

Generation of Endogenous Promoter-Driven Luciferase Reporter System Using CRISPR/Cas9 for Investigating Transcriptional Regulation of the Core Clock Gene BMAL1

Abstract
:
1. Introduction
2. Methods and Material
2.1. Design of sgRNA Targeting BMAL1 Locus
2.2. Construction of Donor Plasmids
2.3. Cell Culture
2.4. Nucleo Transfection
2.5. Establishment of Stable Cell Line
2.6. Cell Viability and Cell Morphology
2.7. PCR-Based Detection
2.8. Immunoblot Analysis
2.9. Luciferase Assay
2.10. Statistical Analysis
3. Results
3.1. Construction of Donor Template
3.2. Generation of the U2-OS-Luc Cell Line
3.3. Validation of the U2-OS-Luc Cell Line
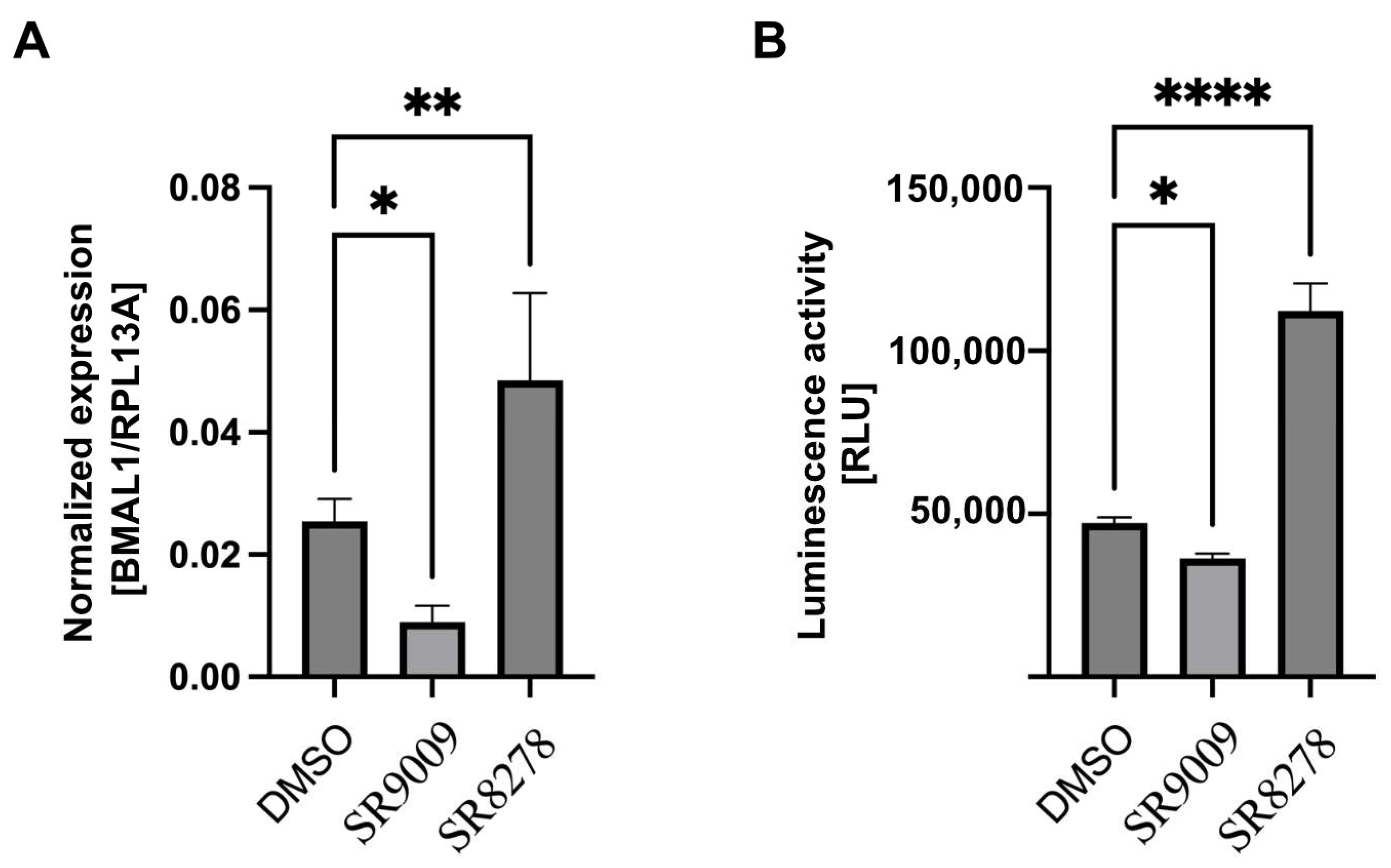
3.4. Luciferase Reporter Gene Responds to Circadian Rhythm in U2-OS Cell Line
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Landrigan, P.J.; Fuller, R.; Acosta, N.J.R.; Adeyi, O.; Arnold, R.; Basu, N.N.; Baldé, A.B.; Bertollini, R.; Bose-O’Reilly, S.; Boufford, J.I.; et al. The Lancet Commission on pollution and health. Lancet 2018, 391, 462–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasi, S.; Shams, T.; Masood, A. Toxicological effects of major environmental pollutants: An overview. Environ. Monit. Assess. 2013, 185, 2585–2593. [Google Scholar] [CrossRef] [PubMed]
- Ng, A.; Weerakoon, D.; Lim, E.; Padhye, L.P. Fate of environmental pollutants. Water Environ. Res. 2019, 91, 1294–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Magueresse-Battistoni, B.; Hubert, V.; Danielle, N. Environmental pollutants and metabolic disorders: The multi-exposure scenario of life. Front. Endocrinol. 2018, 9, 582. [Google Scholar] [CrossRef] [Green Version]
- Joffe, M. Infertility and environmental pollutants. Br. Med. Bull. 2003, 68, 47–70. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Hidaka, T.; Kumagai, Y.; Yamamoto, M. Environmental pollutants and the immune response. Nat. Immunol. 2020, 21, 1486–1495. [Google Scholar] [CrossRef]
- Boffetta, P. Human cancer from environmental pollutants: The epidemiological evidence. Mutat. Res. Genet. Toxicol. Environ. Mutagenesis 2006, 608, 157–162. [Google Scholar] [CrossRef]
- Naville, D.; Gaillard, G.; Julien, B.; Vega, N.; Pinteur, C.; Chanon, S.; Vidal, H.; Le Magueresse-Battistoni, B. Chronic exposure to a pollutant mixture at low doses led to tissue-specific metabolic alterations in male mice fed standard and high-fat high-sucrose diet. Chemosphere 2019, 220, 1187–1199. [Google Scholar] [CrossRef]
- Fossi, M.C. Biomarkers as diagnostic and prognostic tools for wildlife risk assessment: Integrating endocrine-disrupting chemicals. Toxicol. Ind. Health 1998, 14, 291–309. [Google Scholar] [CrossRef]
- Giesy, J.P.; Feyk, L.A.; Jones, P.D.; Kannan, K.; Sanderson, T. Review of the effects of endocrine-disrupting chemicals in birds. Pure Appl. Chem. 2003, 75, 2287–2303. [Google Scholar] [CrossRef]
- Stoker, T.E.; Parks, L.G.; Gray, L.E.; Cooper, R.L. Endocrine-disrupting chemicals: Prepubertal exposures and effects on sexual maturation and thyroid function in the male rat. A focus on the EDSTAC recommendations. Crit. Rev. Toxicol. 2000, 30, 197–252. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Zhang, K.; Zhao, Y.; Fent, K. Environmental chemicals affect circadian rhythms: An underexplored effect influencing health and fitness in animals and humans. Environ. Int. 2021, 149, 106159. [Google Scholar] [CrossRef] [PubMed]
- Jatkowska, N.; Kudłak, B.; Lewandowska, P.; Liu, W.; Williams, M.J.; Schiöth, H.B. Identification of synergistic and antagonistic actions of environmental pollutants: Bisphenols A, S and F in the presence of DEP, DBP, BADGE and BADGE 2HCl in three component mixtures. Sci. Total Environ. 2021, 767, 144286. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Gupta, V.K.; Kumar, A.; Sharma, B. Synergistic effects of heavy metals and pesticides in living systems. Front. Chem. 2017, 5, 70. [Google Scholar] [CrossRef] [PubMed]
- Carnevali, O.; Santangeli, S.; Forner-Piquer, I.; Basili, D.; Maradonna, F. Endocrine-Disrupting Chemicals in Aquatic Environment: What Are the Risks for Fish Gametes? Fish Physiol. Biochem. 2018, 44, 1561–1576. [Google Scholar] [CrossRef] [PubMed]
- Kudłak, B.; Jatkowska, N.; Liu, W.; Williams, M.J.; Barcelo, D.; Schiöth, H.B. Enhanced Toxicity of Bisphenols Together with UV Filters in Water: Identification of Synergy and Antagonism in Three-Component Mixtures. Molecules 2022, 27, 3260. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-H.; David, R.J. Methodological issues in human studies of endocrine disrupting chemicals. Rev. Endocr. Metab. Disord. 2015, 16, 289–297. [Google Scholar] [CrossRef]
- Zhang, S.; Kang, Q.; Peng, H.; Ding, M.; Zhao, F.; Zhou, Y.; Dong, Z.; Zhang, H.; Yang, M.; Tao, S.; et al. Relationship between perfluorooctanoate and perfluorooctane sulfonate blood concentrations in the general population and routine drinking water exposure. Environ. Int. 2019, 126, 54–60. [Google Scholar] [CrossRef]
- Liu, W.; Cao, H.; Liao, S.; Kudłak, B.; Williams, M.J.; Schiöth, H.B. Dibutyl phthalate disrupts conserved circadian rhythm in Drosophila and human cells. Sci. Total Environ. 2021, 783, 147038. [Google Scholar] [CrossRef]
- Brunner, M.; Tobias, S. Transcriptional and post-transcriptional regulation of the circadian clock of cyanobacteria and Neurospora. Genes Dev. 2006, 20, 1061–1074. [Google Scholar] [CrossRef]
- Gachon, F.; Nagoshi, E.; Brown, S.A.; Ripperger, J.; Schibler, U. The mammalian circadian timing system: From gene expression to physiology. Chromosoma 2004, 113, 103–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partch, C.L.; Carla, B.G.; Joseph, S.T. Molecular architecture of the mammalian circadian clock. Trends Cell Biol. 2014, 24, 90–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patke, A.; Michael, W.Y.; Sofia, A. Molecular mechanisms and physiological importance of circadian rhythms. Nat. Rev. Mol. Cell Biol. 2020, 21, 67–84. [Google Scholar] [CrossRef]
- Refinetti, R.; Michael, M. The circadian rhythm of body temperature. Physiol. Behav. 1992, 51, 613–637. [Google Scholar] [CrossRef]
- Guido, M.E.; Monjes, N.M.; Wagner, P.M.; Salvador, G.A. Circadian Regulation and Clock-Controlled Mechanisms of Glycerophospholipid Metabolism from Neuronal Cells and Tissues to Fibroblasts. Mol. Neurobiol. 2022, 59, 326–353. [Google Scholar] [CrossRef] [PubMed]
- Challet, E. The circadian regulation of food intake. Nat. Rev. Endocrinol. 2019, 15, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, H.; Fernández, A.F.; Setién, F.; Ropero, S.; Ballestar, E.; Villanueva, A.; Yamamoto, H.; Imai, K.; Shinomura, Y.; Esteller, M. Epigenetic Inactivation of the Circadian Clock Gene BMAL1 in Hematologic Malignancies BMAL1 Epigenetic Inactivation. Cancer Res. 2009, 69, 8447–8454. [Google Scholar] [CrossRef] [Green Version]
- Miller, B.H.; McDearmon, E.L.; Panda, S.; Hayes, K.R.; Zhang, J.; Andrews, J.L.; Antoch, M.P.; Walker, J.R.; Esser, K.A.; Hogenesch, J.B.; et al. Circadian and CLOCK-controlled regulation of the mouse transcriptome and cell proliferation. Proc. Natl. Acad. Sci. USA 2007, 104, 3342–3347. [Google Scholar] [CrossRef] [Green Version]
- Storch, K.-F.; Lipan, O.; Leykin, I.; Viswanathan, N.; Davis, F.C.; Wong, W.H.; Weitz, C.J. Extensive and divergent circadian gene expression in liver and heart. Nature 2002, 417, 78–83. [Google Scholar] [CrossRef]
- Gauger, M.A.; Aziz, S. Cryptochrome, circadian cycle, cell cycle checkpoints, and cancer. Cancer Res. 2005, 65, 6828–6834. [Google Scholar] [CrossRef]
- Savvidis, C.; Michael, K. Circadian rhythm disruption in cancer biology. Mol. Med. 2012, 18, 1249–1260. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, F.C.; Aparna, R.; Anne, M. Circadian molecular clocks and cancer. Cancer Lett. 2014, 342, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Gaddameedhi, S.; Selby, C.P.; Kaufmann, W.K.; Smart, R.C.; Sancar, A. Control of skin cancer by the circadian rhythm. Proc. Natl. Acad. Sci. USA 2011, 108, 18790–18795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Zhao, S.; Jiang, X.; Zhang, E.; Hu, G.; Hu, B.; Zheng, P.; Xiao, J.; Lu, Z.; Lu, Y.; et al. The circadian clock gene Bmal1 acts as a potential anti-oncogene in pancreatic cancer by activating the p53 tumor suppressor pathway. Cancer Lett. 2016, 371, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Lagunas-Rangel, F.A.; Kudłak, B.; Liu, W.; Williams, M.J.; Schiöth, H.B. The potential interaction of environmental pollutants and circadian rhythm regulations that may cause leukemia. Crit. Rev. Environ. Sci. Technol. 2021, 52, 4094–4112. [Google Scholar] [CrossRef]
- Lagunas-Rangel, F.A.; Wen, L.; Helgi, B. Can Exposure to Environmental Pollutants Be Associated with Less Effective Chemotherapy in Cancer Patients? Int. J. Environ. Res. Public Health 2022, 19, 2064. [Google Scholar] [CrossRef] [PubMed]
- Lagunas-Rangel, F.A.; Linnea-Niemi, J.V.; Kudłak, B.; Williams, M.J.; Jönsson, J.; Schiöth, H.B. Role of the synergistic interactions of environmental pollutants in the development of cancer. GeoHealth. 2022, 6, e2021GH000552. [Google Scholar] [CrossRef]
- Lukashina, N.; Williams, M.J.; Kartysheva, E.; Virko, E.; Kudłak, B.; Fredriksson, R.; Spjuth, O.; Schiöth, H.B. Integrating statistical and machine-learning approach for meta-analysis of bisphenol A-exposure datasets reveals effects on mouse gene expression within pathways of apoptosis and cell survival. Int. J. Mol. Sci. 2021, 22, 10785. [Google Scholar] [CrossRef]
- Vollmers, C.; Satchidananda, P.; Luciano, D. A high-throughput assay for siRNA-based circadian screens in human U2OS cells. PLoS ONE 2008, 3, e3457. [Google Scholar] [CrossRef] [Green Version]
- Hirota, T.; Lee, J.W.; Lewis, W.G.; Zhang, E.E.; Breton, G.; Liu, X.; Garcia, M.; Peters, E.C.; Etchegaray, J.-P.; Traver, D.; et al. High-throughput chemical screen identifies a novel potent modulator of cellular circadian rhythms and reveals CKIα as a clock regulatory kinase. PLoS Biol. 2010, 8, e1000559. [Google Scholar] [CrossRef]
- Hoffmann, J.; Symul, L.; Shostak, A.; Fischer, T.; Naef, F.; Brunner, M. Non-circadian expression masking clock-driven weak transcription rhythms in U2OS cells. PLoS ONE 2014, 9, e102238. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.J.; Cao, H.; Lindkvist, T.; Mothes, T.J.; Schiöth, H.B. Exposure to the Environmental Pollutant Bisphenol A Diglycidyl Ether (BADGE) Causes Cell over-Proliferation in Drosophila. Environ. Sci. Pollut. Res. Int. 2020, 27, 25261–25270. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.J.; Wang, Y.; Klockars, A.; Monica Lind, P.; Fredriksson, R.; Schiöth, H.B. Exposure to bisphenol A affects lipid metabolism in Drosophila melanogaster. Basic Clin. Pharmacol. Toxicol. 2014, 114, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Cao, H. Exposure to xenobiotic chemicals disrupts metabolism, rhythmicity and cell proliferation in Drosophila melanogaster. Ph.D. Thesis, Acta Universitatis Upsaliensis. Uppsala University, Uppsala, Sweden, 2018. [Google Scholar]
- Ramanathan, C.; Khan, S.K.; Kathale, N.D.; Xu, H.; Liu, A.C. Monitoring cell-autonomous circadian clock rhythms of gene expression using luciferase bioluminescence reporters. JoVE J. Vis. Exp. 2012, 67, e4234. [Google Scholar] [CrossRef] [Green Version]
- Isojima, Y.; Nakajima, M.; Ukai, H.; Fujishima, H.; Yamada, R.G.; Masumoto, K.; Kiuchi, R.; Ishida, M.; Ukai-Tadenuma, M.; Minami, Y.; et al. CKI/-dependent phosphorylation is a temperature-insensitive, period-determining process in the mammalian circadian clock. Proc. Natl. Acad. Sci. USA 2009, 106, 15744–15749. [Google Scholar] [CrossRef] [Green Version]
- Maier, B.; Wendt, S.; Vanselow, J.T.; Wallach, T.; Reischl, S.; Oehmke, S.; Schlosser, A.; Kramer, A. A large-scale functional RNAi screen reveals a role for CK2 in the mammalian circadian clock. Genes Dev. 2009, 23, 708–718. [Google Scholar] [CrossRef] [Green Version]
- Wood, A.J.; Lo, T.-W.; Zeitler, B.; Pickle, C.S.; Ralston, E.J.; Lee, A.H.; Amora, R.; Miller, J.C.; Leung, E.; Meng, X.; et al. Targeted genome editing across species using ZFNs and TALENs. Science 2011, 333, 307. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.M.; Kiran, M. Expanding the genetic editing tool kit: ZFNs, TALENs, and CRISPR-Cas9. J. Clin. Investig. 2014, 124, 4154–4161. [Google Scholar] [CrossRef] [Green Version]
- Makarova, K.S.; Haft, D.H.; Barrangou, R.; Brouns, S.J.J.; Charpentier, E.; Horvath, P.; Moineau, S.; Mojica, F.J.M.; Wolf, Y.I.; Yakunin, A.F.; et al. Evolution and classification of the CRISPR–Cas systems. Nat. Rev. Microbiol. 2011, 9, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Hsu, P.D.; Eric, S.L.; Feng, Z. Development and applications of CRISPR-Cas9 for genome engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef]
- Doudna, J.A.; Emmanuelle, C. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef] [PubMed]
- Nishimasu, H.; Shi, X.; Ishiguro, S.; Gao, L.; Hirano, S.; Okazaki, S.; Noda, T.; Abudayyeh, O.O.; Gootenberg, J.S.; Mori, H.; et al. Engineered CRISPR-Cas9 nuclease with expanded targeting space. Science 2018, 361, 1259–1262. [Google Scholar] [CrossRef] [PubMed]
- Gong, K.W.; Zhao, W.; Li, N.; Barajas, B.; Kleinman, M.; Sioutas, C.; Horvath, S.; Lusis, A.J.; Nel, A.; Araujo, J.A. Air-pollutant chemicals and oxidized lipids exhibit genome-wide synergistic effects on endothelial cells. Genome Biol. 2007, 8, R149.1-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palanivel, R.; Vinayachandran, V.; Biswal, S.; Deiuliis, J.A.; Padmanabhan, R.; Park, B.; Gangwar, R.S.; Durieux, J.C.; Ebreo Cara, E.A.; Das, L.; et al. Exposure to air pollution disrupts circadian rhythm through alterations in chromatin dynamics. Iscience 2020, 23, 101728. [Google Scholar] [CrossRef]
- Benedusi, M.; Frigato, E.; Bertolucci, C.; Valacchi, G. Circadian deregulation as possible new player in pollution-induced tissue damage. Atmosphere 2021, 12, 116. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer Sequence (5′-3′) | Use |
|---|---|---|
| PF-luc-puro | AGATGTCGAAGAGAATCCTGGACCGATGGAAGACGCCAAAAACATAAAG | PCR for luciferase expression cassette |
| PR-luc-puro | CTGTTGCCAAAGCAACATGTAGTGTTCCACATAGCGTAAAAGGAGCAACATAG | PCR for luciferase expression cassette |
| PF-L | GCTGGCTAGCGTTTAAACTTAAGCTTTGGAGGTCCAAGTTTGTGCCTGGAA | Infusion for homology domain |
| PR-L | TCCAGGATTCTCTTCGACATCT | Infusion for homology domain |
| PF-R | CTATGTTGCTCCTTTTACGCTATGT | Infusion for homology domain |
| PR-R | GGACTAGTGGATCCGAGCTCGGTACCAGAGAGGTGCAGCATTAGAGAAGCC | Infusion for homology domain |
| PF-luc | CCGGCGCCATTCTATCCTC | PCR validation of luciferase knock-in |
| PR-luc | CCTTTCGGTACTTCGTCCACA | PCR validation of luciferase knock-in |
| PF-BMAL1 | TTAAGAGGTGCCACCAATCC | qPCR for BMAL1 |
| PR-BMAL1 | CTTCCCTCGGTCACATCCTA | qPCR for BMAL1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, C.; Li, C.; Liu, W.; Schiöth, H.B. Generation of Endogenous Promoter-Driven Luciferase Reporter System Using CRISPR/Cas9 for Investigating Transcriptional Regulation of the Core Clock Gene BMAL1. Biomedicines 2022, 10, 3108. https://doi.org/10.3390/biomedicines10123108
Sun C, Li C, Liu W, Schiöth HB. Generation of Endogenous Promoter-Driven Luciferase Reporter System Using CRISPR/Cas9 for Investigating Transcriptional Regulation of the Core Clock Gene BMAL1. Biomedicines. 2022; 10(12):3108. https://doi.org/10.3390/biomedicines10123108
Chicago/Turabian StyleSun, Chengxi, Chen Li, Wen Liu, and Helgi B. Schiöth. 2022. "Generation of Endogenous Promoter-Driven Luciferase Reporter System Using CRISPR/Cas9 for Investigating Transcriptional Regulation of the Core Clock Gene BMAL1" Biomedicines 10, no. 12: 3108. https://doi.org/10.3390/biomedicines10123108