
Cross-Talk between Wnt Signaling and Src Tyrosine Kinase


 , and
, and
Abstract
:1. Introduction
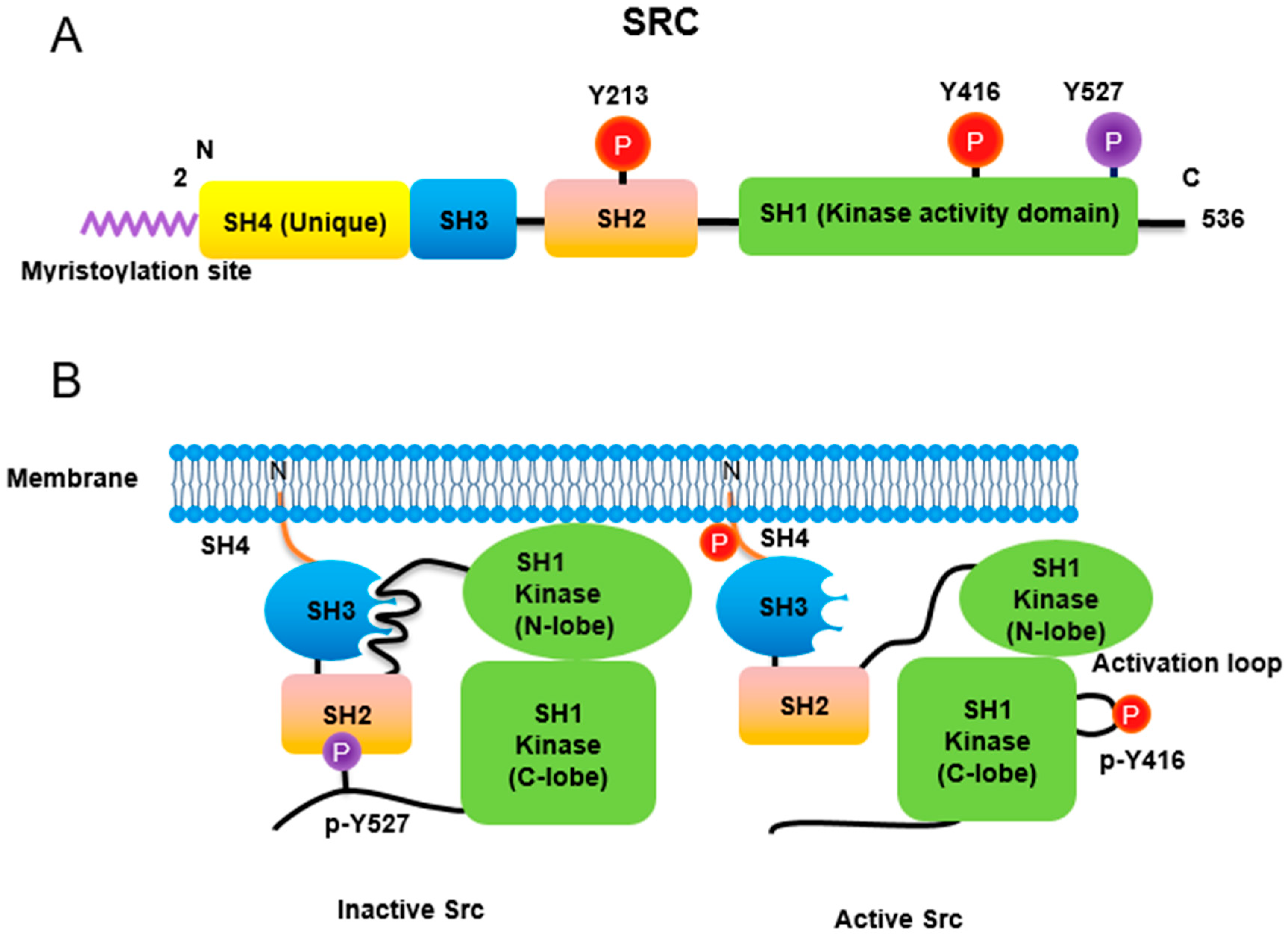
2. Src Non-Receptor Tyrosine Kinase
2.1. Lipidation
2.2. Phosphorylation
2.2.1. Phosphorylation of Tyr416 and Tyr527
2.2.2. Phosphorylation of Ser17, Ser37, Ser69, and Ser75 in the Unique Domain of Src
2.3. Acetylation
2.4. Ubiquitylation
2.5. SUMOylation
2.6. Oxidation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Posttranslational Modification | Description | References |
|---|---|---|
| Lipidation | Myristic acid is covalently linked to Gly2. | [15] |
| The first 14 amino acids of Src contain a recognition sequence for myristoylation of Src | [16] | |
| The initiating Met is cleaved by methionine amino peptidase and Gly2 become to N-terminal amino acid. | [17] | |
| T456A mutation of Src undergoes detached from membrane, suggesting Thr456 is localized in binding pocket and regulates myristoyl switch. | [18] | |
| N-terminal myristoylated SH4 domain interact with SH3 domain when Src in not anchored to a lipid membrane. | [19] | |
| Myristoyl group is particularly essential for high affinity binding to the lipid bilayer via electrostatic interaction with acidic phospholipid in vitro | [20] | |
| SFKs in N-terminal domain and cysteine(s) can be covalently linked to palmitic acid by palmitoyl acyl transferase (PAT). | [17] | |
| Phosphorylation | Tyr416 ensures an active form. | [21] |
| Tyr527 in C-terminal domain reveals inactivation. | [22] | |
| The Tyr527 phosphorylation is carried out by C-terminal Src kinase. | [23] | |
| The Tyr527 phosphorylation is carried out by its homolog Csk homologous kinase (Chk). | [24,25] | |
| Tyr527 of Src promotes assembly SH2 domain of Src, resulting in intramolecular auto-inhibitory Src structure. | [26,27] | |
| PDGF phosphorylates Tyr213 which interferes with p-Tyr527 binding to SH2 domain in Src leading Src activation. | [30] | |
| Three-dimensional structure of Src with phosphorylations of Tyr416 and Tyr527. | [31] | |
| Phosphorylation/dephosphorylation observed in the Unique domain of Src plays a critical regulatory function in Src kinase activity. | [32] | |
| Ser17 of Src was postulated to be a candidate of phosphorylation residue, which likely changes hydrophobicity of Src. | [33] | |
| Ser17 of Src was demonstrated to be phosphorylated by PKA in PC12 cells. Src S17A (dephospho-mimic) inhibits Rap1-dependent ERK activation by NGF and cAMP. | [34] | |
| The phosphorylations of the Thr34, Thr46, and Ser72 residues by p34Cdc2 either sensitize chicken Src to a Tyr527 phosphatase or desensitize Src to a Tyr527 kinase, leading to Src activation. | [35] | |
| The phosphorylation of Ser37 and Ser75 by p25-Cdk5 attenuates lipid binding by the ULBR. | [36,37] | |
| Ser109 phosphorylation of Src undergoes its degradation. | [41] | |
| Wnt3A induces phosphorylation of Src at Ser43, Ser51 and Ser493 residues through p-Ser9 GSK-3β. | [46] | |
| Acetylation | CREB binding protein (CBP) acetylates N-terminal lysine residues (K5, K7 and K9) of c-Src to promote dissociation from the cell membrane. In addition, CBP also acetylates the c-terminal K401, K423, and K427 of c-Src. | [47] |
| Low aggressive osteosarcoma SaOS-2 cells show high level of Src in nucleus. High metastatic 143B osteosarcoma cells present low levels of nuclear Src. | [48] | |
| EGF induces SRC activation, which phosphorylates Tyr845 in EGFR, resulting in mitochondrial localization of Src and EGFR. | [49] | |
| In mitochondria, EGFR binds to cytochrome C oxidase subunit II (CoxII), and EGFR and Src phosphorylate CoxII, leading to decreases of complex IV activity and ATP levels. | [50] | |
| Src is sequestered to mitochondria with AKAP121. | [51] | |
| Overexpression of the downstream of kinase-4 (Dok-4) containing N-terminal mitochondrial targeting sequence increases mitochondrial Src localization through the complex formation with Src. | [52] | |
| Mitochondrial Src is high in breast cancer cells of triple negative subtype, and targets to phosphorylate mitochondrial single stranded DNA-binding protein (SSBP1), a regulator of mtDNA replication. | [53] | |
| Ubiquitylation | p-Tyr419 Src, an active form can undergo Cullin-5-dependent ubiquitylation and activation of Src increases the extent of polyubiquitylation. | [38] |
| Phosphorylation at Ser75 by Cdk5 promotes the ubiquitin-dependent degradation of Src. | [39] | |
| Mutation at Lys429 activates FAK to potentiate Src-induced invasive phenotypes. | [40] | |
| The promoter of FBXL7 is hypermethylated in advanced prostate and pancreatic cancers along with decreased mRNA and protein levels of FBXL7. | [41] | |
| SUMOylation | SUMOylation of Src at K318 negatively modulates its oncogenic function by at least partially, inhibiting Src-FAK complex activity. | [42] |
| Oxidation | Intramolecular disulfide bridge between Cys245 and Cys487 upon exposure to ROS lead to Src activation. | [43] |
| Intermolecular disulfide bridges between Cys277 residues of two different Src proteins result in inactive Src dimers. | [44] | |
| Cytoplasmic Src family kinases are activated gradually by hydrogen peroxide in human aortic endothelial cells (HAECs) and human umbilical vein endothelial cells (HUVECs). | [45] |
3. Localization of Src in Cell Compartments
3.1. Translocation of Src to Nucleus
3.2. Translocation of Src to Mitochondria
4. Wnt Signaling
4.1. Canonical Wnt Signaling
4.2. Non-Canonical Wnt Signaling
4.3. Communication between Canonical and Non-Canonical Pathways of Wnt
4.4. Regulatory Proteins for β-Catenin Activity
4.5. Regulation of β-Catenin Nuclear Translocation
| Proteins Binding to β-Catenin | Description | References |
|---|---|---|
| p-Tyr42 RhoA | Tyrosine 42 phosphorylated form of RhoA (Ras Homolog Family Member A). | [46] |
| Rac/JNK2 | JNK2 phosphorylates β-catenin at Ser191/605 residues. | [68] |
| Bcl9/Lgs | Interaction between β-catenin and BCL9 is mediated by the phosphorylation of β-catenin at Tyrosine 142 residue. | [73,106] |
| Groucho | TLE1 in mammals. | [74,75] |
| CBP (CREB-binding protein) | Prominent histone acetyltransferase (HAT). | [77,78] |
| TBL1 | Transducing β-like protein 1. | [85] |
| Sox | Sry-related HMG box, a key transcriptional factor of animal development. | [89] |
| Sox3 | Sox3 binds to β-catenin, inhibiting TCF activity by competing. | [90] |
| Sox4 | Sox4 enhance β-catenin/TCF activity. | [91] |
| Sox17 | Sox17 binds to β-catenin and regulates gene expression. | [90,91,92] |
| HIF-1α (hypoxia induced factor -1α) | β-catenin enhances HIF-1α-mediated transcription. | [94] |
| hARD1 | hARD1 binds to and acetylates β-catenin, leading to β-catenin activation. | [95] |
| Runx2 | A constitutively active β-catenin enhances LEF1 interaction with and inhibition of Runx2 activity. | [96] |
| PKM2 (pyruvate kinase M2) | PKM2 transactivates β-catenin upon EGF. | [100] |
| FoxM1 (Forkhead box protein M1) | FoxM1 promotes β-catenin nuclear import. | [102,104,105] |
| FOXO (Forkhead box protein O) | ROS induce β-catenin and FOXO interaction. | [103,105] |
| Parafimbromin | Components of the polymerase associated factor 1 (PAF1) complex. | [106] |
| Pygopus | Involved in signal transduction through the Wnt pathway. | [106,107] |
| APC (Adenomatous polyposis coli) | APC is a nuclear-cytoplasmic shuttling protein, and can function as a β-catenin chaperone. | [109] |
| Connexin43 | β-catenin binds to connexin43, p-Tyr265/Tyr313 of connexin43 by Src interfere with the interaction. | [110] |
| HSP27 | Hsp27 interacts with β-catenin, reducing β-catenin-GSK-3β complex. | [111] |
| Kinesin2/IFT-A (intraflagellar Transport A protein) | Kinesin2 promotes nuclear localization of β-catenin during Wnt signaling. | [112] |
| RAPGEF5 (Rap Guanine Nucleotide Exchange Factor 5) | RAPGEF5 binding with Rap1a/b plays a role in β-catenin nuclear import. | [113] |
5. Cross-Talk between Src and Wnt Signaling Pathways
6. Conclusions and Perspectives
| Drug | Descriptions | Cancer Type | Drug Progression | Identifier (NCT Number) | References |
|---|---|---|---|---|---|
| Dasatinib (SPRYCEL®) | Dual inhibitor of Src kinase and Bcr-Abl tyrosine kinase | Philadelphia chromosome-positive Chronic myeloid leukemia (Ph + CML), Philadelphia chromosome-positive acute lymphoblastic leukemia (Ph + ALL) | Approved US (2006) | NCT00064233 | [150] |
| Saracatinib (AZD0530) | Dual inhibitor of Src kinase and Abl family kinases1 | Alzheimer’s disease | Phase 2a completed (2018) | NCT01864655 (Phase 1) NCT02167256 (Phase 2a) | [151,152] |
| Bosutinib (SKI-606) | Dual inhibitor of Src kinase and Abl tyrosine kinase | Ph+ Chronic Myeloid Leukemia | Approved US (2012) | NCT00261846 (Phase 2) | [153,154] |
| Saracatinib (SarCaBon) (AZD0530) | Dual inhibitor of Src kinase and Bcr-Abl tyrosine kinase | Cancer-induced bone pain | Phase 2 completed (2018) | NCT02085603 | |
| KX2-391 (Tirbanibulin) (Klisyri®) | Dual inhibitor of Src kinase and tubulin polymerization | Actinic Keratosis (a precursor of squamous-cell carcinoma) on Face or Scalp | Approved US (2020) | NCT03285477 NCT03285490 | [155] |
| TPX-0046 | Dual inhibitor of RET receptor tyrosine kinase inhibitor and Src kinase | Non-Small Cell Lung Cancer, Medullary Thyroid Cancer, RET Gene Mutation, Metastatic Solid Tumor, Advanced Solid Tumor | Phase 1/2 | NCT04161391 | [156] |
| TPX-0022 (Elzovantinib) | Multi target inhibitor of MET, CSF1R and Src | Non-Small Cell Lung Cancer, Gastric Cancer Advanced Solid Tumor Metastatic Solid Tumors MET Gene Alterations | Phase 1/2 | NCT03993873 |
| Description of Mode of Action | Drugs | Cancer Type | Drug Progression | Identifier (NCT Number) | References |
|---|---|---|---|---|---|
| CBP/β-catenin antagonist | PRI-724 | Advanced pancreatic cancer Metastatic pancreatic cancer Pancreatic adenocarcinoma Advanced solid tumors | Phase 1 | NCT01764477 NCT01302405 NCT01606579 | [157] |
| PRI-724 (with Leucovorin Calcium, Oxaliplatin, or Fluorouracil) | Acute myeloid leukemia Chronic myeloid leukemia | Phase 2 | NCT02413853 | [158] | |
| FZD10 antagonist | OTSA101-DTPA-90Y | Sarcoma Synovial | Phase 1 | NCT01469975 | [159] |
| FZD8 decoy receptor (Fusion protein of FZD8 and IgG Fc domain competing with native FZD8 receptor) | OMP-54F28 | Solid tumors | Phase 1 | NCT01608867 | [160] |
| OMP-54F28 (with Nab-Paclitaxel and Gemcitabine) | Pancreatic cancer Stage IV pancreatic cancer | Phase 1 | NCT02050178 | [161] | |
| OMP-54F28 (with Paclitaxel and Carboplatin) | Ovarian cancer | Phase 1 | NCT02092363 | [162] | |
| OMP-54F28 (with Sorafenib) | Hepatocellular cancer Liver cancer | Phase 1 | NCT02069145 | [163] | |
| Monoclonal antibody against FZD receptors | OMP-18R5 | Metastatic breast cancer Solid tumors | Phase 1 | NCT01973309 NCT01345201 | [164] |
| OMP-18R5 (with Docetaxel) | Solid tumors | Phase 1 | NCT01957007 | [165,166] | |
| OMP-18R5 (with Nab-Paclitaxel and Gemcitabine) | Pancreatic cancer Stage IV pancreatic cancer | Phase 1 | NCT02005315 | [167] | |
| Porcupine inhibitors | CGX1321 | Colorectal adenocarcinoma Gastric adenocarcinoma Pancreatic adenocarcinoma Bile duct carcinoma Hepatocellular carcinoma Esophageal carcinoma Gastrointestinal cancer | Phase 1 | NCT03507998 | [168] |
| CGX1321 (with Pembrolizumab) | Solid tumors GI cancer | Phase 1 | NCT02675946 | [169] | |
| ETC-1922159 | Solid tumor | Phase 1 | NCT02521844 | [170] | |
| RXC004 | Solid tumor | Phase 1 | NCT03447470 | [171] | |
| WNT974 | Squamous cell cancer Head and Neck | Phase 2 | NCT02649530 NCT01351103 | [172] | |
| WNT974 (with LGX818 and Cetuximab) | Metastatic colorectal carcinoma | Phase 1 | NCT02278133 | [173] | |
| β-catenin-controlled gene expression inhibitor | SM08502 | Solid tumors | Phase 1 | NCT03355066 | [174] |
| Wnt signaling pathway inhibitor | SM04690 | Osteoarthritis | Phase 2 | NCT02536833 | [175] |
| SM04755 | Tendinopathy | Phase 1 | NCT03229291 | [176] |
Funding
Conflicts of Interest
Abbreviations
| 15-PGDH | 15-prostaglandin dehydrogenase |
| AKAP121 | A-kinase anchor protein 121 |
| ALL | acute lymphoblastic leukemia |
| APC | Adenomatous polyposis coli |
| ATP | adenosine triphosphate |
| BCL9 | B-cell lymphoma 9 |
| Blk | B lymphocyte kinase |
| CBP | CREB binding protein |
| Cdk5 | Cyclin Dependent Kinase 5 |
| Chk | Csk homologous kinase |
| CK1 | casein kinase 1 |
| CLL | chronic lymphocytic leukemia |
| CML | chronic myelogenous leukemia |
| COX-2 | Cyclooxygenase-2 |
| CoxII | C oxidase subunit II |
| CREB | cAMP-response element binding protein |
| CRIB motifs | Cdc42/Rac interactive binding motifs |
| Csk | C-terminal Src kinase |
| Cx43 | Connexin43 |
| Daam1 | Dishevelled Associated Activator Of Morphogenesis 1 |
| DKK1 | Sclerostin and Dickkopf-related protein 1 |
| DKKs | Dickkopfs |
| Dok-4 | downstream of kinase-4 |
| EGF | epidermal growth factor |
| EGFR | epidermal growth factor receptor |
| ESCC | esophageal squamous cell carcinoma |
| FBXL7 | F-Box And Leucine Rich Repeat Protein 7 |
| Fgr | Gardner-Rasheed feline sarcoma viral oncogene homolog |
| FOXO | Forkhead Box O |
| Frk | fyn-related kinase |
| Fzd | Frizzled receptor |
| GCKR | Glucokinase Regulator |
| GSK-3 | glycogen synthase kinase 3 |
| HAECs | human aortic endothelial cells |
| hARD1 | human ARD1 |
| HAT | histone acetyltransferase |
| Hck | hematopoietic cell kinase |
| HDAC | histone deacetylase |
| Her2 | human epidermal growth factor receptor 2 |
| HIF-1α | hypoxia induced factor-1α |
| Hsp27 | Heat shock protein 27 |
| HUVECs | human umbilical vein endothelial cells |
| IP3 | inositol-3-phosphates |
| JIPs | JNK interacting proteins |
| JNK | c-Jun N-terminal kinases |
| Lck | lymphocyte-specific protein tyrosine kinase |
| Leg | Legless |
| LGR4/5 | Leucine Rich Repeat Containing G Protein-Coupled Receptor 4 and 5 |
| LRP5/6 | low density lipoprotein receptor-related protein (LRP)5 and 6 |
| MAPKK | Mitogen-activated protein kinase |
| MAPKKK | MAPK kinase kinases |
| MEKK1/4 | Mitogen-Activated Protein Kinase Kinase Kinase 1 and 4 |
| MK2 | Mitogen-activated protein kinase-activated protein kinase-2 |
| mtDNA | mitochondrial DNA |
| NF-κB | Nuclear factor-κB |
| NLS | nuclear signaling |
| NMT | N-myristoyl transferase |
| PAF1 | polymerase associated factor 1 |
| PAT | palmitoyl acyl transferase |
| PCP | planar cell polarity |
| PDGF | Platelet-derived growth factor |
| PGE2 | pro-tumorigenic product prostaglandin E2 |
| PGT | prostaglandin transporter |
| PKA | protein kinase A |
| PKC | protein kinase C |
| PKM2 | pyruvate kinase M2 |
| PKN3 | Protein Kinase N3 |
| PLC | phospholipase C |
| PORCN | Porcupine |
| POSH | Plenty of SH3s |
| PPP | Phosphoprotein phosphatase |
| PTEN | phosphatase and tensin homolog |
| PTPD1 | protein tyrosine phosphatase D1 |
| Pyk2 | Proline-rich tyrosine kinase 2 |
| Rac1 | Ras-related C3 botulinum toxin substrate 1 |
| RhoA | Ras Homolog Family Member A |
| Ror1/2 | receptor tyrosine kinase-like orphan receptor 1 and 2 |
| ROS | Reactive Oxygen Species |
| RYK | receptor tyrosine kinase |
| S100a4 | S100 Calcium Binding Protein A4 |
| SFKs | Src family kinases |
| SFRPs | secreted frizzled-related proteins |
| SH domain | Src Homology Domain |
| Sox | SRY-Box Transcription Factor |
| SSBP1 | single stranded DNA-binding protein |
| STAT3 | signal transducer and activator of transcription 3 |
| TAZ | Tafazzin |
| TBL1 | transducing β-like protein 1 |
| TBLR1 | TBL1-related protein 1 |
| TCF/LEF | T cell factor/lymphoid enhancer factor |
| Tiam1 | T-lymphoma invasion and metastasis |
| TNF-α | tumor necrosis factor-α |
| ULBR | unique binding region |
| WGEF | weak-similarity guanine nucleotide-exchange factors |
| WIF-1 | Wnt inhibitory factor-1 |
| Wisp1 | WNT1-inducible-signaling pathway protein 1 |
| Wnt | Wingless and Int-1 |
| WREs | Wnt response element |
| YAP | Yes-associated protein |
| Yes | Yamaguchi sarcoma oncogene |
| Yrk | Yes-related kinase |
| β-TrCP | β-transducin repeat-containing protein |
References
- Simatou, A.; Simatos, G.; Goulielmaki, M.; Spandidos, D.A.; Baliou, S.; Zoumpourlis, V. Historical retrospective of the SRC oncogene and new perspectives (Review). Mol. Clin. Oncol. 2020, 13, 21. [Google Scholar] [CrossRef] [PubMed]
- Stehelin, D.; Varmus, H.E.; Bishop, J.M.; Vogt, P.K. DNA related to the transforming gene(s) of avian sarcoma viruses is present in normal avian DNA. Nature 1976, 260, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Sen, B.; Johnson, F.M. Regulation of SRC family kinases in human cancers. J. Signal. Transduct. 2011, 2011, 865819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oppermann, H.; Levinson, A.D.; Varmus, H.E.; Levintow, L.; Bishop, J.M. Uninfected vertebrate cells contain a protein that is closely related to the product of the avian sarcoma virus transforming gene (src). Proc. Natl. Acad. Sci. USA 1979, 76, 1804–1808. [Google Scholar] [CrossRef] [Green Version]
- Doumpas, N.; Lampart, F.; Robinson, M.D.; Lentini, A.; Nestor, C.E.; Cantu, C.; Basler, K. TCF/LEF dependent and independent transcriptional regulation of Wnt/beta-catenin target genes. EMBO J. 2019, 38. [Google Scholar] [CrossRef]
- Tejeda-Munoz, N.; Robles-Flores, M. Glycogen synthase kinase 3 in Wnt signaling pathway and cancer. IUBMB Life 2015, 67, 914–922. [Google Scholar] [CrossRef]
- Green, J.; Nusse, R.; van Amerongen, R. The role of Ryk and Ror receptor tyrosine kinases in Wnt signal transduction. Cold Spring Harb. Perspect. Biol. 2014, 6, a009175. [Google Scholar] [CrossRef]
- Glinka, A.; Dolde, C.; Kirsch, N.; Huang, Y.L.; Kazanskaya, O.; Ingelfinger, D.; Boutros, M.; Cruciat, C.M.; Niehrs, C. LGR4 and LGR5 are R-spondin receptors mediating Wnt/beta-catenin and Wnt/PCP signalling. EMBO Rep. 2011, 12, 1055–1061. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.A.; Wagle, M.; Tran, K.; Zhan, X.; Dixon, M.A.; Liu, S.; Gros, D.; Korver, W.; Yonkovich, S.; Tomasevic, N.; et al. R-Spondin family members regulate the Wnt pathway by a common mechanism. Mol. Biol. Cell 2008, 19, 2588–2596. [Google Scholar] [CrossRef] [Green Version]
- Yan, K.S.; Janda, C.Y.; Chang, J.; Zheng, G.X.Y.; Larkin, K.A.; Luca, V.C.; Chia, L.A.; Mah, A.T.; Han, A.; Terry, J.M.; et al. Non-equivalence of Wnt and R-spondin ligands during Lgr5(+) intestinal stem-cell self-renewal. Nature 2017, 545, 238–242. [Google Scholar] [CrossRef]
- Martinez-Gil, N.; Ugartondo, N.; Grinberg, D.; Balcells, S. Wnt Pathway Extracellular Components and Their Essential Roles in Bone Homeostasis. Genes 2022, 13, 138. [Google Scholar] [CrossRef] [PubMed]
- Zhan, T.; Rindtorff, N.; Boutros, M. Wnt signaling in cancer. Oncogene 2017, 36, 1461–1473. [Google Scholar] [CrossRef] [PubMed]
- Asem, M.; Young, A.M.; Oyama, C.; Claure De La Zerda, A.; Liu, Y.; Yang, J.; Hilliard, T.S.; Johnson, J.; Harper, E.I.; Guldner, I.; et al. Host Wnt5a Potentiates Microenvironmental Regulation of Ovarian Cancer Metastasis. Cancer Res. 2020, 80, 1156–1170. [Google Scholar] [CrossRef] [PubMed]
- Bagnato, G.; Leopizzi, M.; Urciuoli, E.; Peruzzi, B. Nuclear Functions of the Tyrosine Kinase Src. Int. J. Mol. Sci. 2020, 21, 2675. [Google Scholar] [CrossRef] [Green Version]
- Pellman, D.; Garber, E.A.; Cross, F.R.; Hanafusa, H. An N-terminal peptide from p60src can direct myristylation and plasma membrane localization when fused to heterologous proteins. Nature 1985, 314, 374–377. [Google Scholar] [CrossRef]
- Kamps, M.P.; Buss, J.E.; Sefton, B.M. Mutation of NH2-terminal glycine of p60src prevents both myristoylation and morphological transformation. Proc. Natl. Acad. Sci. USA 1985, 82, 4625–4628. [Google Scholar] [CrossRef] [Green Version]
- Resh, M.D. Fatty acylation of proteins: New insights into membrane targeting of myristoylated and palmitoylated proteins. Biochim. Biophys. Acta 1999, 1451, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Patwardhan, P.; Resh, M.D. Myristoylation and membrane binding regulate c-Src stability and kinase activity. Mol. Cell Biol. 2010, 30, 4094–4107. [Google Scholar] [CrossRef] [Green Version]
- Le Roux, A.L.; Mohammad, I.L.; Mateos, B.; Arbesu, M.; Gairi, M.; Khan, F.A.; Teixeira, J.M.C.; Pons, M. A Myristoyl-Binding Site in the SH3 Domain Modulates c-Src Membrane Anchoring. iScience 2019, 12, 194–203. [Google Scholar] [CrossRef] [Green Version]
- Sigal, C.T.; Zhou, W.; Buser, C.A.; McLaughlin, S.; Resh, M.D. Amino-terminal basic residues of Src mediate membrane binding through electrostatic interaction with acidic phospholipids. Proc. Natl. Acad. Sci. USA 1994, 91, 12253–12257. [Google Scholar] [CrossRef] [Green Version]
- Smart, J.E.; Oppermann, H.; Czernilofsky, A.P.; Purchio, A.F.; Erikson, R.L.; Bishop, J.M. Characterization of sites for tyrosine phosphorylation in the transforming protein of Rous sarcoma virus (pp60v-src) and its normal cellular homologue (pp60c-src). Proc. Natl. Acad. Sci. USA 1981, 78, 6013–6017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, J.A.; King, C.S. Dephosphorylation or antibody binding to the carboxy terminus stimulates pp60c-src. Mol. Cell Biol. 1986, 6, 4467–4477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nada, S.; Okada, M.; MacAuley, A.; Cooper, J.A.; Nakagawa, H. Cloning of a complementary DNA for a protein-tyrosine kinase that specifically phosphorylates a negative regulatory site of p60c-src. Nature 1991, 351, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Hamaguchi, I.; Yamaguchi, N.; Suda, J.; Iwama, A.; Hirao, A.; Hashiyama, M.; Aizawa, S.; Suda, T. Analysis of CSK homologous kinase (CHK/HYL) in hematopoiesis by utilizing gene knockout mice. Biochem. Biophys. Res. Commun. 1996, 224, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Davidson, D.; Chow, L.M.; Veillette, A. Chk, a Csk family tyrosine protein kinase, exhibits Csk-like activity in fibroblasts, but not in an antigen-specific T-cell line. J. Biol. Chem. 1997, 272, 1355–1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weijland, A.; Williams, J.C.; Neubauer, G.; Courtneidge, S.A.; Wierenga, R.K.; Superti-Furga, G. Src regulated by C-terminal phosphorylation is monomeric. Proc. Natl. Acad. Sci. USA 1997, 94, 3590–3595. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Harrison, S.C.; Eck, M.J. Three-dimensional structure of the tyrosine kinase c-Src. Nature 1997, 385, 595–602. [Google Scholar] [CrossRef]
- Su, J.; Muranjan, M.; Sap, J. Receptor protein tyrosine phosphatase alpha activates Src-family kinases and controls integrin-mediated responses in fibroblasts. Curr. Biol. 1999, 9, 505–511. [Google Scholar] [CrossRef] [Green Version]
- Ponniah, S.; Wang, D.Z.; Lim, K.L.; Pallen, C.J. Targeted disruption of the tyrosine phosphatase PTPalpha leads to constitutive downregulation of the kinases Src and Fyn. Curr. Biol. 1999, 9, 535–538. [Google Scholar] [CrossRef] [Green Version]
- Stover, D.R.; Furet, P.; Lydon, N.B. Modulation of the SH2 binding specificity and kinase activity of Src by tyrosine phosphorylation within its SH2 domain. J. Biol. Chem. 1996, 271, 12481–12487. [Google Scholar] [CrossRef] [Green Version]
- Boggon, T.J.; Eck, M.J. Structure and regulation of Src family kinases. Oncogene 2004, 23, 7918–7927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amata, I.; Maffei, M.; Pons, M. Phosphorylation of unique domains of Src family kinases. Front. Genet. 2014, 5, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, F.; deBlaquiere, J.; Burgess, A.W. Translocation of pp60c-src from the plasma membrane to the cytosol after stimulation by platelet-derived growth factor. J. Biol. Chem. 1993, 268, 19552–19558. [Google Scholar] [CrossRef]
- Obara, Y.; Labudda, K.; Dillon, T.J.; Stork, P.J. PKA phosphorylation of Src mediates Rap1 activation in NGF and cAMP signaling in PC12 cells. J. Cell Sci. 2004, 117, 6085–6094. [Google Scholar] [CrossRef] [Green Version]
- Shenoy, S.; Chackalaparampil, I.; Bagrodia, S.; Lin, P.H.; Shalloway, D. Role of p34cdc2-mediated phosphorylations in two-step activation of pp60c-src during mitosis. Proc. Natl. Acad. Sci. USA 1992, 89, 7237–7241. [Google Scholar] [CrossRef] [Green Version]
- Perez, Y.; Gairi, M.; Pons, M.; Bernado, P. Structural characterization of the natively unfolded N-terminal domain of human c-Src kinase: Insights into the role of phosphorylation of the unique domain. J. Mol. Biol. 2009, 391, 136–148. [Google Scholar] [CrossRef]
- Perez, Y.; Maffei, M.; Igea, A.; Amata, I.; Gairi, M.; Nebreda, A.R.; Bernado, P.; Pons, M. Lipid binding by the Unique and SH3 domains of c-Src suggests a new regulatory mechanism. Sci. Rep. 2013, 3, 1295. [Google Scholar] [CrossRef]
- Hakak, Y.; Martin, G.S. Ubiquitin-dependent degradation of active Src. Curr. Biol. 1999, 9, 1039–1042. [Google Scholar] [CrossRef] [Green Version]
- Pan, Q.; Qiao, F.; Gao, C.; Norman, B.; Optican, L.; Zelenka, P.S. Cdk5 targets active Src for ubiquitin-dependent degradation by phosphorylating Src(S75). Cell Mol. Life Sci. 2011, 68, 3425–3436. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Ito, Y.; Kajiwara, K.; Nada, S.; Okada, M. Ubiquitination of Src promotes its secretion via small extracellular vesicles. Biochem. Biophys. Res. Commun. 2020, 525, 184–191. [Google Scholar] [CrossRef]
- Moro, L.; Simoneschi, D.; Kurz, E.; Arbini, A.A.; Jang, S.; Guaragnella, N.; Giannattasio, S.; Wang, W.; Chen, Y.A.; Pires, G.; et al. Epigenetic silencing of the ubiquitin ligase subunit FBXL7 impairs c-SRC degradation and promotes epithelial-to-mesenchymal transition and metastasis. Nat. Cell Biol. 2020, 22, 1130–1142. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Deng, R.; Cui, N.; Zhang, H.; Liu, T.; Dou, J.; Zhao, X.; Chen, R.; Wang, Y.; Yu, J.; et al. Src SUMOylation Inhibits Tumor Growth Via Decreasing FAK Y925 Phosphorylation. Neoplasia 2017, 19, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Giannoni, E.; Buricchi, F.; Raugei, G.; Ramponi, G.; Chiarugi, P. Intracellular reactive oxygen species activate Src tyrosine kinase during cell adhesion and anchorage-dependent cell growth. Mol. Cell Biol. 2005, 25, 6391–6403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemble, D.J.; Sun, G. Direct and specific inactivation of protein tyrosine kinases in the Src and FGFR families by reversible cysteine oxidation. Proc. Natl. Acad. Sci. USA 2009, 106, 5070–5075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, H.; Hao, Q.; Rutherford, S.A.; Low, B.; Zhao, Z.J. Inactivation of SRC family tyrosine kinases by reactive oxygen species in vivo. J. Biol. Chem. 2005, 280, 23918–23925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.G.; Mahmud, S.; Min, J.K.; Lee, Y.B.; Kim, H.; Kang, D.C.; Park, H.S.; Seong, J.; Park, J.B. RhoA GTPase phosphorylated at tyrosine 42 by src kinase binds to beta-catenin and contributes transcriptional regulation of vimentin upon Wnt3A. Redox Biol. 2021, 40, 101842. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Zhang, Z.; Chen, L.; Lee, H.W.; Ayrapetov, M.K.; Zhao, T.C.; Hao, Y.; Gao, J.; Yang, C.; Mehta, G.U.; et al. Acetylation within the N- and C-Terminal Domains of Src Regulates Distinct Roles of STAT3-Mediated Tumorigenesis. Cancer Res. 2018, 78, 2825–2838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urciuoli, E.; Coletta, I.; Rizzuto, E.; De Vito, R.; Petrini, S.; D’Oria, V.; Pezzullo, M.; Milano, G.M.; Cozza, R.; Locatelli, F.; et al. Src nuclear localization and its prognostic relevance in human osteosarcoma. J. Cell Physiol. 2018, 233, 1658–1670. [Google Scholar] [CrossRef]
- Boerner, J.L.; Demory, M.L.; Silva, C.; Parsons, S.J. Phosphorylation of Y845 on the epidermal growth factor receptor mediates binding to the mitochondrial protein cytochrome c oxidase subunit II. Mol. Cell Biol. 2004, 24, 7059–7071. [Google Scholar] [CrossRef] [Green Version]
- Demory, M.L.; Boerner, J.L.; Davidson, R.; Faust, W.; Miyake, T.; Lee, I.; Huttemann, M.; Douglas, R.; Haddad, G.; Parsons, S.J. Epidermal growth factor receptor translocation to the mitochondria: Regulation and effect. J. Biol. Chem. 2009, 284, 36592–36604. [Google Scholar] [CrossRef] [Green Version]
- Cardone, L.; Carlucci, A.; Affaitati, A.; Livigni, A.; DeCristofaro, T.; Garbi, C.; Varrone, S.; Ullrich, A.; Gottesman, M.E.; Avvedimento, E.V.; et al. Mitochondrial AKAP121 binds and targets protein tyrosine phosphatase D1, a novel positive regulator of src signaling. Mol. Cell Biol. 2004, 24, 4613–4626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, S.; Lemay, S.; Osawa, M.; Che, W.; Duan, Y.; Tompkins, A.; Brookes, P.S.; Sheu, S.S.; Abe, J. Mitochondrial Dok-4 recruits Src kinase and regulates NF-kappaB activation in endothelial cells. J. Biol. Chem. 2005, 280, 26383–26396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djeungoue-Petga, M.A.; Lurette, O.; Jean, S.; Hamel-Cote, G.; Martin-Jimenez, R.; Bou, M.; Cannich, A.; Roy, P.; Hebert-Chatelain, E. Intramitochondrial Src kinase links mitochondrial dysfunctions and aggressiveness of breast cancer cells. Cell Death Dis. 2019, 10, 940. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Pan, W. GSK3: A multifaceted kinase in Wnt signaling. Trends Biochem. Sci. 2010, 35, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Piao, S.; Lee, S.H.; Kim, H.; Yum, S.; Stamos, J.L.; Xu, Y.; Lee, S.J.; Lee, J.; Oh, S.; Han, J.K.; et al. Direct inhibition of GSK3beta by the phosphorylated cytoplasmic domain of LRP6 in Wnt/beta-catenin signaling. PLoS ONE 2008, 3, e4046. [Google Scholar] [CrossRef] [Green Version]
- Stamos, J.L.; Chu, M.L.; Enos, M.D.; Shah, N.; Weis, W.I. Structural basis of GSK-3 inhibition by N-terminal phosphorylation and by the Wnt receptor LRP6. Elife 2014, 3, e01998. [Google Scholar] [CrossRef]
- Wu, X.; Yan, T.; Hao, L.; Zhu, Y. Wnt5a induces ROR1 and ROR2 to activate RhoA in esophageal squamous cell carcinoma cells. Cancer Manag. Res. 2019, 11, 2803–2815. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Yan, T.; Li, X.; Sun, J.; Zhang, B.; Wang, H.; Zhu, Y. Daam1 activates RhoA to regulate Wnt5ainduced glioblastoma cell invasion. Oncol. Rep. 2018, 39, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, H.; Yoo, S.K.; Nishita, M.; Kikuchi, A.; Minami, Y. Wnt5a modulates glycogen synthase kinase 3 to induce phosphorylation of receptor tyrosine kinase Ror2. Genes Cells 2007, 12, 1215–1223. [Google Scholar] [CrossRef] [Green Version]
- Hasan, M.K.; Yu, J.; Widhopf, G.F., 2nd; Rassenti, L.Z.; Chen, L.; Shen, Z.; Briggs, S.P.; Neuberg, D.S.; Kipps, T.J. Wnt5a induces ROR1 to recruit DOCK2 to activate Rac1/2 in chronic lymphocytic leukemia. Blood 2018, 132, 170–178. [Google Scholar] [CrossRef] [Green Version]
- Corda, G.; Sala, A. Non-canonical WNT/PCP signalling in cancer: Fzd6 takes centre stage. Oncogenesis 2017, 6, e364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores-Hernandez, E.; Velazquez, D.M.; Castaneda-Patlan, M.C.; Fuentes-Garcia, G.; Fonseca-Camarillo, G.; Yamamoto-Furusho, J.K.; Romero-Avila, M.T.; Garcia-Sainz, J.A.; Robles-Flores, M. Canonical and non-canonical Wnt signaling are simultaneously activated by Wnts in colon cancer cells. Cell Signal. 2020, 72, 109636. [Google Scholar] [CrossRef] [PubMed]
- Tanegashima, K.; Zhao, H.; Dawid, I.B. WGEF activates Rho in the Wnt-PCP pathway and controls convergent extension in Xenopus gastrulation. EMBO J. 2008, 27, 606–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kratzer, M.C.; Becker, S.F.S.; Grund, A.; Merks, A.; Harnos, J.; Bryja, V.; Giehl, K.; Kashef, J.; Borchers, A. The Rho guanine nucleotide exchange factor Trio is required for neural crest cell migration and interacts with Dishevelled. Development 2020, 147. [Google Scholar] [CrossRef]
- Coso, O.A.; Chiariello, M.; Yu, J.C.; Teramoto, H.; Crespo, P.; Xu, N.; Miki, T.; Gutkind, J.S. The small GTP-binding proteins Rac1 and Cdc42 regulate the activity of the JNK/SAPK signaling pathway. Cell 1995, 81, 1137–1146. [Google Scholar] [CrossRef] [Green Version]
- Girardin, S.E.; Yaniv, M. A direct interaction between JNK1 and CrkII is critical for Rac1-induced JNK activation. EMBO J. 2001, 20, 3437–3446. [Google Scholar] [CrossRef] [Green Version]
- Kyriakis, J.M.; Avruch, J. Mammalian MAPK signal transduction pathways activated by stress and inflammation: A 10-year update. Physiol. Rev. 2012, 92, 689–737. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Tu, X.; Joeng, K.S.; Hilton, M.J.; Williams, D.A.; Long, F. Rac1 activation controls nuclear localization of beta-catenin during canonical Wnt signaling. Cell 2008, 133, 340–353. [Google Scholar] [CrossRef] [Green Version]
- Jones, W.M.; Bejsovec, A. RacGap50C negatively regulates wingless pathway activity during Drosophila embryonic development. Genetics 2005, 169, 2075–2086. [Google Scholar] [CrossRef] [Green Version]
- Buongiorno, P.; Pethe, V.V.; Charames, G.S.; Esufali, S.; Bapat, B. Rac1 GTPase and the Rac1 exchange factor Tiam1 associate with Wnt-responsive promoters to enhance beta-catenin/TCF-dependent transcription in colorectal cancer cells. Mol. Cancer 2008, 7, 73. [Google Scholar] [CrossRef] [Green Version]
- Habas, R.; Dawid, I.B.; He, X. Coactivation of Rac and Rho by Wnt/Frizzled signaling is required for vertebrate gastrulation. Genes Dev. 2003, 17, 295–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bikkavilli, R.K.; Feigin, M.E.; Malbon, C.C. G alpha o mediates WNT-JNK signaling through dishevelled 1 and 3, RhoA family members, and MEKK 1 and 4 in mammalian cells. J. Cell Sci. 2008, 121, 234–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anthony, C.C.; Robbins, D.J.; Ahmed, Y.; Lee, E. Nuclear Regulation of Wnt/beta-Catenin Signaling: It’s a Complex Situation. Genes 2020, 11, 886. [Google Scholar] [CrossRef] [PubMed]
- Cavallo, R.A.; Cox, R.T.; Moline, M.M.; Roose, J.; Polevoy, G.A.; Clevers, H.; Peifer, M.; Bejsovec, A. Drosophila Tcf and Groucho interact to repress Wingless signalling activity. Nature 1998, 395, 604–608. [Google Scholar] [CrossRef] [PubMed]
- Roose, J.; Molenaar, M.; Peterson, J.; Hurenkamp, J.; Brantjes, H.; Moerer, P.; van de Wetering, M.; Destree, O.; Clevers, H. The Xenopus Wnt effector XTcf-3 interacts with Groucho-related transcriptional repressors. Nature 1998, 395, 608–612. [Google Scholar] [CrossRef] [PubMed]
- Daniels, D.L.; Weis, W.I. Beta-catenin directly displaces Groucho/TLE repressors from Tcf/Lef in Wnt-mediated transcription activation. Nat. Struct. Mol. Biol. 2005, 12, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Sutter, C.; Parker, D.S.; Blauwkamp, T.; Fang, M.; Cadigan, K.M. CBP/p300 are bimodal regulators of Wnt signaling. EMBO J. 2007, 26, 2284–2294. [Google Scholar] [CrossRef] [Green Version]
- Mosimann, C.; Hausmann, G.; Basler, K. Beta-catenin hits chromatin: Regulation of Wnt target gene activation. Nat. Rev. Mol. Cell Biol. 2009, 10, 276–286. [Google Scholar] [CrossRef]
- Levy, L.; Wei, Y.; Labalette, C.; Wu, Y.; Renard, C.A.; Buendia, M.A.; Neuveut, C. Acetylation of beta-catenin by p300 regulates beta-catenin-Tcf4 interaction. Mol. Cell Biol. 2004, 24, 3404–3414. [Google Scholar] [CrossRef] [Green Version]
- Evans, P.M.; Chen, X.; Zhang, W.; Liu, C. KLF4 interacts with beta-catenin/TCF4 and blocks p300/CBP recruitment by beta-catenin. Mol. Cell Biol. 2010, 30, 372–381. [Google Scholar] [CrossRef] [Green Version]
- Wolf, D.; Rodova, M.; Miska, E.A.; Calvet, J.P.; Kouzarides, T. Acetylation of beta-catenin by CREB-binding protein (CBP). J. Biol. Chem. 2002, 277, 25562–25567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chocarro-Calvo, A.; Garcia-Martinez, J.M.; Ardila-Gonzalez, S.; De la Vieja, A.; Garcia-Jimenez, C. Glucose-induced beta-catenin acetylation enhances Wnt signaling in cancer. Mol. Cell 2013, 49, 474–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Wu, H.; Huang, X.; Jian, Y.; Kong, L.; Xu, H.; Ouyang, Y.; Chen, X.; Wu, G.; Yu, L.; et al. BOP1 confers chemoresistance of triple-negative breast cancer by promoting CBP-mediated beta-catenin acetylation. J. Pathol. 2021, 254, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Olivier-Van Stichelen, S.; Dehennaut, V.; Buzy, A.; Zachayus, J.L.; Guinez, C.; Mir, A.M.; El Yazidi-Belkoura, I.; Copin, M.C.; Boureme, D.; Loyaux, D.; et al. O-GlcNAcylation stabilizes beta-catenin through direct competition with phosphorylation at threonine 41. FASEB J. 2014, 28, 3325–3338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Wang, C.Y. TBL1-TBLR1 and beta-catenin recruit each other to Wnt target-gene promoter for transcription activation and oncogenesis. Nat. Cell Biol. 2008, 10, 160–169. [Google Scholar] [CrossRef]
- Stadeli, R.; Basler, K. Dissecting nuclear Wingless signalling: Recruitment of the transcriptional co-activator Pygopus by a chain of adaptor proteins. Mech. Dev. 2005, 122, 1171–1182. [Google Scholar] [CrossRef]
- Carrera, I.; Janody, F.; Leeds, N.; Duveau, F.; Treisman, J.E. Pygopus activates Wingless target gene transcription through the mediator complex subunits Med12 and Med13. Proc. Natl. Acad. Sci. USA 2008, 105, 6644–6649. [Google Scholar] [CrossRef] [Green Version]
- Kessler, R.; Hausmann, G.; Basler, K. The PHD domain is required to link Drosophila Pygopus to Legless/beta-catenin and not to histone H3. Mech. Dev. 2009, 126, 752–759. [Google Scholar] [CrossRef]
- Sarkar, A.; Hochedlinger, K. The sox family of transcription factors: Versatile regulators of stem and progenitor cell fate. Cell Stem. Cell 2013, 12, 15–30. [Google Scholar] [CrossRef] [Green Version]
- Zorn, A.M.; Barish, G.D.; Williams, B.O.; Lavender, P.; Klymkowsky, M.W.; Varmus, H.E. Regulation of Wnt signaling by Sox proteins: XSox17 alpha/beta and XSox3 physically interact with beta-catenin. Mol. Cell 1999, 4, 487–498. [Google Scholar] [CrossRef]
- Sinner, D.; Kordich, J.J.; Spence, J.R.; Opoka, R.; Rankin, S.; Lin, S.C.; Jonatan, D.; Zorn, A.M.; Wells, J.M. Sox17 and Sox4 differentially regulate beta-catenin/T-cell factor activity and proliferation of colon carcinoma cells. Mol. Cell Biol. 2007, 27, 7802–7815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, S.; Chaturvedi, P.; Rankin, S.A.; Fish, M.B.; Wlizla, M.; Paraiso, K.D.; MacDonald, M.; Chen, X.; Weirauch, M.T.; Blitz, I.L.; et al. Sox17 and beta-catenin co-occupy Wnt-responsive enhancers to govern the endoderm gene regulatory network. Elife 2020, 9, e58029. [Google Scholar] [CrossRef] [PubMed]
- Hill, T.P.; Spater, D.; Taketo, M.M.; Birchmeier, W.; Hartmann, C. Canonical Wnt/beta-catenin signaling prevents osteoblasts from differentiating into chondrocytes. Dev. Cell 2005, 8, 727–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaidi, A.; Williams, A.C.; Paraskeva, C. Interaction between beta-catenin and HIF-1 promotes cellular adaptation to hypoxia. Nat. Cell Biol. 2007, 9, 210–217. [Google Scholar] [CrossRef]
- Lim, J.H.; Chun, Y.S.; Park, J.W. Hypoxia-inducible factor-1alpha obstructs a Wnt signaling pathway by inhibiting the hARD1-mediated activation of beta-catenin. Cancer Res. 2008, 68, 5177–5184. [Google Scholar] [CrossRef] [Green Version]
- Kahler, R.A.; Westendorf, J.J. Lymphoid enhancer factor-1 and beta-catenin inhibit Runx2-dependent transcriptional activation of the osteocalcin promoter. J. Biol. Chem. 2003, 278, 11937–11944. [Google Scholar] [CrossRef] [Green Version]
- Smartt, H.J.; Greenhough, A.; Ordonez-Moran, P.; Talero, E.; Cherry, C.A.; Wallam, C.A.; Parry, L.; Al Kharusi, M.; Roberts, H.R.; Mariadason, J.M.; et al. beta-catenin represses expression of the tumour suppressor 15-prostaglandin dehydrogenase in the normal intestinal epithelium and colorectal tumour cells. Gut 2012, 61, 1306–1314. [Google Scholar] [CrossRef]
- Lilien, J.; Balsamo, J. The regulation of cadherin-mediated adhesion by tyrosine phosphorylation/dephosphorylation of beta-catenin. Curr. Opin. Cell Biol. 2005, 17, 459–465. [Google Scholar] [CrossRef]
- Sayat, R.; Leber, B.; Grubac, V.; Wiltshire, L.; Persad, S. O-GlcNAc-glycosylation of beta-catenin regulates its nuclear localization and transcriptional activity. Exp. Cell Res. 2008, 314, 2774–2787. [Google Scholar] [CrossRef]
- Yang, W.; Xia, Y.; Ji, H.; Zheng, Y.; Liang, J.; Huang, W.; Gao, X.; Aldape, K.; Lu, Z. Nuclear PKM2 regulates beta-catenin transactivation upon EGFR activation. Nature 2011, 480, 118–122. [Google Scholar] [CrossRef]
- Ma, R.Y.; Tong, T.H.; Cheung, A.M.; Tsang, A.C.; Leung, W.Y.; Yao, K.M. Raf/MEK/MAPK signaling stimulates the nuclear translocation and transactivating activity of FOXM1c. J. Cell Sci. 2005, 118, 795–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, N.; Wei, P.; Gong, A.; Chiu, W.T.; Lee, H.T.; Colman, H.; Huang, H.; Xue, J.; Liu, M.; Wang, Y.; et al. FoxM1 promotes beta-catenin nuclear localization and controls Wnt target-gene expression and glioma tumorigenesis. Cancer Cell 2011, 20, 427–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Essers, M.A.; de Vries-Smits, L.M.; Barker, N.; Polderman, P.E.; Burgering, B.M.; Korswagen, H.C. Functional interaction between beta-catenin and FOXO in oxidative stress signaling. Science 2005, 308, 1181–1184. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, Y.; Xue, J.; Gong, A.; Yu, G.; Zhou, A.; Lin, K.; Zhang, S.; Zhang, N.; Gottardi, C.J.; et al. Wnt-induced deubiquitination FoxM1 ensures nucleus beta-catenin transactivation. EMBO J. 2016, 35, 668–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, S. Regulation of Wnt Signaling by FOX Transcription Factors in Cancer. Cancers 2021, 13, 3446. [Google Scholar] [CrossRef]
- Mosimann, C.; Hausmann, G.; Basler, K. Parafibromin/Hyrax activates Wnt/Wg target gene transcription by direct association with beta-catenin/Armadillo. Cell 2006, 125, 327–341. [Google Scholar] [CrossRef] [Green Version]
- Townsley, F.M.; Cliffe, A.; Bienz, M. Pygopus and Legless target Armadillo/beta-catenin to the nucleus to enable its transcriptional co-activator function. Nat. Cell Biol. 2004, 6, 626–633. [Google Scholar] [CrossRef]
- Dubash, A.D.; Guilluy, C.; Srougi, M.C.; Boulter, E.; Burridge, K.; Garcia-Mata, R. The small GTPase RhoA localizes to the nucleus and is activated by Net1 and DNA damage signals. PLoS ONE 2011, 6, e17380. [Google Scholar] [CrossRef] [Green Version]
- Henderson, B.R. Nuclear-cytoplasmic shuttling of APC regulates beta-catenin subcellular localization and turnover. Nat. Cell Biol. 2000, 2, 653–660. [Google Scholar] [CrossRef]
- Spagnol, G.; Trease, A.J.; Zheng, L.; Gutierrez, M.; Basu, I.; Sarmiento, C.; Moore, G.; Cervantes, M.; Sorgen, P.L. Connexin43 Carboxyl-Terminal Domain Directly Interacts with beta-Catenin. Int. J. Mol. Sci. 2018, 19, 1562. [Google Scholar] [CrossRef] [Green Version]
- Cordonnier, T.; Bishop, J.L.; Shiota, M.; Nip, K.M.; Thaper, D.; Vahid, S.; Heroux, D.; Gleave, M.; Zoubeidi, A. Hsp27 regulates EGF/beta-catenin mediated epithelial to mesenchymal transition in prostate cancer. Int. J. Cancer 2015, 136, E496–E507. [Google Scholar] [CrossRef] [PubMed]
- Vuong, L.T.; Iomini, C.; Balmer, S.; Esposito, D.; Aaronson, S.A.; Mlodzik, M. Kinesin-2 and IFT-A act as a complex promoting nuclear localization of beta-catenin during Wnt signalling. Nat. Commun. 2018, 9, 5304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffin, J.N.; Del Viso, F.; Duncan, A.R.; Robson, A.; Hwang, W.; Kulkarni, S.; Liu, K.J.; Khokha, M.K. RAPGEF5 Regulates Nuclear Translocation of beta-Catenin. Dev. Cell 2018, 44, 248–260 e244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, N.; Malbon, C.C. Dishevelled-2 docks and activates Src in a Wnt-dependent manner. J. Cell Sci. 2009, 122, 4439–4451. [Google Scholar] [CrossRef] [Green Version]
- Villarroel, A.; Del Valle-Perez, B.; Fuertes, G.; Curto, J.; Ontiveros, N.; Garcia de Herreros, A.; Dunach, M. Src and Fyn define a new signaling cascade activated by canonical and non-canonical Wnt ligands and required for gene transcription and cell invasion. Cell Mol. Life Sci. 2020, 77, 919–935. [Google Scholar] [CrossRef]
- Kuo, T.L.; Cheng, K.H.; Shan, Y.S.; Chen, L.T.; Hung, W.C. beta-catenin-activated autocrine PDGF/Src signaling is a therapeutic target in pancreatic cancer. Theranostics 2019, 9, 324–336. [Google Scholar] [CrossRef]
- Chen, Q.; Su, Y.; Wesslowski, J.; Hagemann, A.I.; Ramialison, M.; Wittbrodt, J.; Scholpp, S.; Davidson, G. Tyrosine phosphorylation of LRP6 by Src and Fer inhibits Wnt/beta-catenin signalling. EMBO Rep. 2014, 15, 1254–1267. [Google Scholar] [CrossRef] [Green Version]
- Gayrard, C.; Bernaudin, C.; Dejardin, T.; Seiler, C.; Borghi, N. Src- and confinement-dependent FAK activation causes E-cadherin relaxation and beta-catenin activity. J. Cell Biol. 2018, 217, 1063–1077. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Liu, X.; Deng, R.; Gao, S.; Yu, H.; Huang, K.; Jiang, Q.; Liu, R.; Li, X.; Zhang, L.; et al. Nintedanib Inhibits Wnt3a-Induced Myofibroblast Activation by Suppressing the Src/beta-Catenin Pathway. Front. Pharmacol. 2020, 11, 310. [Google Scholar] [CrossRef]
- Miravet, S.; Piedra, J.; Castano, J.; Raurell, I.; Franci, C.; Dunach, M.; Garcia de Herreros, A. Tyrosine phosphorylation of plakoglobin causes contrary effects on its association with desmosomes and adherens junction components and modulates beta-catenin-mediated transcription. Mol. Cell Biol. 2003, 23, 7391–7402. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.K.; Wei, Y.; Szekeres, C.; Kugler, M.C.; Wolters, P.J.; Hill, M.L.; Frank, J.A.; Brumwell, A.N.; Wheeler, S.E.; Kreidberg, J.A.; et al. Epithelial cell alpha3beta1 integrin links beta-catenin and Smad signaling to promote myofibroblast formation and pulmonary fibrosis. J. Clin. Investig. 2009, 119, 213–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiota, M.; Bishop, J.L.; Nip, K.M.; Zardan, A.; Takeuchi, A.; Cordonnier, T.; Beraldi, E.; Bazov, J.; Fazli, L.; Chi, K.; et al. Hsp27 regulates epithelial mesenchymal transition, metastasis, and circulating tumor cells in prostate cancer. Cancer Res. 2013, 73, 3109–3119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wang, G.; Cheng, D.; Huang, S.; Chang, A.; Tan, X.; Wang, Q.; Zhao, S.; Wu, D.; Liu, A.T.; et al. Her2 promotes early dissemination of breast cancer by suppressing the p38-MK2-Hsp27 pathway that is targetable by Wip1 inhibition. Oncogene 2020, 39, 6313–6326. [Google Scholar] [CrossRef] [PubMed]
- Herrero-Gonzalez, S.; Gangoso, E.; Giaume, C.; Naus, C.C.; Medina, J.M.; Tabernero, A. Connexin43 inhibits the oncogenic activity of c-Src in C6 glioma cells. Oncogene 2010, 29, 5712–5723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Sanchez, A.; Jaraiz-Rodriguez, M.; Dominguez-Prieto, M.; Herrero-Gonzalez, S.; Medina, J.M.; Tabernero, A. Connexin43 recruits PTEN and Csk to inhibit c-Src activity in glioma cells and astrocytes. Oncotarget 2016, 7, 49819–49833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talaveron, R.; Matarredona, E.R.; Herrera, A.; Medina, J.M.; Tabernero, A. Connexin43 Region 266-283, via Src Inhibition, Reduces Neural Progenitor Cell Proliferation Promoted by EGF and FGF-2 and Increases Astrocytic Differentiation. Int. J. Mol. Sci. 2020, 21, 8852. [Google Scholar] [CrossRef] [PubMed]
- Azzolin, L.; Panciera, T.; Soligo, S.; Enzo, E.; Bicciato, S.; Dupont, S.; Bresolin, S.; Frasson, C.; Basso, G.; Guzzardo, V.; et al. YAP/TAZ incorporation in the beta-catenin destruction complex orchestrates the Wnt response. Cell 2014, 158, 157–170. [Google Scholar] [CrossRef] [Green Version]
- Matozaki, T.; Kotani, T.; Murata, Y.; Saito, Y. Roles of Src family kinase, Ras, and mTOR signaling in intestinal epithelial homeostasis and tumorigenesis. Cancer Sci. 2020, 112, 16–21. [Google Scholar] [CrossRef]
- Barry, E.R.; Morikawa, T.; Butler, B.L.; Shrestha, K.; de la Rosa, R.; Yan, K.S.; Fuchs, C.S.; Magness, S.T.; Smits, R.; Ogino, S.; et al. Restriction of intestinal stem cell expansion and the regenerative response by YAP. Nature 2013, 493, 106–110. [Google Scholar] [CrossRef]
- Imajo, M.; Miyatake, K.; Iimura, A.; Miyamoto, A.; Nishida, E. A molecular mechanism that links Hippo signalling to the inhibition of Wnt/beta-catenin signalling. EMBO J. 2012, 31, 1109–1122. [Google Scholar] [CrossRef] [Green Version]
- Ikeuchi, M.; Yuki, R.; Saito, Y.; Nakayama, Y. The tumor suppressor LATS2 reduces v-Src-induced membrane blebs in a kinase activity-independent manner. FASEB J. 2021, 35, e21242. [Google Scholar] [CrossRef] [PubMed]
- Guillermin, O.; Angelis, N.; Sidor, C.M.; Ridgway, R.; Baulies, A.; Kucharska, A.; Antas, P.; Rose, M.R.; Cordero, J.; Sansom, O.; et al. Wnt and Src signals converge on YAP-TEAD to drive intestinal regeneration. EMBO J. 2021, 40, e105770. [Google Scholar] [CrossRef] [PubMed]
- Thorup, A.S.; Strachan, D.; Caxaria, S.; Poulet, B.; Thomas, B.L.; Eldridge, S.E.; Nalesso, G.; Whiteford, J.R.; Pitzalis, C.; Aigner, T.; et al. ROR2 blockade as a therapy for osteoarthritis. Sci. Transl. Med. 2020, 12, eaax3063. [Google Scholar] [CrossRef] [PubMed]
- Baron, R.; Kneissel, M. WNT signaling in bone homeostasis and disease: From human mutations to treatments. Nat. Med. 2013, 19, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Holmen, S.L.; Giambernardi, T.A.; Zylstra, C.R.; Buckner-Berghuis, B.D.; Resau, J.H.; Hess, J.F.; Glatt, V.; Bouxsein, M.L.; Ai, M.; Warman, M.L.; et al. Decreased BMD and limb deformities in mice carrying mutations in both Lrp5 and Lrp6. J. Bone Miner. Res. 2004, 19, 2033–2040. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, V.; Bryant, H.U.; Macdougald, O.A. Regulation of bone mass by Wnt signaling. J. Clin. Investig. 2006, 116, 1202–1209. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, P.; Liu, W.; Maye, P.; Zhang, J.; Zhang, Y.; Hurley, M.; Guo, C.; Boskey, A.; Sun, L.; et al. Dkk2 has a role in terminal osteoblast differentiation and mineralized matrix formation. Nat. Genet. 2005, 37, 945–952. [Google Scholar] [CrossRef]
- Uehara, S.; Udagawa, N.; Mukai, H.; Ishihara, A.; Maeda, K.; Yamashita, T.; Murakami, K.; Nishita, M.; Nakamura, T.; Kato, S.; et al. Protein kinase N3 promotes bone resorption by osteoclasts in response to Wnt5a-Ror2 signaling. Sci. Signal. 2017, 10. [Google Scholar] [CrossRef] [Green Version]
- Uehara, S.; Udagawa, N.; Kobayashi, Y. Regulation of osteoclast function via Rho-Pkn3-c-Src pathways. J. Oral. Biosci. 2019, 61, 135–140. [Google Scholar] [CrossRef]
- Lai, S.S.; Xue, B.; Yang, Y.; Zhao, L.; Chu, C.S.; Hao, J.Y.; Wen, C.J. Ror2-Src signaling in metastasis of mouse melanoma cells is inhibited by NRAGE. Cancer Genet. 2012, 205, 552–562. [Google Scholar] [CrossRef]
- Goc, A.; Al-Husein, B.; Katsanevas, K.; Steinbach, A.; Lou, U.; Sabbineni, H.; DeRemer, D.L.; Somanath, P.R. Targeting Src-mediated Tyr216 phosphorylation and activation of GSK-3 in prostate cancer cells inhibit prostate cancer progression in vitro and in vivo. Oncotarget 2014, 5, 775–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keating, G.M. Dasatinib: A Review in Chronic Myeloid Leukaemia and Ph+ Acute Lymphoblastic Leukaemia. Drugs 2017, 77, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Gavert, N.; Zwang, Y.; Weiser, R.; Greenberg, O.; Halperin, S.; Jacobi, O.; Mallel, G.; Sandler, O.; Berger, A.J.; Stossel, E.; et al. Ex vivo organotypic cultures for synergistic therapy prioritization identify patient-specific responses to combined MEK and Src inhibition in colorectal cancer. Nat. Cancer 2022, 3, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Simpkins, F.; Jang, K.; Yoon, H.; Hew, K.E.; Kim, M.; Azzam, D.J.; Sun, J.; Zhao, D.; Ince, T.A.; Liu, W.; et al. Dual Src and MEK Inhibition Decreases Ovarian Cancer Growth and Targets Tumor Initiating Stem-Like Cells. Clin. Cancer Res. 2018, 24, 4874–4886. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Kodithuwakku, S.P.; Ng, P.Y.; Chai, J.; Ng, E.H.; Yeung, W.S.; Ho, P.C.; Lee, K.F. Excessive ovarian stimulation up-regulates the Wnt-signaling molecule DKK1 in human endometrium and may affect implantation: An in vitro co-culture study. Hum. Reprod. 2010, 25, 479–490. [Google Scholar] [CrossRef] [Green Version]
- Bafico, A.; Gazit, A.; Pramila, T.; Finch, P.W.; Yaniv, A.; Aaronson, S.A. Interaction of frizzled related protein (FRP) with Wnt ligands and the frizzled receptor suggests alternative mechanisms for FRP inhibition of Wnt signaling. J. Biol. Chem. 1999, 274, 16180–16187. [Google Scholar] [CrossRef] [Green Version]
- Jung, Y.S.; Park, J.I. Wnt signaling in cancer: Therapeutic targeting of Wnt signaling beyond beta-catenin and the destruction complex. Exp. Mol. Med. 2020, 52, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Y, K.N.; Perumalsamy, N.K.; Warrier, S.; Perumalsamy, L.R.; Dharmarajan, A. Wnt antagonist as therapeutic targets in ovarian cancer. Int. J. Biochem. Cell Biol. 2022, 145, 106191. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X. Targeting the Wnt/beta-catenin signaling pathway in cancer. J. Hematol. Oncol. 2020, 13, 165. [Google Scholar] [CrossRef]
- Talpaz, M.; Shah, N.P.; Kantarjian, H.; Donato, N.; Nicoll, J.; Paquette, R.; Cortes, J.; O’Brien, S.; Nicaise, C.; Bleickardt, E.; et al. Dasatinib in imatinib-resistant Philadelphia chromosome-positive leukemias. N. Engl. J. Med. 2006, 354, 2531–2541. [Google Scholar] [CrossRef] [Green Version]
- Nygaard, H.B.; Wagner, A.F.; Bowen, G.S.; Good, S.P.; MacAvoy, M.G.; Strittmatter, K.A.; Kaufman, A.C.; Rosenberg, B.J.; Sekine-Konno, T.; Varma, P.; et al. A phase Ib multiple ascending dose study of the safety, tolerability, and central nervous system availability of AZD0530 (saracatinib) in Alzheimer’s disease. Alzheimers Res. Ther. 2015, 7, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Dyck, C.H.; Nygaard, H.B.; Chen, K.; Donohue, M.C.; Raman, R.; Rissman, R.A.; Brewer, J.B.; Koeppe, R.A.; Chow, T.W.; Rafii, M.S.; et al. Effect of AZD0530 on Cerebral Metabolic Decline in Alzheimer Disease: A Randomized Clinical Trial. JAMA Neurol. 2019, 76, 1219–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortes, J.E.; Kantarjian, H.M.; Brummendorf, T.H.; Kim, D.W.; Turkina, A.G.; Shen, Z.X.; Pasquini, R.; Khoury, H.J.; Arkin, S.; Volkert, A.; et al. Safety and efficacy of bosutinib (SKI-606) in chronic phase Philadelphia chromosome-positive chronic myeloid leukemia patients with resistance or intolerance to imatinib. Blood 2011, 118, 4567–4576. [Google Scholar] [CrossRef] [Green Version]
- Gambacorti-Passerini, C.; Brummendorf, T.H.; Kim, D.W.; Turkina, A.G.; Masszi, T.; Assouline, S.; Durrant, S.; Kantarjian, H.M.; Khoury, H.J.; Zaritskey, A.; et al. Bosutinib efficacy and safety in chronic phase chronic myeloid leukemia after imatinib resistance or intolerance: Minimum 24-month follow-up. Am. J. Hematol. 2014, 89, 732–742. [Google Scholar] [CrossRef] [PubMed]
- Blauvelt, A.; Kempers, S.; Lain, E.; Schlesinger, T.; Tyring, S.; Forman, S.; Ablon, G.; Martin, G.; Wang, H.; Cutler, D.L.; et al. Phase 3 Trials of Tirbanibulin Ointment for Actinic Keratosis. N. Engl. J. Med. 2021, 384, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Oki, E.; Makiyama, A.; Miyamoto, Y.; Kotaka, M.; Kawanaka, H.; Miwa, K.; Kabashima, A.; Noguchi, T.; Yuge, K.; Kashiwada, T.; et al. Trifluridine/tipiracil plus bevacizumab as a first-line treatment for elderly patients with metastatic colorectal cancer (KSCC1602): A multicenter phase II trial. Cancer Med. 2021, 10, 454–461. [Google Scholar] [CrossRef]
- Moroney, M.R.; Woodruff, E.; Qamar, L.; Bradford, A.P.; Wolsky, R.; Bitler, B.G.; Corr, B.R. Inhibiting Wnt/beta-catenin in CTNNB1-mutated endometrial cancer. Mol. Carcinog. 2021, 60, 511–523. [Google Scholar] [CrossRef]
- Danesh Pouya, F.; Rasmi, Y.; Nemati, M. Signaling Pathways Involved in 5-FU Drug Resistance in Cancer. Cancer Investig. 2022, 1–28. [Google Scholar] [CrossRef]
- Goswami, V.G.; Patel, B.D. Recent updates on Wnt signaling modulators: A patent review (2014-2020). Expert Opin. Ther. Pat. 2021, 31, 1009–1043. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, W.; Zhao, C. Frizzled Receptors in Tumors, Focusing on Signaling, Roles, Modulation Mechanisms, and Targeted Therapies. Oncol. Res. 2021, 28, 661–674. [Google Scholar] [CrossRef]
- Dotan, E.; Cardin, D.B.; Lenz, H.J.; Messersmith, W.; O’Neil, B.; Cohen, S.J.; Denlinger, C.S.; Shahda, S.; Astsaturov, I.; Kapoun, A.M.; et al. Phase Ib Study of Wnt Inhibitor Ipafricept with Gemcitabine and nab-paclitaxel in Patients with Previously Untreated Stage IV Pancreatic Cancer. Clin. Cancer Res. 2020, 26, 5348–5357. [Google Scholar] [CrossRef] [PubMed]
- Shetti, D.; Zhang, B.; Fan, C.; Mo, C.; Lee, B.H.; Wei, K. Low Dose of Paclitaxel Combined with XAV939 Attenuates Metastasis, Angiogenesis and Growth in Breast Cancer by Suppressing Wnt Signaling. Cells 2019, 8, 892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, W.; Chen, L.; Cai, X.; Zhang, Y.; Zhang, J.; Ma, D.; Cai, X.; Fu, T.; Yu, Z.; Yu, F.; et al. Long non-coding RNA TUC338 is functionally involved in sorafenib-sensitized hepatocarcinoma cells by targeting RASAL1. Oncol. Rep. 2017, 37, 273–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, S.; Tang, S. WNT/beta-catenin signaling in the development of liver cancers. Biomed. Pharmacother. 2020, 132, 110851. [Google Scholar] [CrossRef] [PubMed]
- Neiheisel, A.; Kaur, M.; Ma, N.; Havard, P.; Shenoy, A.K. Wnt pathway modulators in cancer therapeutics: An update on completed and ongoing clinical trials. Int. J. Cancer 2022, 150, 727–740. [Google Scholar] [CrossRef]
- Attia, R.T.; Tolba, M.F.; Trivedi, R.; Tadros, M.G.; Arafa, H.M.; Abdel-Naim, A.B. The chemomodulatory effects of glufosfamide on docetaxel cytotoxicity in prostate cancer cells. PeerJ 2016, 4, e2168. [Google Scholar] [CrossRef]
- Wang, H.; Word, B.R.; Lyn-Cook, B.D. Enhanced efficacy of gemcitabine by indole-3-carbinol in pancreatic cell lines: The role of human equilibrative nucleoside transporter 1. Anticancer Res. 2011, 31, 3171–3180. [Google Scholar]
- Jiang, J.; Lan, C.; Li, L.; Yang, D.; Xia, X.; Liao, Q.; Fu, W.; Chen, X.; An, S.; Wang, W.E.; et al. A novel porcupine inhibitor blocks WNT pathways and attenuates cardiac hypertrophy. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 3459–3467. [Google Scholar] [CrossRef]
- Schachter, J.; Ribas, A.; Long, G.V.; Arance, A.; Grob, J.J.; Mortier, L.; Daud, A.; Carlino, M.S.; McNeil, C.; Lotem, M.; et al. Pembrolizumab versus ipilimumab for advanced melanoma: Final overall survival results of a multicentre, randomised, open-label phase 3 study (KEYNOTE-006). Lancet 2017, 390, 1853–1862. [Google Scholar] [CrossRef]
- Madan, B.; Ke, Z.; Harmston, N.; Ho, S.Y.; Frois, A.O.; Alam, J.; Jeyaraj, D.A.; Pendharkar, V.; Ghosh, K.; Virshup, I.H.; et al. Wnt addiction of genetically defined cancers reversed by PORCN inhibition. Oncogene 2016, 35, 2197–2207. [Google Scholar] [CrossRef] [Green Version]
- Shah, K.; Panchal, S.; Patel, B. Porcupine inhibitors: Novel and emerging anti-cancer therapeutics targeting the Wnt signaling pathway. Pharmacol. Res. 2021, 167, 105532. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Pan, S.; Hsieh, M.H.; Ng, N.; Sun, F.; Wang, T.; Kasibhatla, S.; Schuller, A.G.; Li, A.G.; Cheng, D.; et al. Targeting Wnt-driven cancer through the inhibition of Porcupine by LGK974. Proc. Natl. Acad. Sci. USA 2013, 110, 20224–20229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.K.; Cho, Y.H.; Cha, P.H.; Yoon, J.S.; Ro, E.J.; Jeong, W.J.; Park, J.; Kim, H.; Il Kim, T.; Min, D.S.; et al. A small molecule approach to degrade RAS with EGFR repression is a potential therapy for KRAS mutation-driven colorectal Cancer Res.istance to cetuximab. Exp. Mol. Med. 2018, 50, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, B.Y.; Chiu, K.; Chung, H.; Bossard, C.; Nguyen, J.D.; Creger, E.; Eastman, B.W.; Mak, C.C.; Ibanez, M.; Ghias, A.; et al. The CLK inhibitor SM08502 induces anti-tumor activity and reduces Wnt pathway gene expression in gastrointestinal cancer models. Cancer Lett. 2020, 473, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, V.; Hu, H.; Barroga, C.; Bossard, C.; Kc, S.; Dellamary, L.; Stewart, J.; Chiu, K.; Ibanez, M.; Pedraza, M.; et al. A small-molecule inhibitor of the Wnt pathway (SM04690) as a potential disease modifying agent for the treatment of osteoarthritis of the knee. Osteoarthr. Cartil. 2018, 26, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, V.; Seo, T.; O’Green, A.L.; Ibanez, M.; Hofilena, B.; Kc, S.; Stewart, J.; Dellamary, L.; Chiu, K.; Ghias, A.; et al. SM04755, a small-molecule inhibitor of the Wnt pathway, as a potential topical treatment for tendinopathy. J Orthop Res. 2021, 39, 2048–2061. [Google Scholar] [CrossRef]






| Substrate Proteins | Description | References |
|---|---|---|
| β-catenin | Src phosphorylates Tyr333 of β-catenin and renders to bind with PKM2. | [100] |
| β-catenin Tyr654 phosphorylated by Src is released from E-cadherin. | [118] | |
| p-Tyr654 β-catenin translocates to nucleus. | [119] | |
| Src phosphorylates Tyr86 in β-catenin, leading to dissociation from E-cadherin. | [120] | |
| β-catenin phosphorylation at Tyr654 facilitates binding with p-Smad2 in TGF-β signaling. | [121] | |
| Dishevelled-2 | Src phosphorylates Tyr18, Tyr27, Tyr275 and Tyr463 of Dishevelled-2. Src binds to Dishevelled and Dishevelled disrupts Src autoinhibition, thereby Src can phosphorylate its substrate. | [114] |
| LRP6 | Src associates with LRP6 and phosphorylates Tyr1460, leading to feedback inhibition in Wnt signaling. | [117] |
| Frizzled receptor | Src associated with LRP5/6 receptor phosphorylates Fzd2 receptor at Tyr552. | [115] |
| ROR2 | Interaction of Src with ROR2 is critical for metastasis. | [140] |
| GSK-3β | Src phosphorylates Tyr216 of GSK-3β, an active form. | [141] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Min, J.K.; Park, H.-S.; Lee, Y.-B.; Kim, J.-G.; Kim, J.-I.; Park, J.-B. Cross-Talk between Wnt Signaling and Src Tyrosine Kinase. Biomedicines 2022, 10, 1112. https://doi.org/10.3390/biomedicines10051112
Min JK, Park H-S, Lee Y-B, Kim J-G, Kim J-I, Park J-B. Cross-Talk between Wnt Signaling and Src Tyrosine Kinase. Biomedicines. 2022; 10(5):1112. https://doi.org/10.3390/biomedicines10051112
Chicago/Turabian StyleMin, Jung Ki, Hwee-Seon Park, Yoon-Beom Lee, Jae-Gyu Kim, Jong-Il Kim, and Jae-Bong Park. 2022. "Cross-Talk between Wnt Signaling and Src Tyrosine Kinase" Biomedicines 10, no. 5: 1112. https://doi.org/10.3390/biomedicines10051112