0.587 | 0.745 | 1.267 | 0.5944 |
| ch15 | 0.769 | 0.927 | −0.4180 | 0.492 | 0.0007 | 0.539 | 1.007 | 0.992 | 1.262 | 0.3415 |
| ch16 | 1.351 | 1.081 | −0.1440 | 0.984 | 0.0018 | 1.102 | 1.105 | 1.267 | 1.064 | 0.7129 |
| ch17 | 1.070 | 0.853 | −0.0160 | 0.480 | 0.0009 | 0.845 | 0.953 | 1.073 | 1.059 | 0.5858 |
| ch18 | 1.005 | 0.703 | 0.2550 | 0.575 | 0.0091 | 0.956 | 0.775 | 1.158 | 1.002 | 0.5856 |
| ch19 | 1.135 | 0.867 | −0.1270 | 0.909 | 0.0021 | 1.056 | 1.146 | 0.749 | 0.841 | 0.4626 |
| ch20 | 1.200 | 1.074 | −0.2870 | 1.024 | 0.0022 | 1.130 | 1.094 | 0.693 | 1.027 | 0.3242 |
| ch21 | 1.201 | 1.199 | −0.2430 | 0.869 | 0.0027 | 1.221 | 1.044 | 0.784 | 1.079 | 0.3244 |
| ch22 | 0.951 | 0.757 | 0.0020 | 0.711 | 0.0044 | 1.119 | 0.980 | 0.712 | 1.038 | 0.3339 |
| Total right | 7.932 | 6.379 | −2.4990 | 4.791 | 0.0002 | 6.229 | 7.453 | 6.602 | 8.604 | 0.9106 |
| Total left | 9.239 | 6.990 | 1.3200 | 4.209 | 0.0028 | 8.236 | 8.480 | 9.391 | 7.542 | 0.7277 |
| L1 Task | L2 Task | ||||
|---|---|---|---|---|---|
| Channel | Mean | SD | Mean | SD | p-Value |
| ch1 | 0.739 | 0.621 | 0.641 | 0.863 | 0.7526 |
| ch2 | 0.606 | 0.636 | 0.532 | 0.946 | 0.8248 |
| ch3 | 0.611 | 0.619 | 0.585 | 0.806 | 0.9310 |
| ch4 | 0.840 | 1.065 | 0.748 | 1.370 | 0.8555 |
| ch5 | 0.876 | 0.659 | 0.699 | 0.833 | 0.5693 |
| ch6 | 0.516 | 0.653 | 0.318 | 0.100 | 0.5706 |
| ch7 | 0.487 | 0.507 | 0.327 | 0.801 | 0.5664 |
| ch8 | 0.663 | 0.725 | 0.508 | 1.081 | 0.6850 |
| ch9 | 1.094 | 0.885 | 0.729 | 1.246 | 0.4180 |
| ch10 | 0.621 | 0.664 | 0.364 | 0.653 | 0.3492 |
| ch11 | 0.618 | 0.459 | 0.422 | 0.598 | 0.3780 |
| ch12 | 0.672 | 0.520 | 0.630 | 0.664 | 0.8660 |
| ch13 | 1.132 | 0.674 | 0.894 | 0.290 | 0.4874 |
| ch14 | 0.851 | 0.804 | 0.528 | 0.587 | 0.2721 |
| ch15 | 0.769 | 0.927 | 0.539 | 1.007 | 0.5659 |
| ch16 | 1.351 | 1.081 | 1.102 | 1.105 | 0.5832 |
| ch17 | 1.07 | 0.853 | 0.845 | 0.953 | 0.5486 |
| ch18 | 1.005 | 0.703 | 0.956 | 0.775 | 0.8723 |
| ch19 | 1.135 | 0.867 | 1.056 | 1.146 | 0.8506 |
| ch20 | 1.200 | 1.074 | 1.130 | 1.094 | 0.8769 |
| ch21 | 1.201 | 1.199 | 1.221 | 1.044 | 0.9648 |
| ch22 | 0.951 | 0.757 | 1.119 | 0.98 | 0.6439 |
| Total right | 7.932 | 6.379 | 6.229 | 7.453 | 0.5539 |
| Total left | 9.239 | 6.990 | 8.236 | 8.480 | 0.7549 |
| L1 Task | L2 Task | ||||
|---|---|---|---|---|---|
| Channel | Mean | SD | Mean | SD | p-Value |
| ch1 | −0.057 | 0.705 | 0.837 | 0.855 | 0.0106 |
| ch2 | −0.149 | 0.660 | 0.414 | 0.843 | 0.0821 |
| ch3 | 0.113 | 0.569 | 0.537 | 0.879 | 0.1742 |
| ch4 | 0.181 | 0.719 | 0.936 | 1.150 | 0.0667 |
| ch5 | −0.216 | 0.818 | 0.698 | 1.180 | 0.0381 |
| ch6 | −0.486 | 0.772 | 0.350 | 1.004 | 0.0322 |
| ch7 | −0.093 | 0.581 | 0.460 | 0.775 | 0.0605 |
| ch8 | 0.216 | 0.649 | 0.827 | 0.820 | 0.0552 |
| ch9 | 0.408 | 0.588 | 1.252 | 0.976 | 0.0176 |
| ch10 | −0.585 | 0.732 | 0.442 | 1.192 | 0.0185 |
| ch11 | −0.225 | 0.533 | 0.680 | 0.936 | 0.0081 |
| ch12 | 0.058 | 0.529 | 0.692 | 0.725 | 0.0227 |
| ch13 | 0.349 | 0.685 | 1.420 | 0.290 | 0.0073 |
| ch14 | 0.050 | 0.427 | 0.745 | 1.267 | 0.0855 |
| ch15 | −0.418 | 0.492 | 0.992 | 1.262 | 0.0016 |
| ch16 | −0.144 | 0.984 | 1.267 | 1.064 | 0.0027 |
| ch17 | −0.016 | 0.480 | 1.073 | 1.059 | 0.0037 |
| ch18 | 0.255 | 0.575 | 1.158 | 1.002 | 0.0013 |
| ch19 | −0.127 | 0.909 | 0.749 | 0.841 | 0.0266 |
| ch20 | −0.287 | 1.024 | 0.693 | 1.027 | 0.0287 |
| ch21 | −0.243 | 0.869 | 0.784 | 1.079 | 0.0175 |
| ch22 | 0.002 | 0.711 | 0.712 | 1.038 | 0.0630 |
| Total right | −2.499 | 4.791 | 6.602 | 8.604 | 0.0041 |
| Total left | 1.320 | 4.209 | 9.391 | 7.542 | 0.0038 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cong, L.; Miyaguchi, H.; Ishizuki, C. Comparison of Activation in the Prefrontal Cortex of Native Speakers of Mandarin by Ability of Japanese as a Second Language Using a Novel Speaking Task. Healthcare 2021, 9, 412. https://doi.org/10.3390/healthcare9040412
Cong L, Miyaguchi H, Ishizuki C. Comparison of Activation in the Prefrontal Cortex of Native Speakers of Mandarin by Ability of Japanese as a Second Language Using a Novel Speaking Task. Healthcare. 2021; 9(4):412. https://doi.org/10.3390/healthcare9040412
Chicago/Turabian StyleCong, Li, Hideki Miyaguchi, and Chinami Ishizuki. 2021. "Comparison of Activation in the Prefrontal Cortex of Native Speakers of Mandarin by Ability of Japanese as a Second Language Using a Novel Speaking Task" Healthcare 9, no. 4: 412. https://doi.org/10.3390/healthcare9040412