
Molecular and Cellular Markers in Chlorhexidine-Induced Peritoneal Fibrosis in Mice
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Permission
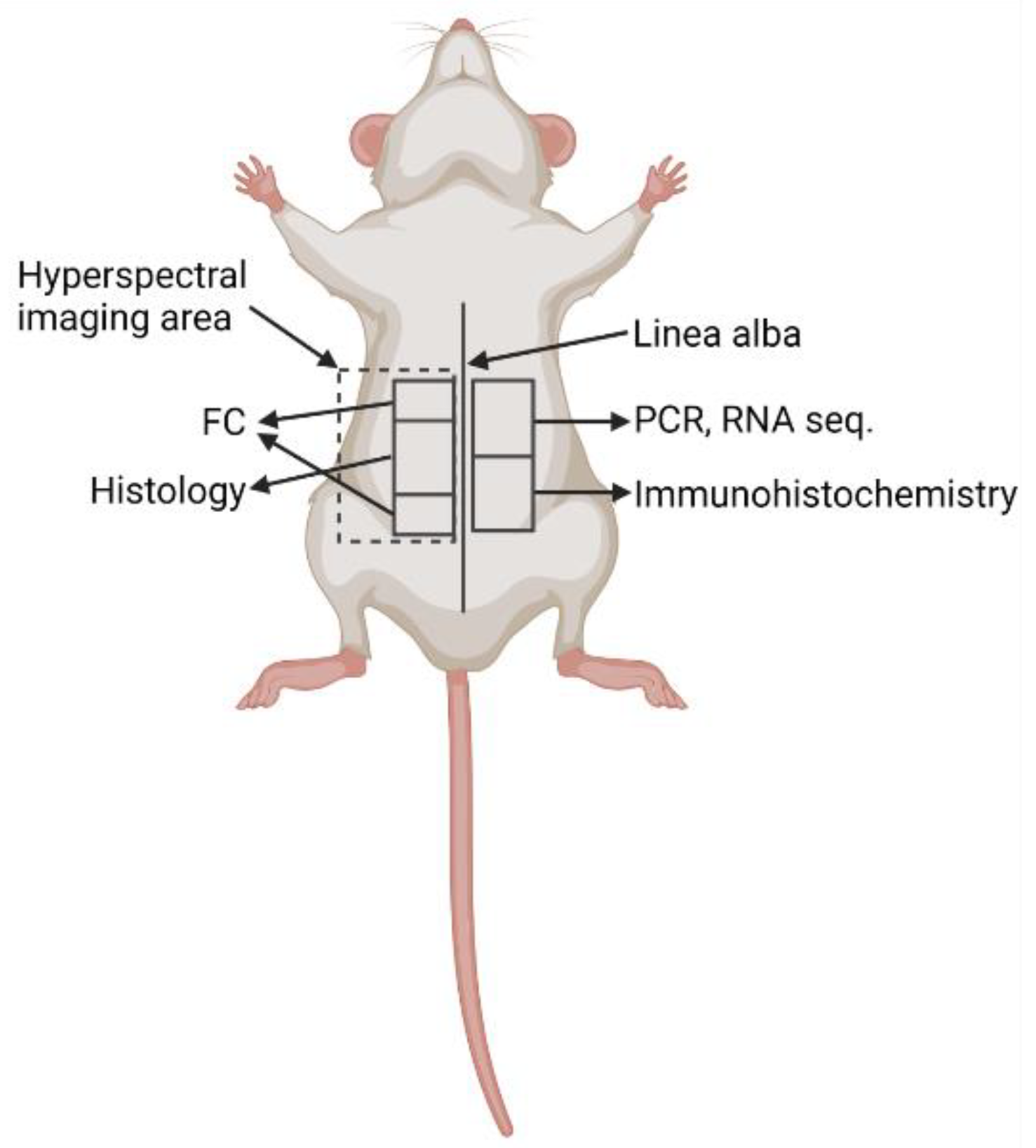
2.2. Animals and Experimental Design
2.3. Induction of Peritonitis/Peritoneal Fibrosis
2.4. Collection of Biological Material
2.5. Flow Cytometry
2.6. Histology
2.7. Immunohistochemistry
2.8. RNA Isolation
2.9. RNA Sequencing
2.10. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.11. Enzyme-Linked Immunosorbent Assay (ELISA)
2.12. Luminex Assay
2.13. Hyperspectral Imaging
2.14. Statistical Analysis
3. Results
3.1. CHX-Treated Mice Show Abdominal Pain Behavior and No Obvious Macroscopicaly Visible Changes in the Abdomen after 1 Week of Treatment
3.2. Histological Changes in Parietal and Visceral Peritoneum Start Localy and Expand after Continious Stimulus
3.3. Use of Novel Sensitive Techniques to Evaluate Tissue Damage
3.4. Inflammation in CHX-Treated Mice Is Present Locally, with Increased Polymorphonuclear Cells in the Blood
3.5. CHX-Induced Peritoneal Fibrosis Is Characterized by the Increased Expression of Extracellular Matrix Genes and Genes That Regulate Extracellular Matrix
3.6. RNA Sequencing of the Abdominal Wall Reveals High Heterogenity between Intragroup Experimental Subjects and Points out Novel Gene Expression Pathways That Change with Continous CHX Treatment
3.7. Local Expression of the Components of the Complement Cascade in the Abdominal Wall of CHX-Induced Model Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Terri, M.; Trionfetti, F.; Montaldo, C.; Cordani, M.; Tripodi, M.; Lopez-Cabrera, M.; Strippoli, R. Mechanisms of Peritoneal Fibrosis: Focus on Immune Cells–Peritoneal Stroma Interactions. Front. Immunol. 2021, 12, 607204. [Google Scholar] [CrossRef] [PubMed]
- Junor, B.J.R.; Briggs, J.D.; Forwell, M.A.; Dobbie, J.W.; Henderson, I. Sclerosing Peritonitis—The Contribution of Chlorhexidine in Alcohol. Perit. Dial. Int. 1985, 5, 101–104. [Google Scholar] [CrossRef]
- Suga, H.; Teraoka, S.; Ota, K.; Komemushi, S.; Furutani, S.; Yamauchi, S.; Margolin, S. Preventive effect of pirfenidone against experimental sclerosing peritonitis in rats. Exp. Toxicol. Pathol. 1995, 47, 287–291. [Google Scholar] [CrossRef]
- Ishii, Y.; Sawada, T.; Shimizu, A.; Tojimbara, T.; Nakajima, I.; Fuchinoue, S.; Teraoka, S. An experimental sclerosing encapsulating peritonitis model in mice. Nephrol. Dial. Transplant. 2001, 16, 1262–1266. [Google Scholar] [CrossRef] [PubMed]
- Nishino, T.; Ashida, R.; Obata, Y.; Furusu, A.; Abe, K.; Miyazaki, M.; Koji, T.; Kohno, S. Involvement of Lymphocyte Infiltration in the Progression of Mouse Peritoneal Fibrosis Model. Ren. Fail. 2012, 34, 760–766. [Google Scholar] [CrossRef]
- Yoshio, Y.; Miyazaki, M.; Abe, K.; Nishino, T.; Furusu, A.; Mizuta, Y.; Harada, T.; Ozono, Y.; Koji, T.; Kohno, S. TNP-470, An angiogenesis inhibitor, suppresses the progression of peritoneal fibrosis in mouse experimental model. Kidney Int. 2004, 66, 1677–1685. [Google Scholar] [CrossRef] [Green Version]
- Sawada, T.; Ishii, Y.; Tojimbara, T.; Nakajima, I.; Fuchinoue, S.; Teraoka, S. The ACE inhibitor, quinapril, ameliorates peritoneal fibrosis in an encapsulating peritoneal sclerosis model in mice. Pharm. Res. 2002, 46, 505–510. [Google Scholar] [CrossRef]
- Tanabe, K.; Maeshima, Y.; Ichinose, K.; Kitayama, H.; Takazawa, Y.; Hirokoshi, K.; Kinomura, M.; Sugiyama, H.; Makino, H. Endostatin peptide, an inhibitor of angiogenesis, prevents the progression of peritoneal sclerosis in a mouse experimental model. Kidney Int. 2007, 71, 227–238. [Google Scholar] [CrossRef] [Green Version]
- Sekiguchi, Y.; Hamada, C.; Ro, Y.; Nakamoto, H.; Inaba, M.; Shimaoka, T.; Io, H.; Koyanagi, I.; Aruga, S.; Inuma, J.; et al. Differentiation of bone marrow-derived cells into regenerated mesothelial cells in peritoneal remodeling using a peritoneal fibrosis mouse model. J. Artif. Organs 2012, 15, 272–282. [Google Scholar] [CrossRef]
- Nakav, S.; Kachko, L.; Vorobiov, M.; Rogachev, B.; Chaimovitz, C.; Zlotnik, M.; Douvdevani, A. Blocking adenosine A2A receptor reduces peritoneal fibrosis in two independent experimental models. Nephrol. Dial. Transplant. 2009, 24, 2392–2399. [Google Scholar] [CrossRef]
- Yokoi, H.; Kasahara, M.; Mori, K.; Kuwabara, T.; Toda, N.; Yamada, R.; Namoto, S.; Yamamoto, T.; Seki, N.; Souma, N.; et al. Peritoneal Fibrosis and High Transport are Induced in Mildly Pre-Injured Peritoneum by 3,4-Dideoxyglucosone-3-Ene in Mice. Perit. Dial. Int. J. Int. Soc. Perit. Dial. 2013, 33, 143–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, S.; Takezawa, T.; Nagase, K.; Oshikata-Mitazaki, A.; Morito, S.; Sakumoto, T.; Masuda, M.; Yamamoto-Rikitake, M.; Akutagawa, T.; Toda, S. A high-density collagen xerogel thread prevents the progression of peritoneal fibrosis. Biomater. Sci. 2019, 7, 125–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrasekaran, P.; Izadjoo, S.; Stimely, J.; Palaniyandi, S.; Zhu, X.; Tafuri, W.; Mosser, D.M. Regulatory Macrophages Inhibit Alternative Macrophage Activation and Attenuate Pathology Associated with Fibrosis. J. Immunol. 2019, 203, 2130–2140. [Google Scholar] [CrossRef] [PubMed]
- Ertilav, M.; Hur, E.; Bozkurt, D.; Sipahi, S.; Timur, O.; Sarsik, B.; Akcicek, F.; Duman, S. Octreotide lessens peritoneal injury in experimental encapsulated peritoneal sclerosis model. Nephrology 2011, 16, 552–557. [Google Scholar] [CrossRef] [PubMed]
- Yokoi, H.; Kasahara, M.; Mori, K.; Ogawa, Y.; Kuwabara, T.; Imamaki, H.; Kawanishi, T.; Koga, K.; Ishii, A.; Kato, Y.; et al. Pleiotrophin triggers inflammation and increased peritoneal permeability leading to peritoneal fibrosis. Kidney Int. 2012, 81, 160–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakai, N.; Chun, J.; Duffield, J.S.; Wada, T.; Luster, A.D.; Tager, A.M. LPA 1—induced cytoskeleton reorganization drives fibrosis through CTGF-dependent fibroblast proliferation. FASEB J. 2013, 27, 1830–1846. [Google Scholar] [CrossRef] [Green Version]
- Kuret, T.; Sodin-Šemrl, S.; Mrak-Poljšak, K.; Čučnik, S.; Lakota, K.; Erman, A. Interleukin-1β Induces Intracellular Serum Amyloid A1 Expression in Human Coronary Artery Endothelial Cells and Promotes its Intercellular Exchange. Inflammation 2019, 42, 1413–1425. [Google Scholar] [CrossRef]
- STRING. Available online: https://string-db.org/ (accessed on 27 August 2021).
- Stergar, J.; Dolenec, R.; Kojc, N.; Lakota, K.; Perše, M.; Tomšič, M.; Milanic, M. Hyperspectral evaluation of peritoneal fibrosis in mouse models. Biomed. Opt. Express 2020, 11, 1991–2006. [Google Scholar] [CrossRef]
- Rogelj, L.; Pavlovcic, U.; Stergar, J.; Jezersek, M.; Simoncic, U.; Milanic, M. Curvature and height corrections of hyperspectral images using built-in 3D laser profilometry. Appl. Opt. 2019, 58, 9002–9012. [Google Scholar] [CrossRef]
- Dolenec, R.; Rogelj, L.; Stergar, J.; Milanic, M. Modular Multi-Wavelength LED Based Light Source for Hyperspectral Imaging. In Proceedings of the European Conference on Biomedical Optics 2019, Munich, Germany, 23–25 June 2019; Volume 11075. [Google Scholar]
- Stergar, J.; Hren, R.; Milanič, M. Design and Validation of a Custom-Made Laboratory Hyperspectral Imaging System for Biomedical Applications Using a Broadband LED Light Source. Sensors 2022, 22, 6274. [Google Scholar] [CrossRef]
- Welch, A.J.; Gemert, M.J.C. (Eds.) Optical-Thermal Response of Laser-Irradiated Tissue; Springer: Berlin, Germany, 2011. [Google Scholar] [CrossRef] [Green Version]
- Stergar, J.; Lakota, K.; Perše, M.; Tomšič, M.; Milanič, M. Hyperspectral evaluation of vasculature in induced peritonitis mouse models. Biomed. Opt. Express 2022, 13, 3461–3475. [Google Scholar] [CrossRef] [PubMed]
- Friebel, M.; Helfmann, J.; Netz, U.; Meinke, M. Influence of oxygen saturation on the optical scattering properties of human red blood cells in the spectral range 250 to 2000 nm. J. Biomed. Opt. 2009, 14, 034001. [Google Scholar] [CrossRef] [PubMed]
- Jacques, S.L. Optical properties of biological tissues: A review. Phys. Med. Biol. 2013, 58, R37–R61. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Wang, L.; Wang, M.; Zhou, S.; Lu, Y.; Cui, H.; Racanelli, A.C.; Zhang, L.; Ye, T.; Ding, B.; et al. Targeting fibrosis: Mechanisms and clinical trials. Signal Transduct. Target. Ther. 2022, 7, 206. [Google Scholar] [CrossRef]
- Sansbury, B.E.; Spite, M. Resolution of Acute Inflammation and the Role of Resolvins in Immunity, Thrombosis, and Vascular Biology. Circ. Res. 2016, 119, 113–130. [Google Scholar] [CrossRef]
- Lua, I.; Li, Y.; Pappoe, L.S.; Asahina, K. Myofibroblastic Conversion and Regeneration of Mesothelial Cells in Peritoneal and Liver Fibrosis. Am. J. Pathol. 2015, 185, 3258–3273. [Google Scholar] [CrossRef]
- Zou, J.; Li, Y.; Yu, J.; Dong, L.; Husain, A.N.; Shen, L.; Weber, C.R. Idiopathic pulmonary fibrosis is associated with tight junction protein alterations. Biochim. Biophys. Acta (BBA)-Biomembr. 2020, 1862, 183205. [Google Scholar] [CrossRef]
- Oakes, P.W.; Patel, D.C.; Morin, N.A.; Zitterbart, D.P.; Fabry, B.; Reichner, J.S.; Tang, J.X. Neutrophil morphology and migration are affected by substrate elasticity. Blood 2009, 114, 1387–1395. [Google Scholar] [CrossRef] [Green Version]
- Io, H.; Hamada, C.; Ro, Y.; Ito, Y.; Hirahara, I.; Tomino, Y. Morphologic changes of peritoneum and expression of VEGF in encapsulated peritoneal sclerosis rat models. Kidney Int. 2004, 65, 1927–1936. [Google Scholar] [CrossRef]
- Wang, H.-H.; Lee, T.-Y.; Lin, C.-Y. Integrins mediate adherence and migration of T lymphocytes on human peritoneal mesothelial cells. Kidney Int. 2008, 74, 808–816. [Google Scholar] [CrossRef] [Green Version]
- Hirose, M.; Nishino, T.; Obata, Y.; Nakazawa, M.; Nakazawa, Y.; Furusu, A.; Abe, K.; Miyazaki, M.; Koji, T.; Kohno, S. 22-Oxacalcitriol Prevents Progression of Peritoneal Fibrosis in a Mouse Model. Perit. Dial. Int. J. Int. Soc. Perit. Dial. 2013, 33, 132–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borceux, P.; Morelle, J.; Goffin, E. Complement system activation and peritoneal membrane alterations: Culprit or innocent bystander? Perit. Dial. Int. 2020, 40, 115–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poppelaars, F.; Faria, B.; Gaya Da Costa, M.; Franssen, C.F.M.; Van Son, W.J.; Berger, S.P.; Daha, M.R.; Seelen, M.A. The Complement System in Dialysis: A Forgotten Story? Front. Immunol. 2018, 9, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lubbers, R.; Van Essen, M.F.; Van Kooten, C.; Trouw, L.A. Production of complement components by cells of the immune system. Clin. Exp. Immunol. 2017, 188, 183–194. [Google Scholar] [CrossRef] [Green Version]
- Sei, Y.; Mizuno, M.; Suzuki, Y.; Imai, M.; Higashide, K.; Harris, C.L.; Sakata, F.; Iguchi, D.; Fujiwara, M.; Kodera, Y.; et al. Expression of membrane complement regulators, CD46, CD55 and CD59, in mesothelial cells of patients on peritoneal dialysis therapy. Mol. Immunol. 2015, 65, 302–309. [Google Scholar] [CrossRef]
- Kitterer, D.; Biegger, D.; Segerer, S.; Braun, N.; Alscher, M.D.; Latus, J. Alteration of membrane complement regulators is associated with transporter status in patients on peritoneal dialysis. PLoS ONE 2017, 12, e0177487. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, E.; Araújo, J.E.; Gómez-Meire, S.; Lodeiro, C.; Perez-Melon, C.; Iglesias-Lamas, E.; Otero-Glez, A.; Capelo, J.L.; Santos, H.M. Proteomics analysis of the peritoneal dialysate effluent reveals the presence of calcium-regulation proteins and acute inflammatory response. Clin. Proteom. 2014, 11, 17. [Google Scholar] [CrossRef] [Green Version]
- Reddingius, R.E.; Schroder, C.H.; Daha, M.R.; Willems, H.L.; Koster, A.M.; Monnens, L.A. Complement in serum and dialysate in children on continuous ambulatory peritoneal dialysis. Perit. Dial. Int. 1995, 15, 49–53. [Google Scholar] [CrossRef]
- Raby, A.-C.; Colmont, C.S.; Kift-Morgan, A.; Köhl, J.; Eberl, M.; Fraser, D.; Topley, N.; Labéta, M.O. Toll-Like Receptors 2 and 4 Are Potential Therapeutic Targets in Peritoneal Dialysis–Associated Fibrosis. J. Am. Soc. Nephrol. 2017, 28, 461–478. [Google Scholar] [CrossRef] [Green Version]
- Strang, C.J.; Cholin, S.; Spragg, J.; Davis, A.E.; Schneeberger, E.E.; Donaldson, V.H.; Rosen, F.S. Angioedema induced by a peptide derived from complement component C2. J. Exp. Med. 1988, 168, 1685–1698. [Google Scholar] [CrossRef]
- Théroux, P.; Martel, C. Complement activity and pharmacological inhibition in cardiovascular disease. Can. J. Cardiol. 2006, 22, 18B–24B. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1st Week | 2nd Week | 3rd Week | |
|---|---|---|---|
| Group 1 Ctrl 1 week (n = 4) | 200 μL PBS q.o.d. 1.6% EtOH in 100 μL PBS q.d. | / | |
| Group 2 CHX 1 week (n = 5) | 200 μL 0.1% CHX in 15% EtOH in PBS q.o.d. 1.6% EtOH in 100 μL PBS q.d. | ||
| Group 3 Ctrl 3 weeks (n = 4) | 200 μL PBS q.o.d. 1.6% EtOH in 100 μL PBS q.d. | ||
| Group 4 CHX 3 weeks (n = 5) | 200 μL 0.1% CHX in 15% EtOH in PBS q.o.d. 1.6% EtOH in 100 μL PBS q.d. | ||
| Group 5 Untreated Ctrl (n = 3) | No treatment | / | |
| Organ | Parameter | CHX | Control | ||
|---|---|---|---|---|---|
| 1 Week | 3 Weeks | 1 Week | 3 Weeks | ||
| liver | lymphocytes | ++ | +/++ | - | - |
| neutrophils | ++ | +/++ | - | - | |
| fibrosis | + | ++ | - | - | |
| spleen | lymphocytes | -/+ | -/+ | - | -/+ |
| neutrophils | + | ++ | -/+ | -/+ | |
| capsule | + | + | -/+ | + | |
| kidney | PMN/MNC | -/+ | -/+ | - | - |
| fibrosis | -/+ | -/+ | - | - | |
| Abd. wall peritoneum | PMNs | +/++ | +/++ | - | - |
| MNCs | + | -/+ | - | - | |
| edema | +/++ | +++ | - | - | |
| fibroblasts | + | ++ | - | - | |
| Abd. wall injury | depth | up to 25% | up to 25% | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brezovec, N.; Kojc, N.; Erman, A.; Hladnik, M.; Stergar, J.; Milanič, M.; Tomšič, M.; Čučnik, S.; Sodin-Šemrl, S.; Perše, M.; et al. Molecular and Cellular Markers in Chlorhexidine-Induced Peritoneal Fibrosis in Mice. Biomedicines 2022, 10, 2726. https://doi.org/10.3390/biomedicines10112726
Brezovec N, Kojc N, Erman A, Hladnik M, Stergar J, Milanič M, Tomšič M, Čučnik S, Sodin-Šemrl S, Perše M, et al. Molecular and Cellular Markers in Chlorhexidine-Induced Peritoneal Fibrosis in Mice. Biomedicines. 2022; 10(11):2726. https://doi.org/10.3390/biomedicines10112726
Chicago/Turabian StyleBrezovec, Neža, Nika Kojc, Andreja Erman, Matjaž Hladnik, Jošt Stergar, Matija Milanič, Matija Tomšič, Saša Čučnik, Snežna Sodin-Šemrl, Martina Perše, and et al. 2022. "Molecular and Cellular Markers in Chlorhexidine-Induced Peritoneal Fibrosis in Mice" Biomedicines 10, no. 11: 2726. https://doi.org/10.3390/biomedicines10112726