
Plasma Metabolomics and Machine Learning-Driven Novel Diagnostic Signature for Non-Alcoholic Steatohepatitis

 , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. Clinical and Laboratory Evaluation
2.3. Imaging Biomarker Studies
2.4. Liver Tissue Sampling and Analyses
2.5. Metabolomics Analysis
2.6. Star Pattern Recognition Analysis
2.7. Identification of Potential Biomarkers and Related Metabolic Pathways
2.8. Machine Learning and Multinomial Logistic Regression
2.9. Multinomial Logistic Regression-Based Feature Selection
2.10. Decision Tree-Based Feature Selection
2.11. Formulation of metaNASH Score
2.12. Statistical Analysis
3. Results
3.1. Characteristics of Participants
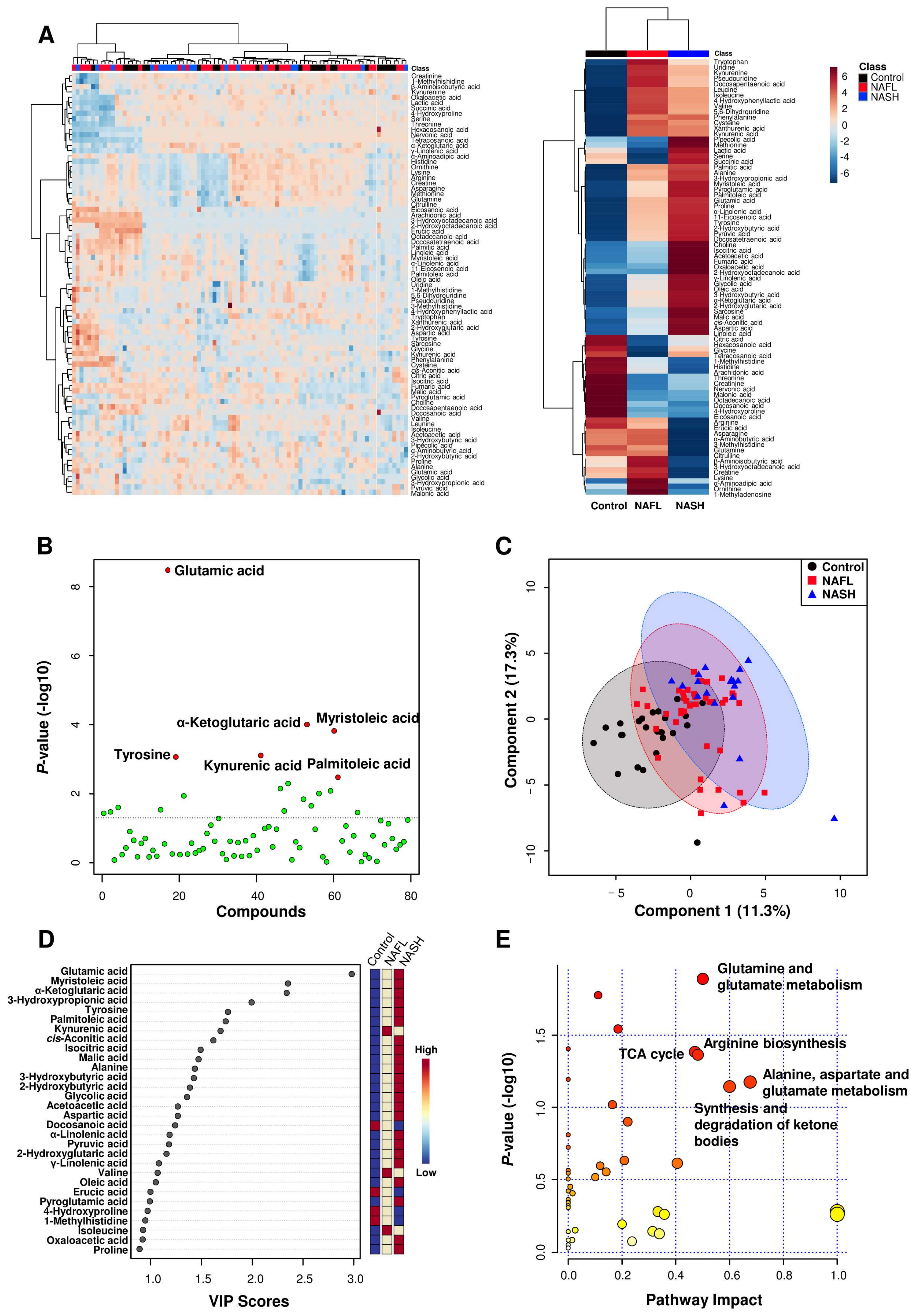
3.2. Results of Metabolomics Analyses
3.3. Plasma Metabolite Profiling and Univariate Analyses
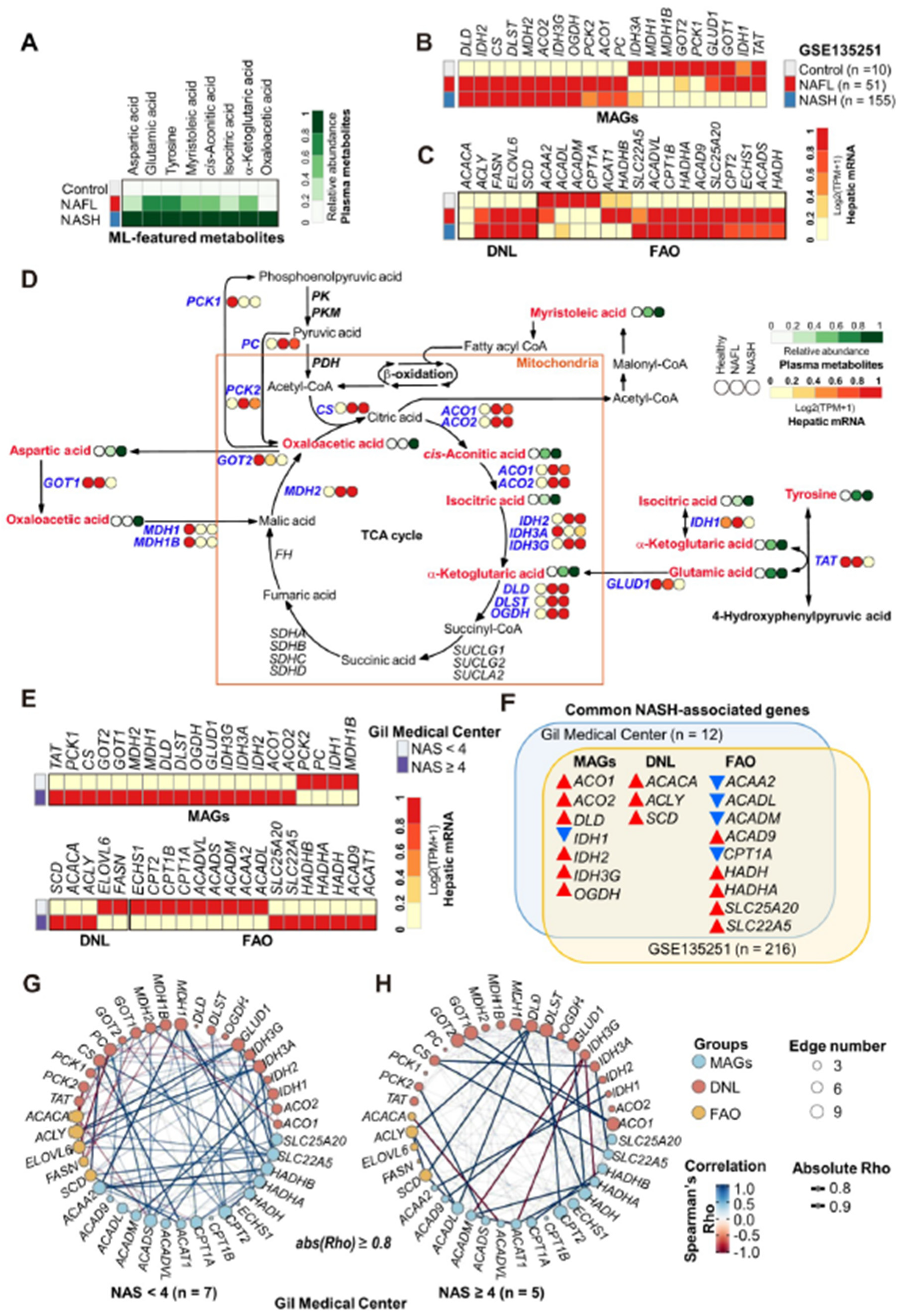
3.4. Identification of Potential Biomarkers and the Construction of Metabolic Pathways
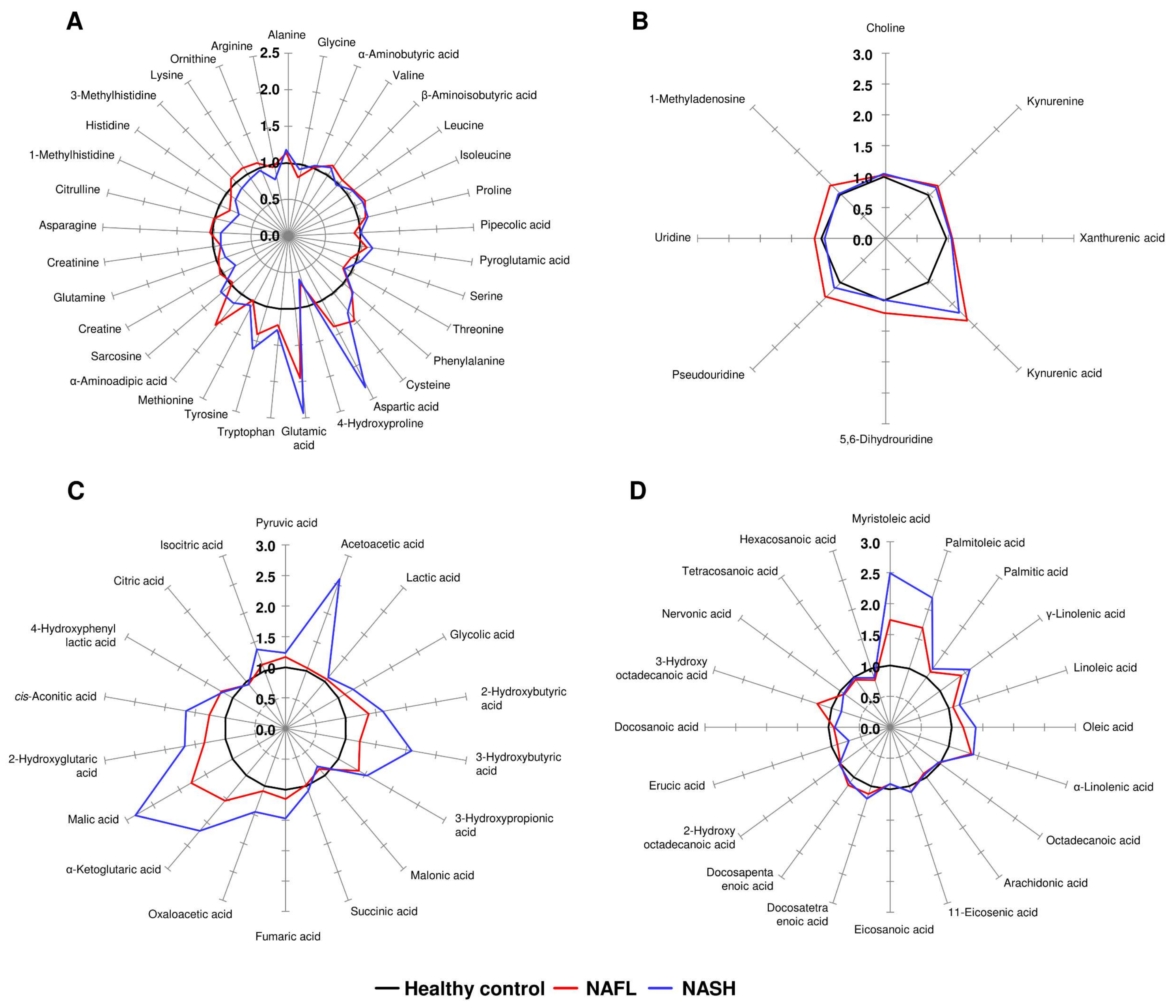
3.5. Star Pattern Recognition Analysis
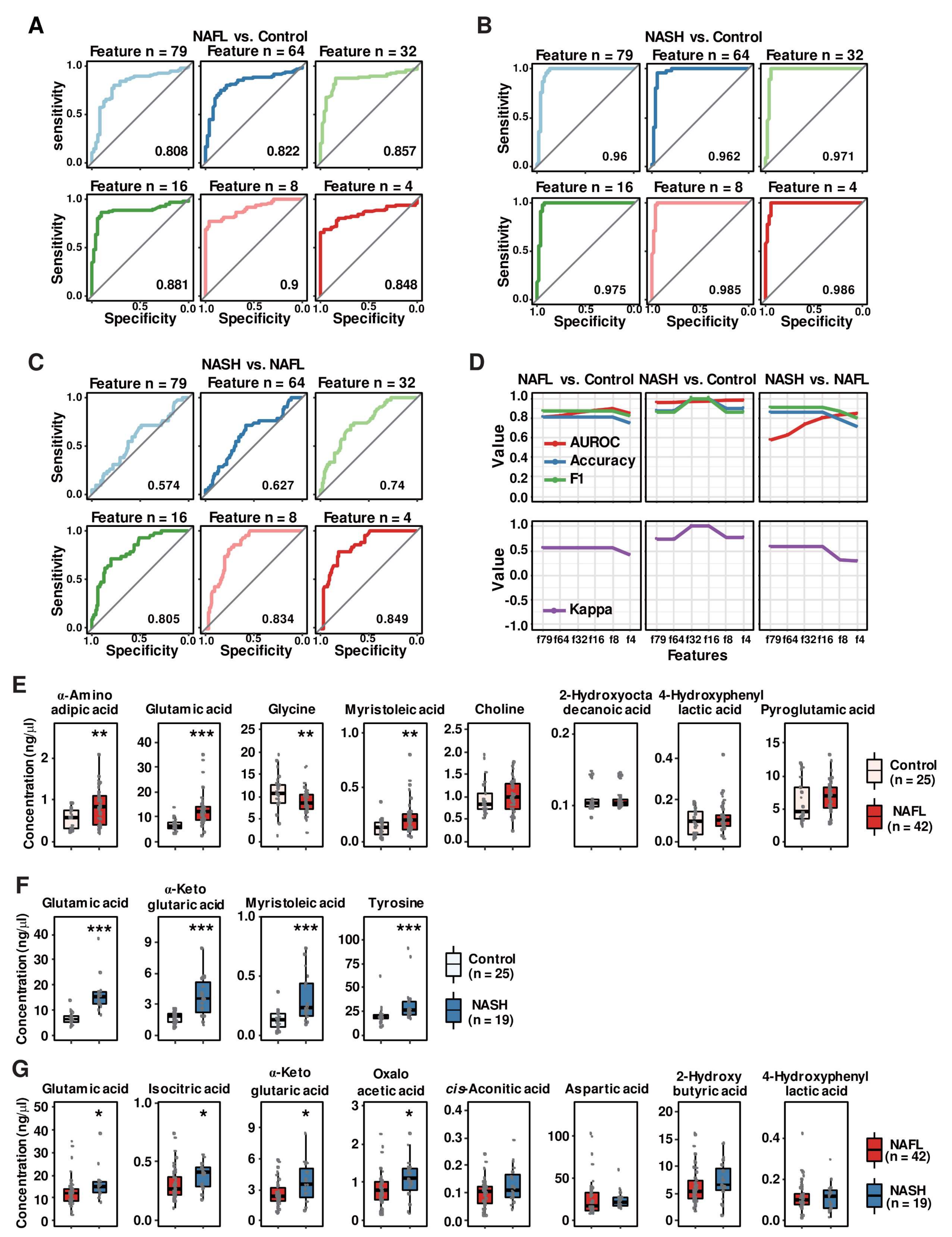
3.6. Random Forest Algorithm-Based Modeling Predicts Metabolites That Distinguish the Progression of NAFLD
3.7. Multinomial Logistic Regression Analysis Identified Eight Plasma Metabolites
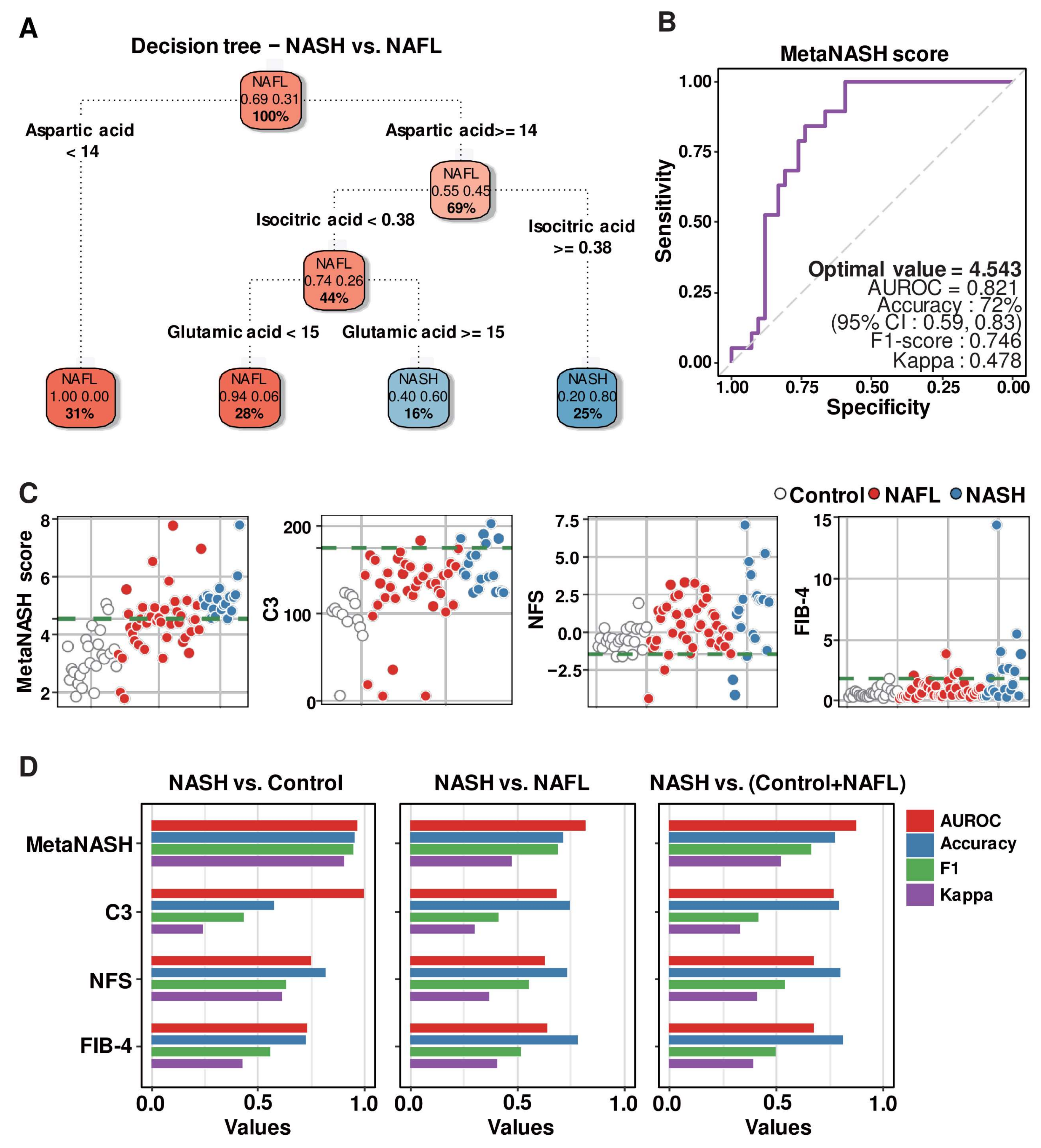
3.8. Decision Tree Algorithm Defined the Three Most Critical Plasma Metabolites, Distinguishing Patients with NAFL from Those with NASH
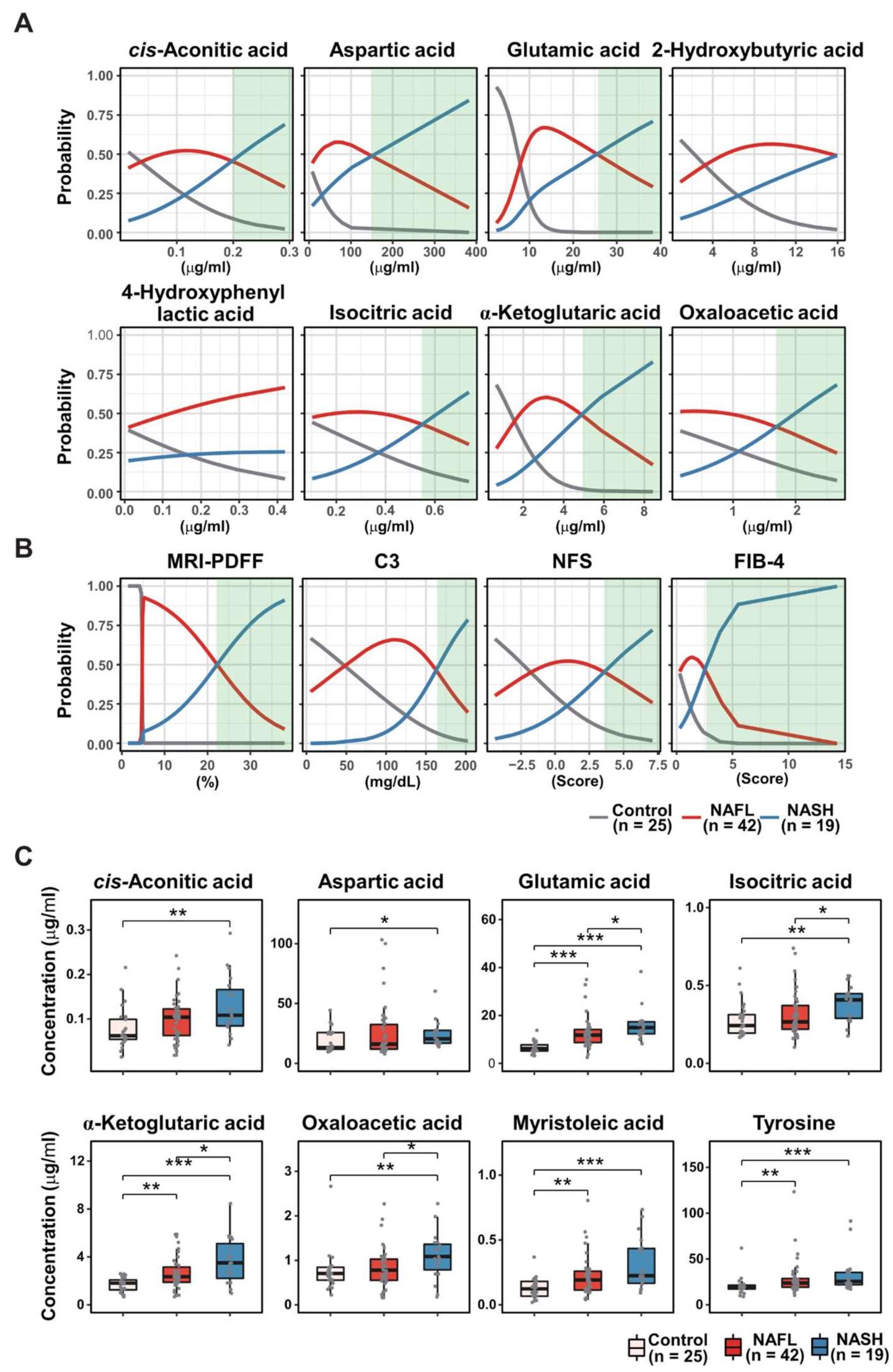
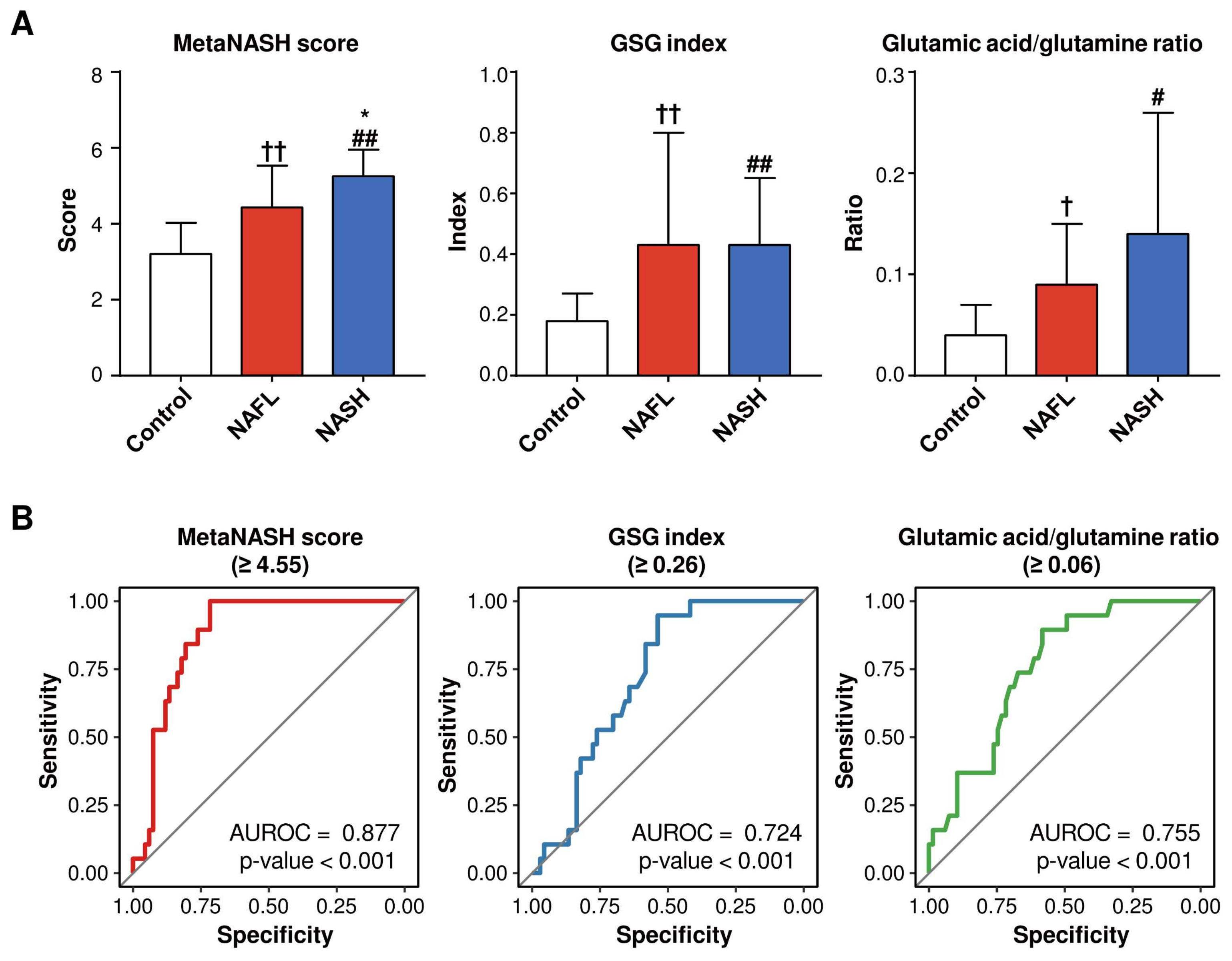
3.9. MetaNASH Score: A Metabolite-Based NASH Diagnostic Tool with Acceptable Performance
3.10. The MetaNASH Score Performed Better Than the GSG Index and Glutamic Acid/Glutamine Ratio in the Discrimination of NASH
3.11. Metabolic Remodeling in NASH Was Associated with Altered Gene Expression Profiles in the Liver
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marjot, T.; Moolla, A.; Cobbold, J.F.; Hodson, L.; Tomlinson, J.W. Nonalcoholic Fatty Liver Disease in Adults: Current Concepts in Etiology, Outcomes, and Management. Endocr. Rev. 2020, 41, 66–117. [Google Scholar] [CrossRef] [PubMed]
- Mato, J.M.; Alonso, C.; Noureddin, M.; Lu, S.C. Biomarkers and subtypes of deranged lipid metabolism in non-alcoholic fatty liver disease. World J. Gastroenterol. 2019, 25, 3009–3020. [Google Scholar] [CrossRef] [PubMed]
- Athyros, V.G.; Alexandrides, T.K.; Bilianou, H.; Cholongitas, E.; Doumas, M.; Ganotakis, E.S.; Goudevenos, J.; Elisaf, M.S.; Germanidis, G.; Giouleme, O.; et al. The use of statins alone, or in combination with pioglitazone and other drugs, for the treatment of non-alcoholic fatty liver disease/non-alcoholic steatohepatitis and related cardiovascular risk. An Expert Panel Statement. Metab. Clin. Exp. 2017, 71, 17–32. [Google Scholar] [CrossRef]
- Sheka, A.C.; Adeyi, O.; Thompson, J.; Hameed, B.; Crawford, P.A.; Ikramuddin, S. Nonalcoholic Steatohepatitis: A Review. JAMA 2020, 323, 1175–1183. [Google Scholar] [CrossRef]
- Lee, D.H. Noninvasive Evaluation of Nonalcoholic Fatty Liver Disease. Endocrinol. Metab. 2020, 35, 243–259. [Google Scholar] [CrossRef]
- Zhou, Y.; Orešič, M.; Leivonen, M.; Gopalacharyulu, P.; Hyysalo, J.; Arola, J.; Verrijken, A.; Francque, S.; Van Gaal, L.; Hyötyläinen, T.; et al. Noninvasive Detection of Nonalcoholic Steatohepatitis Using Clinical Markers and Circulating Levels of Lipids and Metabolites. Clin. Gastroenterol. Hepatol. Off. Clin. Pract. J. Am. Gastroenterol. Assoc. 2016, 14, 1463–1472.e1466. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.J.; Kim, S.M.; Kim, Y.S.; Kwon, O.S.; Shin, S.K.; Kim, K.K.; Lee, K.; Park, I.B.; Choi, C.S.; Chung, D.H.; et al. Magnetic Resonance-Based Assessments Better Capture Pathophysiologic Profiles and Progression in Nonalcoholic Fatty Liver Disease. Diabetes Metab. J. 2021, 45, 739–752. [Google Scholar] [CrossRef]
- Lee, B.W.; Lee, Y.H.; Park, C.Y.; Rhee, E.J.; Lee, W.Y.; Kim, N.H.; Choi, K.M.; Park, K.G.; Choi, Y.K.; Cha, B.S.; et al. Non-Alcoholic Fatty Liver Disease in Patients with Type 2 Diabetes Mellitus: A Position Statement of the Fatty Liver Research Group of the Korean Diabetes Association. Diabetes Metab. J. 2020, 44, 382–401. [Google Scholar] [CrossRef]
- Perakakis, N.; Stefanakis, K.; Mantzoros, C.S. The role of omics in the pathophysiology, diagnosis and treatment of non-alcoholic fatty liver disease. Metab. Clin. Exp. 2020, 111s, 154320. [Google Scholar] [CrossRef]
- Pirola, C.J.; Sookoian, S. Multiomics biomarkers for the prediction of nonalcoholic fatty liver disease severity. World J. Gastroenterol. 2018, 24, 1601–1615. [Google Scholar] [CrossRef] [PubMed]
- Newgard, C.B. Metabolomics and Metabolic Diseases: Where Do We Stand? Cell Metab. 2017, 25, 43–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puri, P.; Wiest, M.M.; Cheung, O.; Mirshahi, F.; Sargeant, C.; Min, H.K.; Contos, M.J.; Sterling, R.K.; Fuchs, M.; Zhou, H.; et al. The plasma lipidomic signature of nonalcoholic steatohepatitis. Hepatology 2009, 50, 1827–1838. [Google Scholar] [CrossRef] [Green Version]
- Puri, P.; Daita, K.; Joyce, A.; Mirshahi, F.; Santhekadur, P.K.; Cazanave, S.; Luketic, V.A.; Siddiqui, M.S.; Boyett, S.; Min, H.K.; et al. The presence and severity of nonalcoholic steatohepatitis is associated with specific changes in circulating bile acids. Hepatology 2018, 67, 534–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalhan, S.C.; Guo, L.; Edmison, J.; Dasarathy, S.; McCullough, A.J.; Hanson, R.W.; Milburn, M. Plasma metabolomic profile in nonalcoholic fatty liver disease. Metab. Clin. Exp. 2011, 60, 404–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mardinoglu, A.; Agren, R.; Kampf, C.; Asplund, A.; Uhlen, M.; Nielsen, J. Genome-scale metabolic modelling of hepatocytes reveals serine deficiency in patients with non-alcoholic fatty liver disease. Nat. Commun. 2014, 5, 3083. [Google Scholar] [CrossRef] [Green Version]
- Sookoian, S.; Pirola, C.J. NAFLD. Metabolic make-up of NASH: From fat and sugar to amino acids. Nat. Reviews. Gastroenterol. Hepatol. 2014, 11, 205–207. [Google Scholar] [CrossRef]
- Sookoian, S.; Castaño, G.O.; Scian, R.; Fernández Gianotti, T.; Dopazo, H.; Rohr, C.; Gaj, G.; San Martino, J.; Sevic, I.; Flichman, D.; et al. Serum aminotransferases in nonalcoholic fatty liver disease are a signature of liver metabolic perturbations at the amino acid and Krebs cycle level. Am. J. Clin. Nutr. 2016, 103, 422–434. [Google Scholar] [CrossRef] [Green Version]
- Sookoian, S.; Puri, P.; Castaño, G.O.; Scian, R.; Mirshahi, F.; Sanyal, A.J.; Pirola, C.J. Nonalcoholic steatohepatitis is associated with a state of betaine-insufficiency. Liver Int. Off. J. Int. Assoc. Study Liver 2017, 37, 611–619. [Google Scholar] [CrossRef]
- Alonso, C.; Fernández-Ramos, D.; Varela-Rey, M.; Martínez-Arranz, I.; Navasa, N.; Van Liempd, S.M.; Lavín Trueba, J.L.; Mayo, R.; Ilisso, C.P.; de Juan, V.G.; et al. Metabolomic Identification of Subtypes of Nonalcoholic Steatohepatitis. Gastroenterology 2017, 152, 1449–1461.e1447. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Gallego, E.; Guirro, M.; Riera-Borrull, M.; Hernández-Aguilera, A.; Mariné-Casadó, R.; Fernández-Arroyo, S.; Beltrán-Debón, R.; Sabench, F.; Hernández, M.; del Castillo, D.; et al. Mapping of the circulating metabolome reveals α-ketoglutarate as a predictor of morbid obesity-associated non-alcoholic fatty liver disease. Int. J. Obes. 2015, 39, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Liebal, U.W.; Phan, A.N.T.; Sudhakar, M.; Raman, K.; Blank, L.M. Machine Learning Applications for Mass Spectrometry-Based Metabolomics. Metabolites 2020, 10, 243. [Google Scholar] [CrossRef] [PubMed]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Seo, C.; Kim, S.H.; Lee, H.S.; Ji, M.; Min, J.; Son, Y.J.; Kim, I.H.; Lee, K.; Paik, M.J. Metabolomic study on bleomycin and polyhexamethylene guanidine phosphate-induced pulmonary fibrosis mice models. Metabolomics 2019, 15, 111. [Google Scholar] [CrossRef]
- Choi, R.Y.; Ji, M.; Lee, M.K.; Paik, M.J. Metabolomics Study of Serum from a Chronic Alcohol-Fed Rat Model Following Administration of Defatted Tenebrio molitor Larva Fermentation Extract. Metabolites 2020, 10, 436. [Google Scholar] [CrossRef]
- Seo, C.; Hwang, Y.H.; Lee, H.S.; Kim, Y.; Shin, T.H.; Lee, G.; Son, Y.J.; Kim, H.; Yee, S.T.; Park, A.K.; et al. Metabolomic study for monitoring of biomarkers in mouse plasma with asthma by gas chromatography-mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2017, 1063, 156–162. [Google Scholar] [CrossRef]
- Lee, H.S.; Seo, C.; Hwang, Y.H.; Shin, T.H.; Park, H.J.; Kim, Y.; Ji, M.; Min, J.; Choi, S.; Kim, H.; et al. Metabolomic approaches to polyamines including acetylated derivatives in lung tissue of mice with asthma. Metabolomics 2019, 15, 8. [Google Scholar] [CrossRef]
- Caret: Classification and Regression Training. Available online: https://github.com/topepo/caret/ (accessed on 19 April 2022).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002; ISBN 0-387-95457-0. [Google Scholar]
- Bae, J.H.; Jeong, H.J.; Kim, H.; Leem, Y.E.; Ryu, D.; Park, S.C.; Lee, Y.I.; Cho, S.C.; Kang, J.S. ZNF746/PARIS overexpression induces cellular senescence through FoxO1/p21 axis activation in myoblasts. Cell Death Dis. 2020, 11, 359. [Google Scholar] [CrossRef]
- Kim, H.; Cho, S.C.; Jeong, H.J.; Lee, H.Y.; Jeong, M.H.; Pyun, J.H.; Ryu, D.; Kim, M.; Lee, Y.S.; Kim, M.S.; et al. Indoprofen prevents muscle wasting in aged mice through activation of PDK1/AKT pathway. J. Cachexia Sarcopenia Muscle 2020, 11, 1070–1088. [Google Scholar] [CrossRef]
- Geurts, P.; Irrthum, A.; Wehenkel, L. Supervised learning with decision tree-based methods in computational and systems biology. Mol. Biosyst. 2009, 5, 1593–1605. [Google Scholar] [CrossRef]
- Rensen, S.S.; Slaats, Y.; Driessen, A.; Peutz-Kootstra, C.J.; Nijhuis, J.; Steffensen, R.; Greve, J.W.; Buurman, W.A. Activation of the complement system in human nonalcoholic fatty liver disease. Hepatology 2009, 50, 1809–1817. [Google Scholar] [CrossRef] [PubMed]
- Youden, W.J. Index for rating diagnostic tests. Cancer 1950, 3, 32–35. [Google Scholar] [CrossRef]
- Gaggini, M.; Carli, F.; Rosso, C.; Buzzigoli, E.; Marietti, M.; Della Latta, V.; Ciociaro, D.; Abate, M.L.; Gambino, R.; Cassader, M.; et al. Altered amino acid concentrations in NAFLD: Impact of obesity and insulin resistance. Hepatology 2018, 67, 145–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, K.; Chitneni, S.K.; Suzuki, A.; Wang, Y.; Henao, R.; Hyun, J.; Premont, R.T.; Naggie, S.; Moylan, C.A.; Bashir, M.R.; et al. Increased Glutaminolysis Marks Active Scarring in Nonalcoholic Steatohepatitis Progression. Cell Mol. Gastroenterol. Hepatol. 2020, 10, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Govaere, O.; Cockell, S.; Tiniakos, D.; Queen, R.; Younes, R.; Vacca, M.; Alexander, L.; Ravaioli, F.; Palmer, J.; Petta, S.; et al. Transcriptomic profiling across the nonalcoholic fatty liver disease spectrum reveals gene signatures for steatohepatitis and fibrosis. Sci. Transl. Med. 2020, 12, eaba4448. [Google Scholar] [CrossRef]
- Portillo-Sanchez, P.; Bril, F.; Maximos, M.; Lomonaco, R.; Biernacki, D.; Orsak, B.; Subbarayan, S.; Webb, A.; Hecht, J.; Cusi, K. High Prevalence of Nonalcoholic Fatty Liver Disease in Patients With Type 2 Diabetes Mellitus and Normal Plasma Aminotransferase Levels. J. Clin. Endocrinol. Metab. 2015, 100, 2231–2238. [Google Scholar] [CrossRef]
- Hyötyläinen, T.; Jerby, L.; Petäjä, E.M.; Mattila, I.; Jäntti, S.; Auvinen, P.; Gastaldelli, A.; Yki-Järvinen, H.; Ruppin, E.; Orešič, M. Genome-scale study reveals reduced metabolic adaptability in patients with non-alcoholic fatty liver disease. Nat. Commun. 2016, 7, 8994. [Google Scholar] [CrossRef]
- Bianchi, G.; Marchesini, G.; Brunetti, N.; Manicardi, E.; Montuschi, F.; Chianese, R.; Zoli, M. Impaired insulin-mediated amino acid plasma disappearance in non-alcoholic fatty liver disease: A feature of insulin resistance. Dig. Liver Dis. 2003, 35, 722–727. [Google Scholar] [CrossRef]
- Tufi, R.; Gandhi, S.; de Castro, I.P.; Lehmann, S.; Angelova, P.R.; Dinsdale, D.; Deas, E.; Plun-Favreau, H.; Nicotera, P.; Abramov, A.Y.; et al. Enhancing nucleotide metabolism protects against mitochondrial dysfunction and neurodegeneration in a PINK1 model of Parkinson’s disease. Nat. Cell Biol. 2014, 16, 157–166. [Google Scholar] [CrossRef] [Green Version]
- Brosnan, M.E.; Brosnan, J.T. Hepatic glutamate metabolism: A tale of 2 hepatocytes. Am. J. Clin. Nutr. 2009, 90, 857s–861s. [Google Scholar] [CrossRef] [Green Version]
- Walker, V. Ammonia metabolism and hyperammonemic disorders. Adv. Clin. Chem. 2014, 67, 73–150. [Google Scholar] [CrossRef]
- Yang, L.; Venneti, S.; Nagrath, D. Glutaminolysis: A Hallmark of Cancer Metabolism. Annu. Rev. Biomed. Eng. 2017, 19, 163–194. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.M.; Kim, H.H.; Kim, M.H.; Cinar, R.; Yi, H.S.; Eun, H.S.; Kim, S.H.; Choi, Y.J.; Lee, Y.S.; Kim, S.Y.; et al. Glutamate Signaling in Hepatic Stellate Cells Drives Alcoholic Steatosis. Cell Metab. 2019, 30, 877–889.e877. [Google Scholar] [CrossRef] [PubMed]
- Mallat, A.; Lotersztajn, S. Glutamate Signaling in Alcohol-associated Fatty Liver: “Pas de Deux”. Hepatology 2020, 72, 350–352. [Google Scholar] [CrossRef] [PubMed]
- Iacobazzi, V.; Infantino, V. Citrate—New functions for an old metabolite. Biol. Chem. 2014, 395, 387–399. [Google Scholar] [CrossRef]
- Van de Wier, B.; Balk, J.M.; Haenen, G.R.; Giamouridis, D.; Bakker, J.A.; Bast, B.C.; den Hartog, G.J.; Koek, G.H.; Bast, A. Elevated citrate levels in non-alcoholic fatty liver disease: The potential of citrate to promote radical production. FEBS Lett. 2013, 587, 2461–2466. [Google Scholar] [CrossRef] [Green Version]
- Tricò, D.; Biancalana, E.; Solini, A. Protein and amino acids in nonalcoholic fatty liver disease. Curr. Opin Clin. Nutr. Metab. Care 2021, 24, 96–101. [Google Scholar] [CrossRef]
- Lake, A.D.; Novak, P.; Shipkova, P.; Aranibar, N.; Robertson, D.G.; Reily, M.D.; Lehman-McKeeman, L.D.; Vaillancourt, R.R.; Cherrington, N.J. Branched chain amino acid metabolism profiles in progressive human nonalcoholic fatty liver disease. Amino Acids 2015, 47, 603–615. [Google Scholar] [CrossRef]
- Kakazu, E.; Sano, A.; Morosawa, T.; Inoue, J.; Ninomiya, M.; Iwata, T.; Nakamura, T.; Takai, S.; Sawada, S.; Katagiri, H.; et al. Branched chain amino acids are associated with the heterogeneity of the area of lipid droplets in hepatocytes of patients with non-alcoholic fatty liver disease. Hepatol. Res. Off. J. Jpn. Soc. Hepatol. 2019, 49, 860–871. [Google Scholar] [CrossRef]
- Seo, C.; Na, M.; Jang, J.; Park, M.; Choi, B.; Lee, S.; Paik, M.J. Monitoring of altered amino acid metabolic pattern in rat urine following intraperitoneal injection with γ-hydroxybutyric acid. Metabolomics 2018, 14, 111. [Google Scholar] [CrossRef]
- Shin, T.H.; Seo, C.; Lee, D.Y.; Ji, M.; Manavalan, B.; Basith, S.; Chakkarapani, S.K.; Kang, S.H.; Lee, G.; Paik, M.J.; et al. Silica-coated magnetic nanoparticles induce glucose metabolic dysfunction in vitro via the generation of reactive oxygen species. Arch. Toxicol. 2019, 93, 1201–1212. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Control (n = 25) | NAFL (n = 42) | NASH (n = 19) | p-Values |
|---|---|---|---|---|
| Age (years) | 35.2 (15.3) | 43.2 (15.7) † | 41 (16.2) | 0.11 |
| Sex (male/female) | 15/10 | 19/23 | 12/7 | NA |
| Weight (kg) | 65.9 (10.2) | 84.4 (18.5) ††† | 104.3 (29.4) ###, ** | <0.001 |
| BMI (kg/m2) | 23.2 (2.9) | 30.6 (5.9) ††† | 35.5 (7) ###, ** | <0.001 |
| Waist circumference (cm) | 79.9 (6.8) | 100.6 (14.7) ††† | 113.5 (15.8) ###, ** | <0.001 |
| SBP (mmHg) | 128.7 (18.3) | 128.6 (14.4) | 130.2 (14.1) | 0.93 |
| DBP (mmHg) | 82.8 (12) | 85.2 (9.4) | 87.2 (12.3) | 0.3 |
| AST (U/L) | 20.4 (5.5) | 42.8 (45.1) ††† | 87.1 (58.1) ###, *** | <0.001 |
| ALT (U/L) | 18.4 (7.4) | 62.3 (78.3) ††† | 118.8 (100.3) ###, * | <0.001 |
| GGT (U/L) | 18.3 (8.2) | 45.7 (32.1) ††† | 96.7 (58.8) ###, *** | <0.001 |
| Total cholesterol (mg/dL) | 189.3 (33.9) | 194.5 (35.6) | 180.9 (38.6) | 0.4 |
| HDL-C (mg/dL) | 59.3 (14.2) | 50.5 (11.2) †† | 49.4 (27.2) ## | <0.05 |
| Triglycerides (mg/dL) | 97.1 (46.6) | 156.6 (110.1) †† | 190.2 (110.9) ### | <0.001 |
| White blood cell (×109/L) | 5.4 (1.6) | 6.6 (2) †† | 7.6 (2.2) ## | <0.05 |
| Platelet (×109/L) | 244.1 (59.1) | 267.6 (73.2) | 242.2 (94.2) | 0.28 |
| Hemoglobin A1c (%) | 8.7 (16.5) | 6.4 (1.8) †† | 7.6 (1.9) ###, ** | <0.001 |
| Glucose (mg/dL) | 86.5 (18.3) | 108.3 (31.9) †† | 129.8 (57) ###, * | <0.001 |
| Insulin (μU/mL) | 6.8 (3.8) | 17.6 (16.1) ††† | 27.9 (17.6) ###, ** | <0.001 |
| HOMA-IR | 1.6 (1) | 4.2 (3.9) ††† | 7.9 (5.2) ###, ** | <0.001 |
| C3 (mg/dL) | 94.9 (30.5) | 125.9 (43.4) ††† | 154.3 (26.1) ###, * | <0.001 |
| C4 (mg/dL) | 24.7 (5.5) | 28 (8.2) | 29.5 (11.4) | 0.24 |
| ELF score | 8.2 (0.8) | 8.8 (0.9) † | 9.7 (0.8) ###, *** | <0.001 |
| Liver MRI-PDFF (%) | 3.4 (0.8) | 12.6 (6.6) ††† | 23.2 (10) ###, *** | <0.001 |
| MRE-LSM (kPa) | 3.1 (0.6) | 3.4 (0.7) | 5.2 (1) ###, *** | <0.001 |
| NFS | −0.4 (0.8) | 0.4 (1.6) † | 1.4 (2.8) ## | <0.05 |
| FIB-4 | 0.7 (0.4) | 1.0 (0.7) | 2.4 (3.2) ## | <0.05 |
| Plasma Metabolites (ng/μL) | Control (n = 25) | NAFL (n = 42) | NASH (n = 19) | Normalized Values a | Kruskal-Wallis Test | ||
|---|---|---|---|---|---|---|---|
| NAFL | NASH | p Values | Q Values b | ||||
| Glutamic acid | 6.52 ± 2.47 | 12.74 ± 6.45 ††† | 15.88 ± 6.36 ###, * | 1.95 | 2.43 | <0.001 | <0.001 |
| Tyrosine | 20.15 ± 9.59 | 28.21 ± 18.65 †† | 32.41 ± 19.79 ### | 1.40 | 1.61 | 0.001 | 0.015 |
| Kynurenic acid | 0.005 ± 0.006 | 0.010 ± 0.009 ††† | 0.009 ± 0.006 ## | 1.88 | 1.70 | 0.001 | 0.015 |
| α-Ketoglutaric acid | 1.70 ± 0.56 | 2.63 ± 1.32 †† | 3.71 ± 1.85 ###, * | 1.54 | 2.18 | <0.001 | 0.004 |
| Myristoleic acid (C14:1) | 0.13 ± 0.08 | 0.22 ± 0.15 †† | 0.31 ± 0.19 ### | 1.74 | 2.50 | <0.001 | 0.004 |
| Palmitoleic acid (C16:1) | 3.26 ± 1.87 | 5.53 ± 3.97 †† | 7.18 ± 5.16 ## | 1.69 | 2.20 | 0.004 | 0.048 |
| Group | Mean ± SD | Mean of MetaNASH Score | Accuracy | F1 Score | |
|---|---|---|---|---|---|
| BMI | NAFL (n = 23) | 31.73 ± 2.64 | 4.326 | 0.6765 | 0.6857 |
| NASH (n = 11) | 32.26 ± 2.54 | 5.275 | |||
| Insulin | NAFL (n = 19) | 14.52 ± 2.15 | 4.328 | 0.7083 | 0.7742 |
| NASH (n = 5) | 13.87 ± 2.21 | 5.799 | |||
| Glucose | NAFL (n = 33) | 95.46 ±8.45 | 4.305 | 0.7273 | 0.7778 |
| NASH (n = 11) | 101.0 ± 9.91 | 5.150 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, M.; Jo, Y.; Choi, S.J.; Kim, S.M.; Kim, K.K.; Oh, B.-C.; Ryu, D.; Paik, M.-J.; Lee, D.H. Plasma Metabolomics and Machine Learning-Driven Novel Diagnostic Signature for Non-Alcoholic Steatohepatitis. Biomedicines 2022, 10, 1669. https://doi.org/10.3390/biomedicines10071669
Ji M, Jo Y, Choi SJ, Kim SM, Kim KK, Oh B-C, Ryu D, Paik M-J, Lee DH. Plasma Metabolomics and Machine Learning-Driven Novel Diagnostic Signature for Non-Alcoholic Steatohepatitis. Biomedicines. 2022; 10(7):1669. https://doi.org/10.3390/biomedicines10071669
Chicago/Turabian StyleJi, Moongi, Yunju Jo, Seung Joon Choi, Seong Min Kim, Kyoung Kon Kim, Byung-Chul Oh, Dongryeol Ryu, Man-Jeong Paik, and Dae Ho Lee. 2022. "Plasma Metabolomics and Machine Learning-Driven Novel Diagnostic Signature for Non-Alcoholic Steatohepatitis" Biomedicines 10, no. 7: 1669. https://doi.org/10.3390/biomedicines10071669
APA StyleJi, M., Jo, Y., Choi, S. J., Kim, S. M., Kim, K. K., Oh, B.-C., Ryu, D., Paik, M.-J., & Lee, D. H. (2022). Plasma Metabolomics and Machine Learning-Driven Novel Diagnostic Signature for Non-Alcoholic Steatohepatitis. Biomedicines, 10(7), 1669. https://doi.org/10.3390/biomedicines10071669