
Chlorogenic Acid Isomers Isolated from Artemisia lavandulaefolia Exhibit Anti-Rosacea Effects In Vitro

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Chemical Standards and Solvents
2.3. Preparation of A. lavandulaefolia extract (ALE)
2.4. Preparation of Standard and ALE Solution
2.5. HPLC Analysis
2.6. HPLC MS/MS Analysis
2.7. KLK5 Protease Activity
2.8. KLK5 Expression
2.9. Cathelicidin Cleavage Assay
2.10. Co-Culture Conditions
2.10.1. HEKn/THP-1 Co-Culture
2.10.2. HEKn/MC/9 Co-Culture
2.11. Total RNA Extraction, cDNA Synthesis, and Quantitative PCR
2.12. HMEC-1 Proliferation Assay
2.13. HMEC-1 Migration Assay
2.14. Statistical Analysis
3. Results
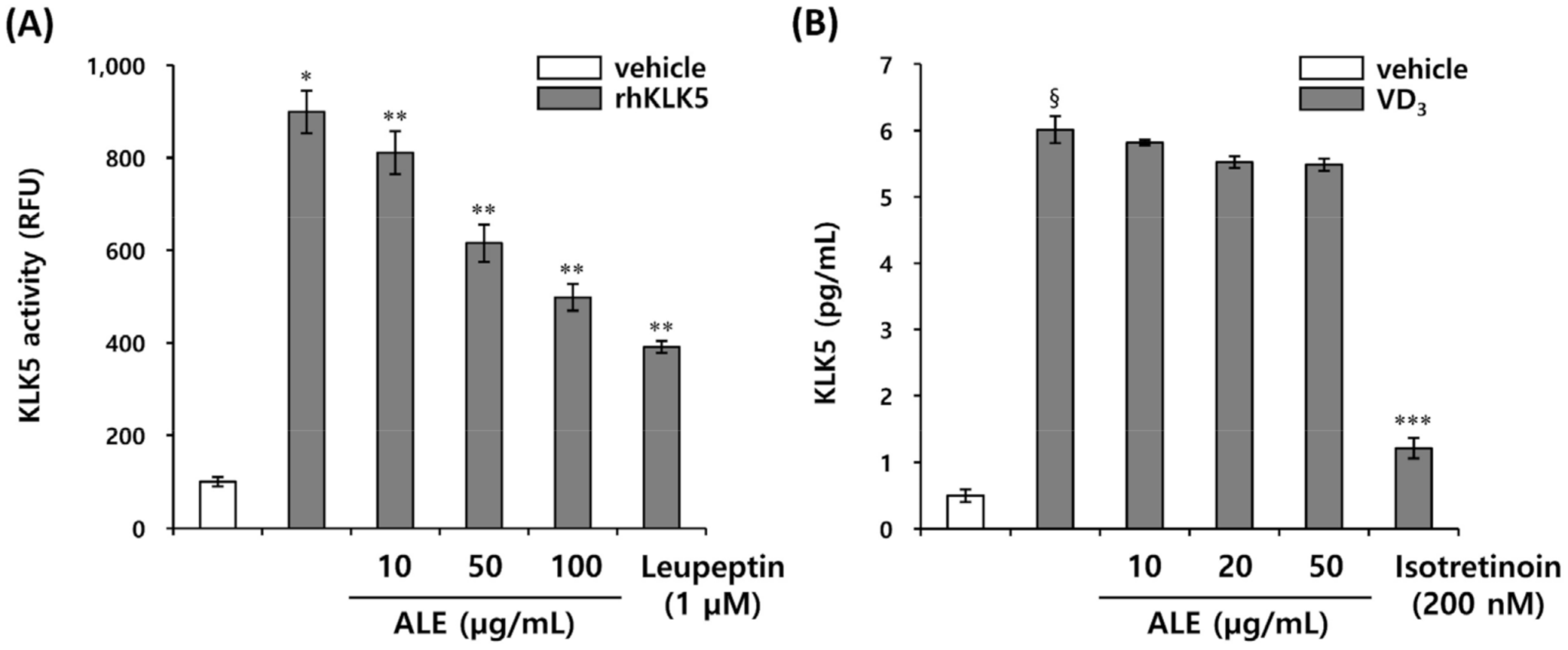
3.1. ALE Inhibits KLK5 Activity
3.2. Isolation and Identification of Active Compounds from A. lavandulaefolia as KLK5 Inhibitors
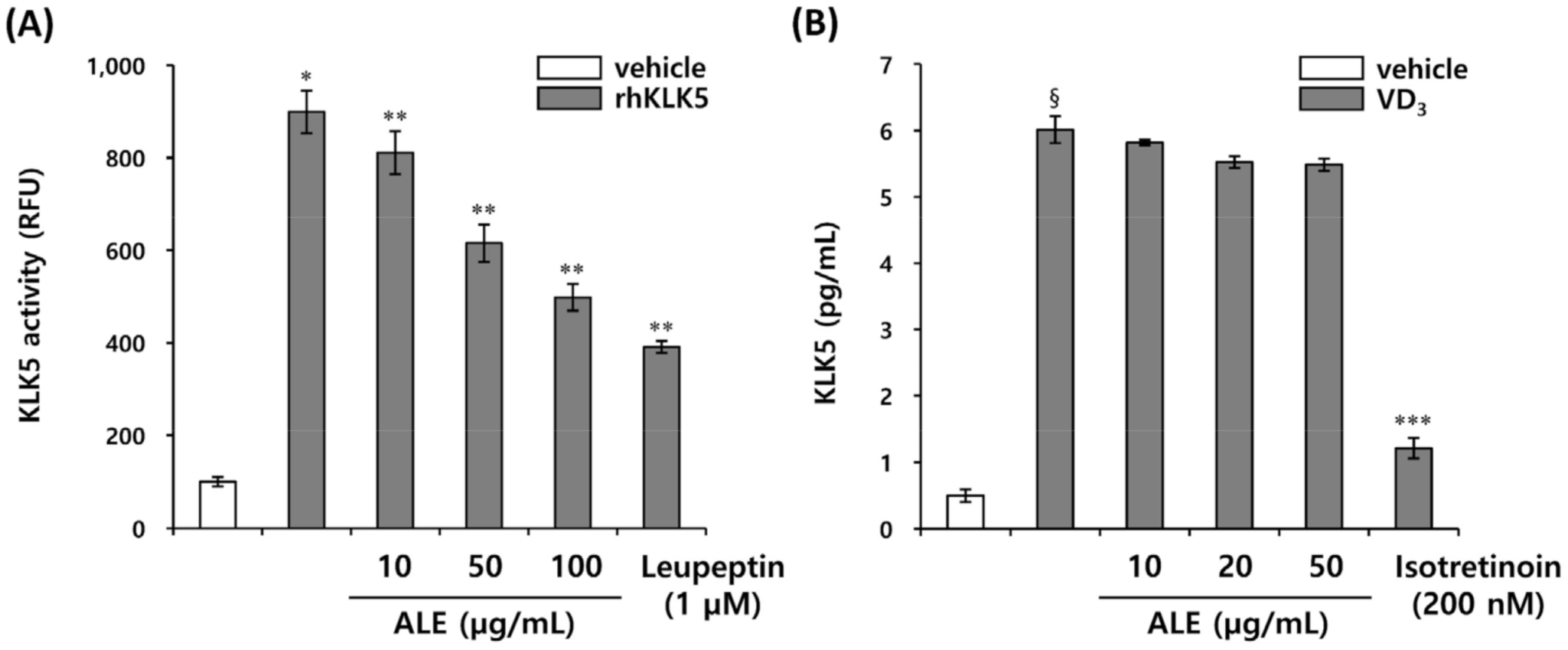
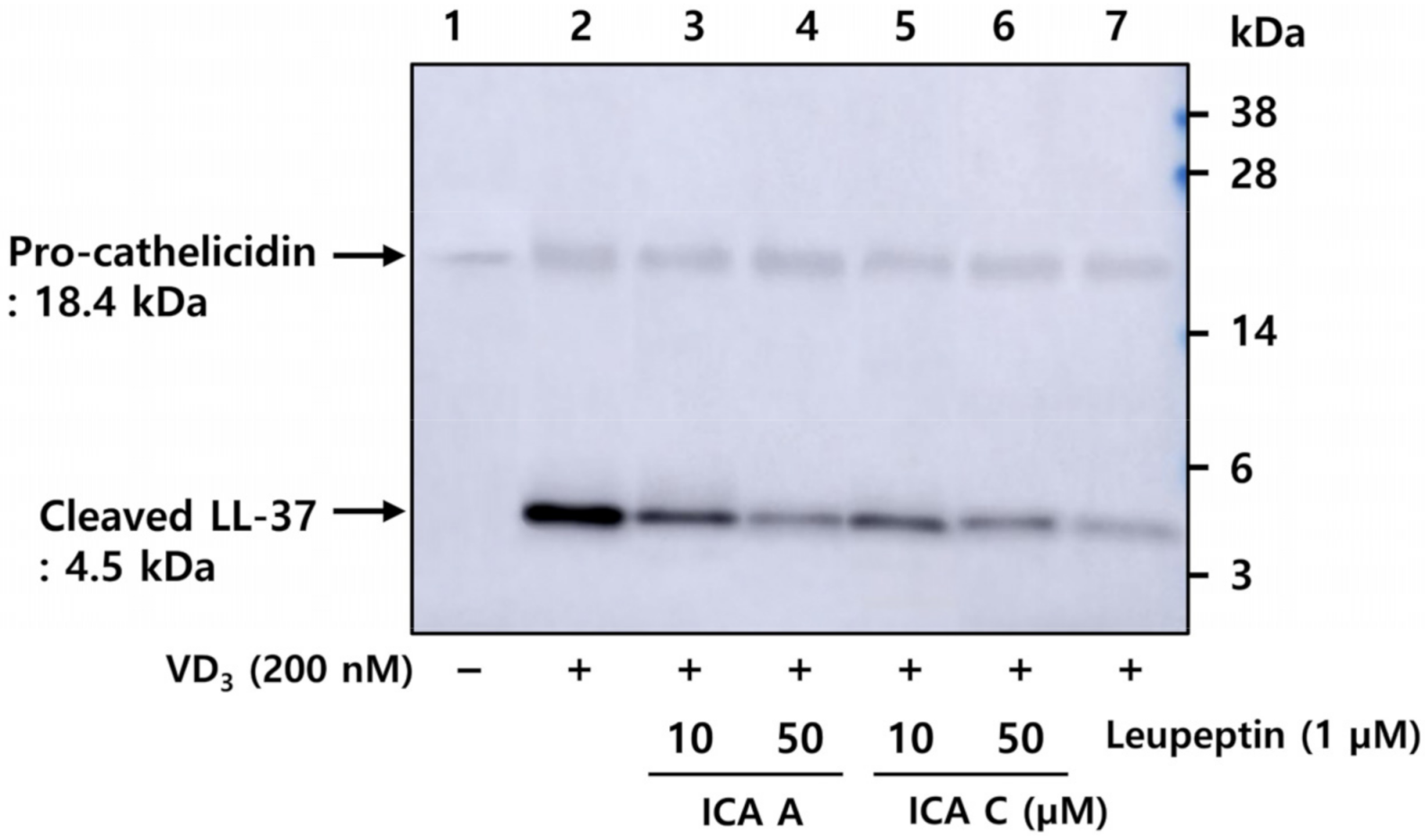
3.3. Chlorogenic Acid Isomers Inhibit the Proteolytic Cleavage of Cathelicidin by Blocking KLK5 Protease Activity
3.4. Chlorogenic Acid Isomers Decrease the Expression of Proinflammatory Cytokines Induced by VD3
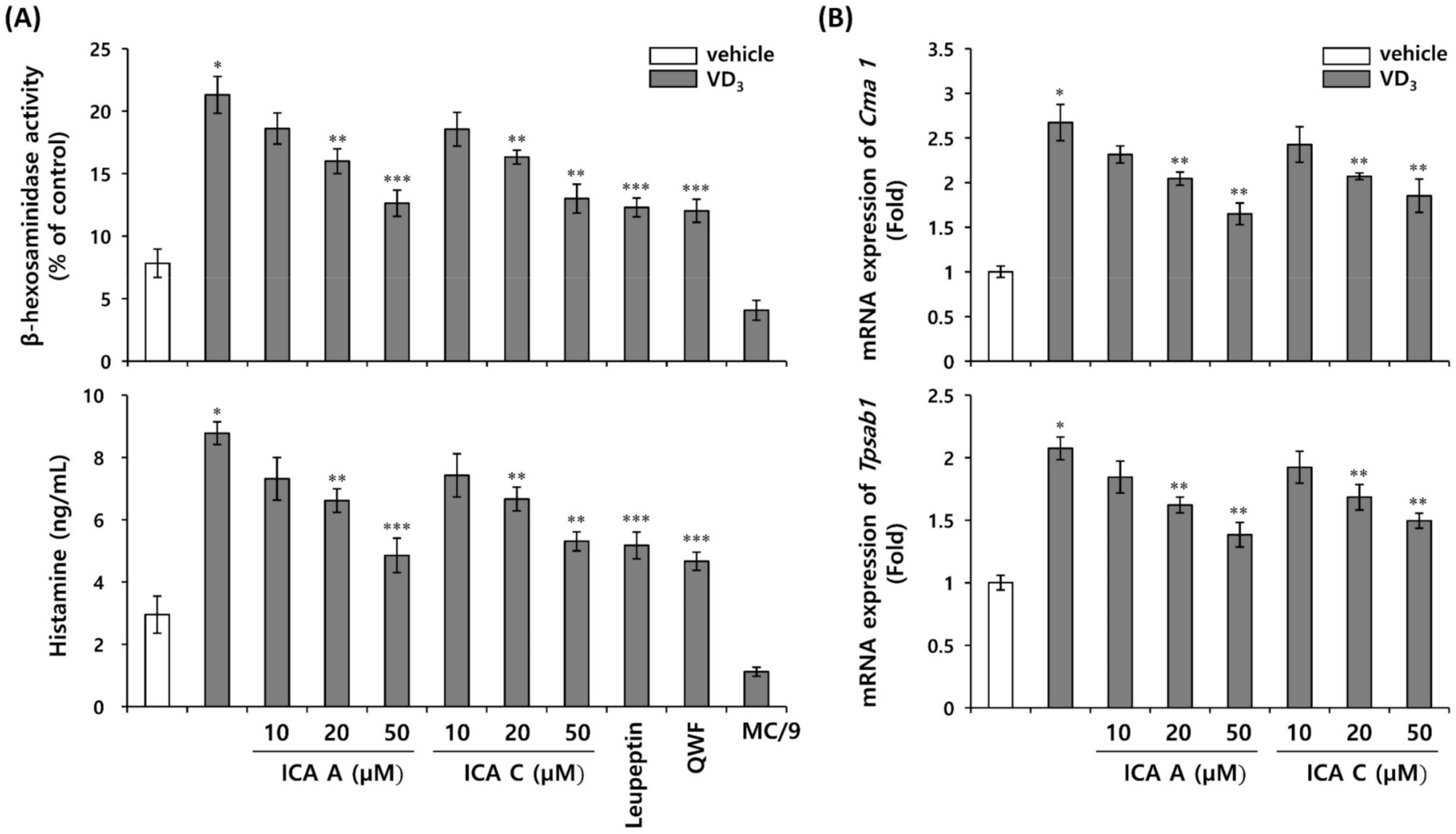
3.5. Inhibition of VD3-Induced Mast Cell Activation by Chlorogenic Acid Isomers
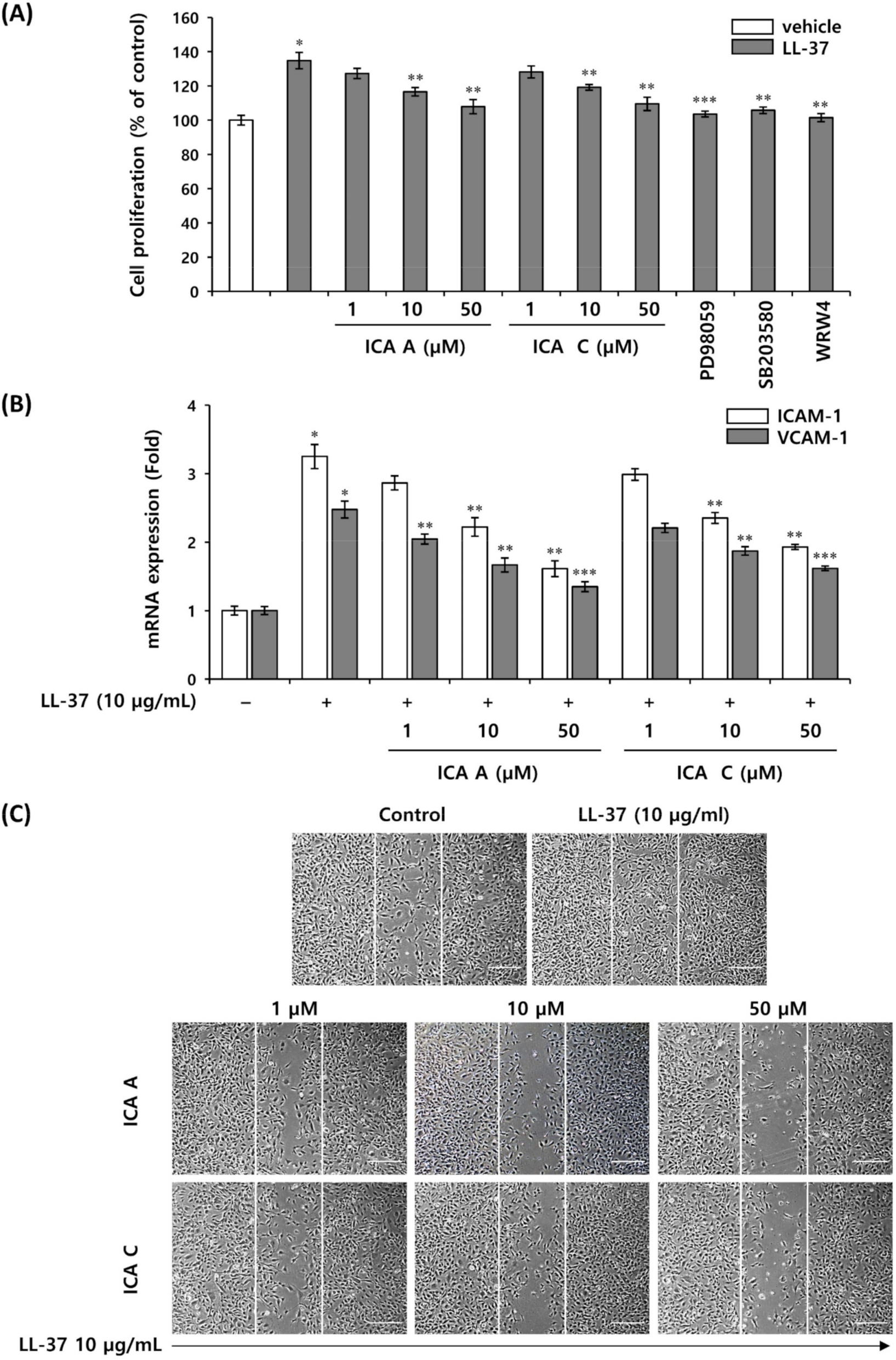
3.6. Isochlorogenic Acid Isomers Regulate LL-37-Induced Vascular Endothelial Cell Activation
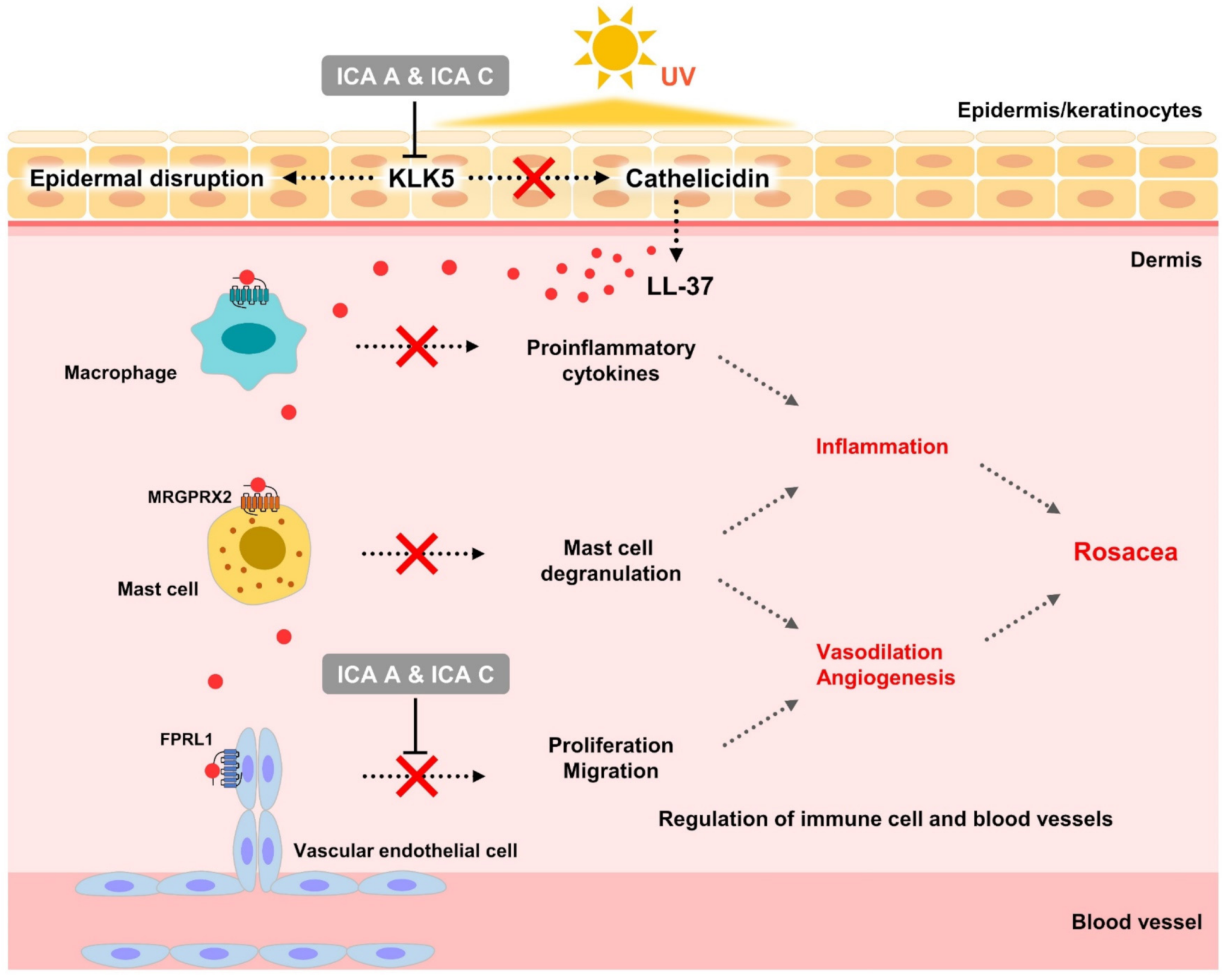
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kościuczuk, E.M.; Lisowski, P.; Jarczak, J.; Strzałkowska, N.; Jóźwik, A.; Horbańczuk, J.; Krzyżewski, J.; Zwierzchowski, L.; Bagnicka, E. Cathelicidins: Family of antimicrobial peptides. Mol. Biol. Rep. 2012, 39, 10957–10970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, M.G.; Davidson, D.J.; Gold, M.R.; Bowdish, D.; Hancock, R.E. The human antimicrobial peptide LL-37 is a multifunctional modulator of innate immune responses. J. Immunol. 2002, 169, 3883–3891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Chen, Q.; Schmidt, A.P.; Anderson, G.M.; Wang, J.M.; Wooters, J.; Oppenheim, J.J.; Chertov, O. LL-37, the neutrophil granule- and epithelial cell-derived cathelicidin, utilizes formyl peptide receptor-like 1 (FPRL1) as a receptor to chemoattract human peripheral blood neutrophils, monocytes, and T cells. J. Exp. Med. 2000, 192, 1069–1074. [Google Scholar] [CrossRef] [PubMed]
- Boekstegers, P.; Welsch, U.; Hiemstra, P.S.; Vogelmeier, C.; Gallo, R.L.; Clauss, M.; Bals, R. An angiogenic role for the human peptide antibiotic LL-37/hCAP-18. J. Clin. Investig. 2003, 111, 1665–1672. [Google Scholar]
- Gallo, R.L.; Ono, M.; Povsic, T.; Page, C.; Eriksson, E.; Klagsbrun, M.; Bernfield, M. Syndecans, cell surface heparan sulfate proteoglycans, are induced by a proline-rich antimicrobial peptide from wounds. Proc. Natl. Acad. Sci. USA 1994, 91, 11035–11039. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, K.; Di Nardo, A.; Bardan, A.; Murakami, M.; Ohtake, T.; Coda, A.; Dorschner, R.A.; Bonnart, C.; Descargues, P.; Hovnanian, A.; et al. Increased serine protease activity and cathelicidin promotes skin inflammation in rosacea. Nat. Med. 2007, 13, 975–980. [Google Scholar] [CrossRef]
- Woo, Y.R.; Lim, J.H.; Cho, D.H.; Park, H.J. Rosacea: Molecular mechanisms and management of a chronic cutaneous inflammatory condition. Int. J. Mol. Sci. 2016, 17, 1562. [Google Scholar] [CrossRef] [Green Version]
- Byrne, A.M.; Bouchier-Hayes, D.J.; Harmey, J.H. Angiogenic and cell survival functions of vascular endothelial growth factor (VEGF). J. Cell. Mol. Med. 2005, 9, 777–794. [Google Scholar] [CrossRef]
- Fràter-Schröder, M.; Risau, W.; Hallmann, R.; Gautschi, P.; Böhlen, P. Tumor necrosis factor type alpha, a potent inhibitor of endothelial cell growth in vitro, is angiogenic in vivo. Proc. Natl. Acad. Sci. USA 1987, 84, 5277–5281. [Google Scholar] [CrossRef] [Green Version]
- Salcedo, R.; Ponce, M.L.; Young, H.A.; Wasserman, K.; Ward, J.M.; Kleinman, H.K.; Oppenheim, J.J.; Murphy, W.J. Human endothelial cells express CCR2 and respond to MCP-1: Direct role of MCP-1 in angiogenesis and tumor progression. Blood 2000, 96, 34–40. [Google Scholar] [CrossRef]
- Lee, S.Y.; Lee, M.S.; Lee, H.Y.; Kim, S.D.; Shim, J.W.; Jo, S.H.; Lee, J.W.; Kim, J.Y.; Choi, Y.W.; Baek, S.H.; et al. F2L, a peptide derived from heme-binding protein, inhibits LL-37-induced cell proliferation and tube formation in human umbilical vein endothelial cells. FEBS Lett. 2008, 582, 273–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krystel-Whittemore, M.; Dileepan, K.N.; Wood, J.G. Mast cell: A multi-functional master cell. Front. Immunol. 2016, 6, 620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caughey, G.H. Mast cell tryptases and chymases in inflammation and host defense. Immunol. Rev. 2007, 217, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.A.; Park, M.; Kim, Y.H.; Woo, S.Y. Th17 cell-mediated immune responses promote mast cell proliferation by triggering stem cell factor in keratinocytes. Biochem. Biophys. Res. Commun. 2017, 487, 856–861. [Google Scholar] [CrossRef] [PubMed]
- Stassen, M.; Hartmann, A.K.; Delgado, S.J.; Dehmel, S.; Braun, A. Mast cells within cellular networks. J. Allergy Clin. Immunol. 2019, 144, S46–S54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchitto, M.C.; Chien, A.L. Mast cell stabilizers in the treatment of rosacea: A review of existing and emerging therapies. Dermatol. Ther. 2021, 11, 1541–1549. [Google Scholar] [CrossRef] [PubMed]
- Muto, Y.; Wang, Z.; Vanderberghe, M.; Two, A.; Gallo, R.L.; Di Nardo, A. Mast cells are key mediators of cathelicidin-initiated skin inflammation in rosacea. J. Investig. Dermatol. 2014, 134, 2728–2736. [Google Scholar] [CrossRef] [Green Version]
- Di Nardo, A.; Vitiello, A.; Gallo, R.L. Cutting edge: Mast cell antimicrobial activity is mediated by expression of cathelicidin antimicrobial peptide. J. Immunol. 2003, 170, 2274–2278. [Google Scholar] [CrossRef]
- Hussein, R.A.; El-Anssary, A.A. Plants secondary metabolites: The key drivers of the pharmacological actions of medicinal plants. In Herbal Medicine; Builders, P.F., Ed.; IntechOpen: London, UK, 2018. [Google Scholar]
- Li, F.S.; Weng, J.K. Demystifying traditional herbal medicine with modern approach. Nat. Plants 2017, 3, 17109. [Google Scholar] [CrossRef]
- Cha, J.D.; Jeong, M.R.; Choi, H.J.; Jeong, S.I.; Moon, S.E.; Yun, S.I.; Kim, Y.H.; Kil, B.S.; Song, Y.H. Chemical composition and antimicrobial activity of the essential oil of Artemisia lavandulaefolia. Planta Med. 2005, 71, 575–577. [Google Scholar] [CrossRef]
- Abad, M.J.; Bedoya, L.M.; Apaza, L.; Bermejo, P. The Artemisia L. genus: A review of bioactive essential oils. Molecules 2012, 17, 2542–2566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Liu, T.; Zhou, C.; Huang, Y.; Liu, X.; Yuan, H. Antifungal activity of essential oils from three Artemisia species against Colletotrichum gloeosporioides of mango. Antibiotics 2021, 10, 1331. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.B.; Guo, L.M.; Wang, F.L.; Lü, J.L. Phytochemical profile and anti-inflammatory activity of the fraction from Artemisia lavandulaefolia. Chem. Biodivers. 2021, 18, e2000989. [Google Scholar] [CrossRef] [PubMed]
- Yi, E.Y.; Han, K.S.; Kim, Y.J. Extract of Artemisia lavandulaefolia inhibits in vitro angiogenesis in human umbilical vein endothelial cells. J. Cancer Prev. 2014, 19, 247–252. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Zou, K.; Zhang, W.; Guo, S.; Liu, H.; Sun, J.; Li, J.; Huang, D.; Wu, Y.; Du, S.; et al. Efficacy of compounds isolated from the essential oil of Artemisia lavandulaefolia in control of the cigarette beetle, Lasioderma serricorne. Molecules 2018, 23, 343. [Google Scholar] [CrossRef] [Green Version]
- Roh, K.B.; Ryu, D.H.; Cho, E.; Weon, J.B.; Park, D.; Kweon, D.H.; Jung, E. Coptis chinensis franch directly inhibits proteolytic activation of kallikrein 5 and cathelicidin associated with rosacea in epidermal keratinocytes. Molecules 2020, 25, 5556. [Google Scholar] [CrossRef]
- Enerbäck, L.; Norrby, K. The mast cells. Curr. Top. Pathol. 1989, 79, 169–204. [Google Scholar]
- Pejler, G.; Abrink, M.; Ringvall, M.; Wernersson, S. Mast cell proteases. Adv. Immunol. 2007, 95, 167–255. [Google Scholar]
- Schwartz, L.B.; Irani, A.M.; Roller, K.; Castells, M.C.; Schechter, N.M. Quantitation of histamine, tryptase, and chymase in dispersed human T and TC mast cells. J. Immunol. 1987, 138, 2611–2615. [Google Scholar]
- Wang, L.; Wang, Y.J.; Hao, D.; Wen, X.; Du, D.; He, G.; Jiang, X. The theranostics role of mast cells in the pathophysiology of rosacea. Front. Med. 2020, 6, 324. [Google Scholar] [CrossRef] [Green Version]
- Koch, A.E.; Halloran, M.M.; Haskell, C.J.; Shah, M.R.; Polverini, P.J. Angiogenesis mediated by soluble forms of E-selectin and vascular cell adhesion molecule-1. Nature 1995, 376, 517–519. [Google Scholar] [CrossRef] [PubMed]
- Gho, Y.S.; Kleinman, H.K.; Sosne, G. Angiogenic activity of human soluble intercellular adhesion molecule-1. Cancer Res. 1999, 59, 5128–5132. [Google Scholar] [PubMed]
- Reinholz, M.; Ruzicka, T.; Schauber, J. Cathelicidin LL-37: An antimicrobial peptide with a role in inflammatory skin disease. Ann. Dermatol. 2012, 24, 126–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, P.Y.; Ohtake, T.; Brandt, C.; Strickland, I.; Boguniewicz, M.; Ganz, T.; Gallo, R.L.; Leung, D.Y. Endogenous antimicrobial peptides and skin infections in atopic dermatitis. N. Engl. J. Med. 2002, 347, 1151–1160. [Google Scholar] [CrossRef] [Green Version]
- Nigam, M.; Atanassova, M.; Mishra, A.P.; Pezzani, R.; Devkota, H.P.; Plygun, S.; Salehi, B.; Setzer, W.N.; Sharifi-Rad, J. Bioactive compounds and health benefits of Artemisia species. Nat. Prod. Commun. 2019, 14, 1–17. [Google Scholar]
- Elfawal, M.A.; Towler, M.J.; Reich, N.G.; Golenbock, D.; Weathers, P.J.; Rich, S.M. Dried whole plant Artemisia annua as an antimalarial therapy. PLoS ONE 2012, 7, e52746. [Google Scholar] [CrossRef]
- Yuan, X.; Li, J.; Li, Y.; Deng, Z.; Zhou, L.; Long, J.; Tang, Y.; Zuo, Z.; Zhang, Y.; Xie, H. Artemisinin, a potential option to inhibit inflammation and angiogenesis in rosacea. Biomed. Pharmacother. 2019, 117, 109181. [Google Scholar] [CrossRef]
- Mannan, A.; Ahmed, I.; Arshad, W.; Asim, M.F.; Qureshi, R.A.; Hussain, I.; Mirza, B. Survey of artemisinin production by diverse Artemisia species in northern Pakistan. Malar. J. 2010, 9, 310. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; But, P.P.; Ooi, V.E. Antiviral activity and mode of action of caffeoylquinic acids from Schefflera heptaphylla (L.) frodin. Antivir. Res. 2005, 68, 1–9. [Google Scholar] [CrossRef]
- Carlotto, J.; da Silva, L.M.; Dartora, N.; Maria-Ferreira, D.; Sabry Dde, A.; Filho, A.P.; de Paula Werner, M.F.; Sassaki, G.L.; Gorin, P.A.; Iacomini, M.; et al. Identification of a dicaffeoylquinic acid isomer from Arctium lappa with a potent anti-ulcer activity. Talanta 2015, 135, 50–57. [Google Scholar] [CrossRef]
- Hong, S.; Joo, T.; Jhoo, J.W. Antioxidant and anti-inflammatory activities of 3,5-dicaffeoylquinic acid isolated from Ligularia fischeri leaves. Food Sci. Biotechnol. 2015, 24, 257–263. [Google Scholar] [CrossRef]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; FangFang, X.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Tajik, N.; Tajik, M.; Mack, I.; Enck, P. The potential effects of chlorogenic acid, the main phenolic components in coffee, on health: A comprehensive review of the literature. Eur. J. Nutr. 2017, 56, 2215–2244. [Google Scholar] [CrossRef]
- Murakami, T.; Suzuki, K.; Niyonsaba, F.; Tada, H.; Reich, J.; Tamura, H.; Nagaoka, I. MrgX2-mediated internalization of LL-37 and degranulation of human LAD2 mast cells. Mol. Med. Rep. 2018, 18, 4951–4959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssens, A.S.; Heide, R.; den Hollander, J.C.; Mulder, P.G.; Tank, B.; Oranje, A.P. Mast cell distribution in normal adult skin. J. Clin. Pathol. 2005, 58, 285–289. [Google Scholar] [CrossRef] [Green Version]
- Rothe, M.J.; Nowak, M.; Kerdel, F.A. The mast cell in health and disease. J. Am. Acad. Dermatol. 1990, 23, 615–624. [Google Scholar] [CrossRef]
- Weber, S.; Krüger-Krasagakes, S.; Grabbe, J.; Zuberbier, T.; Czarnetzki, B.M. Mast cells. Int. J. Dermatol. 1995, 34, 1–10. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′-3′) | Product Size (bp) |
|---|---|---|
| Cma1 | Forward TCTGCTGTGTGCTGGGATAG | 190 |
| Reverse GGCACACAAAACCTGCACTA | ||
| Tpsab1 | Forward GTGCTGGGAATGAAGGACAT | 187 |
| Reverse TTGGGGACATAGTGGTGGAT | ||
| ICAM1 | Forward GGCTGGAGCTGTTTGAGAAC | 202 |
| Reverse ACTGTGGGGTTCAACCTCTG | ||
| VCAM1 | Forward CAGACAGGAAGTCCCTGGAA | 212 |
| Reverse TTCTTGCAGCTTTGTGGATG | ||
| FPR2 | Forward CAACCCCATGCTTTACGTCT | 184 |
| Reverse ATATCCCTGACCCCATCCTC | ||
| Mrgprx2 | Forward TGAAAGCAACCATACTGGAATGTC | 113 |
| Reverse ACCACAGCACTGTGGCATTTCC | ||
| GAPDH | Forward TGCACCACCAACTGCTTAGC | 87 |
| Reverse GGCATGGACTGTGGTCATGAG | ||
| Gapdh | Forward CATCACTGCCACCCAGAAGACTG | 153 |
| Reverse ATGCCAGTGAGCTTCCCGTTCAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roh, K.-B.; Jang, Y.; Cho, E.; Park, D.; Kweon, D.-H.; Jung, E. Chlorogenic Acid Isomers Isolated from Artemisia lavandulaefolia Exhibit Anti-Rosacea Effects In Vitro. Biomedicines 2022, 10, 463. https://doi.org/10.3390/biomedicines10020463
Roh K-B, Jang Y, Cho E, Park D, Kweon D-H, Jung E. Chlorogenic Acid Isomers Isolated from Artemisia lavandulaefolia Exhibit Anti-Rosacea Effects In Vitro. Biomedicines. 2022; 10(2):463. https://doi.org/10.3390/biomedicines10020463
Chicago/Turabian StyleRoh, Kyung-Baeg, Youngsu Jang, Eunae Cho, Deokhoon Park, Dae-Hyuk Kweon, and Eunsun Jung. 2022. "Chlorogenic Acid Isomers Isolated from Artemisia lavandulaefolia Exhibit Anti-Rosacea Effects In Vitro" Biomedicines 10, no. 2: 463. https://doi.org/10.3390/biomedicines10020463
APA StyleRoh, K. -B., Jang, Y., Cho, E., Park, D., Kweon, D. -H., & Jung, E. (2022). Chlorogenic Acid Isomers Isolated from Artemisia lavandulaefolia Exhibit Anti-Rosacea Effects In Vitro. Biomedicines, 10(2), 463. https://doi.org/10.3390/biomedicines10020463