
Multiple Sclerosis: Enzymatic Cross Site-Specific Recognition and Hydrolysis of H2A Histone by IgGs against H2A, H1, H2B, H3 Histones, Myelin Basic Protein, and DNA


Abstract
:1. Introduction
2. Material and Methods
2.1. Chemicals, Donors, and Patients
2.2. Antibody Purification
2.3. Affinity Chromatography of IgGs on MBP-Sepharose
2.4. Affinity Chromatography of IgGs on DNA-Cellulose
2.5. Proteolytic Activity Assay
2.6. DNA Hydrolysis
2.7. MALDI-TOF Analysis of Abs-Dependent H2A Histone Hydrolysis
2.8. Analysis of Sequence Homology
2.9. Statistical Analysis
3. Results
3.1. Purification of Antibodies
3.2. Isolation of IgGs against MBP
3.3. Isolation of IgGs against Individual Histones
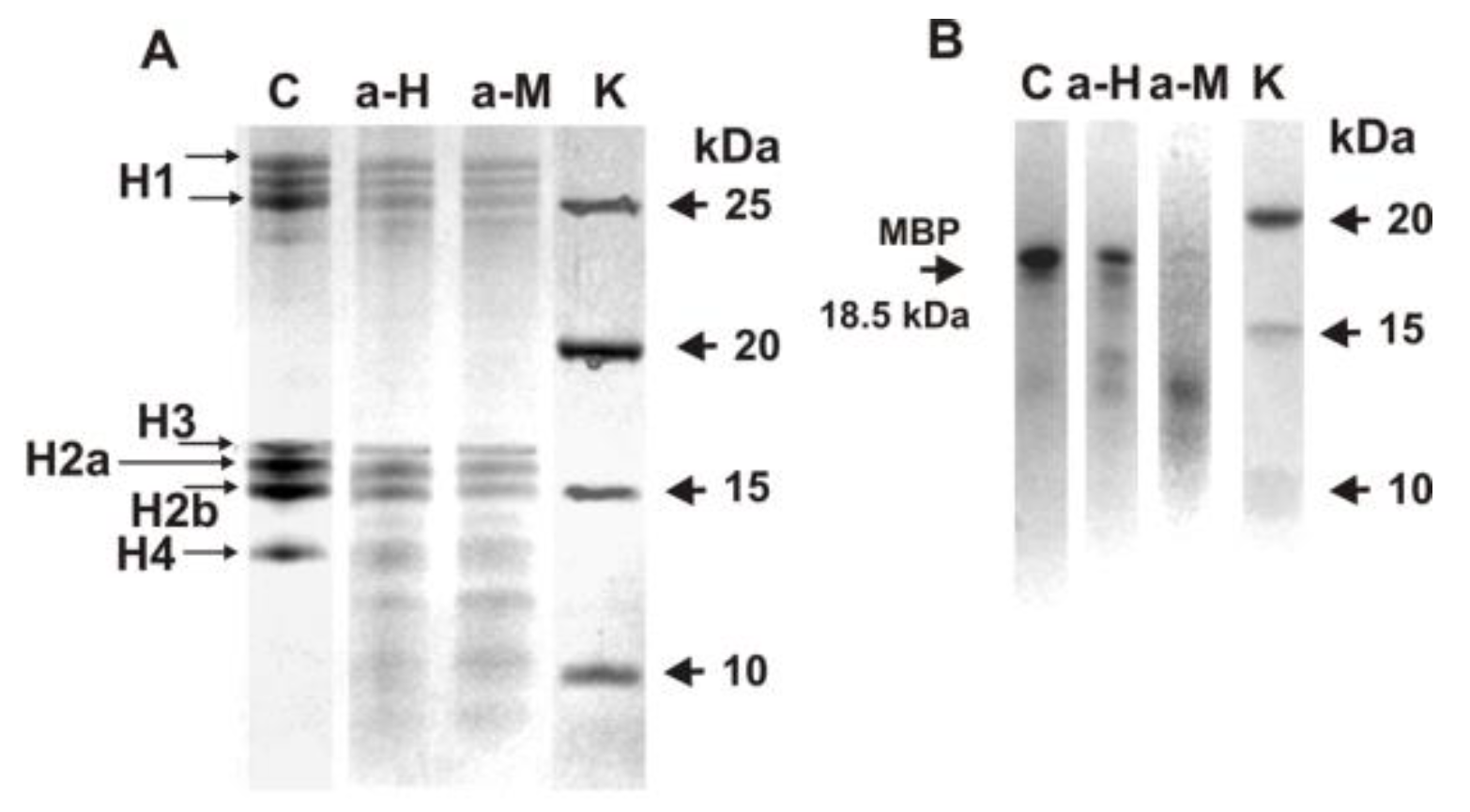
3.4. SDS-PAGE Analysis of Histones and MBP Hydrolysis
3.5. DNase Activity of IgGs against DNA and Proteins
3.6. MALDI Analysis of H2A Histone Hydrolysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Ab | antibody |
| AA | amino acid |
| ABZs | abzymes |
| ADs | autoimmune diseases |
| HIV-1 | human immunodeficiency virus type 1 |
| MALDI-TOF | laser matrix-assisted desorption/ionization time-of-flight mass spectrometry |
| MBP | myelin basic protein |
| SDS-PAGE | sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| MS | multiple sclerosis |
| SLE | systemic lupus erythematosus |
References
- David, B.S. Catalytic Antibodies; Keinan, E., Ed.; Wiley-VCH Verlag GmbH and Co. KgaA: Weinheim, Germany, 2005; pp. 1–586. [Google Scholar]
- Nevinsky, G.A.; Buneva, V.N. Natural catalytic antibodies–abzymes. In Catalytic Antibodies; Keinan, E., Ed.; VCH-Wiley Press: Weinheim, Germany, 2005; pp. 505–569. [Google Scholar]
- Nevinsky, G.A. Natural catalytic antibodies in norm and in autoimmune diseases, In Autoimmune Diseases: Symptoms, Diagnosis and Treatment; Brenner, K.J., Ed.; Nova Science Publishers Inc.: New York, NY, USA, 2010; pp. 1–107. [Google Scholar]
- Nevinsky, G.A. Natural catalytic antibodies in norm and in HIV-infected patients. In Understanding HIV/AIDS Management and Care—Pandemic Approaches the 21st Century; Kasenga, F.H., Ed.; InTech: Rijeka, Croatia, 2011; pp. 151–192. [Google Scholar]
- Nevinsky, G.A. Autoimmune processes in multiple sclerosis: Production of harmful catalytic antibodies associated with significant changes in the hematopoietic stem cell differentiation and proliferation. In Multiple Sclerosis; Conzalez-Quevedo, A., Ed.; InTech: Rijeka, Croatia, 2016; pp. 100–147. [Google Scholar]
- Nevinsky, G.A. Catalytic antibodies in norm and systemic lupus erythematosus. In Lupus; Khan, W.A., Ed.; InTech: Rijeka, Croatia, 2017; pp. 41–101. [Google Scholar]
- Jerne, N.K. Towards a network theory of the immune system. Ann. Immunol. 1974, 125, 373–398. [Google Scholar]
- Shuster, A.M.; Gololobov, G.V.; Kvashuk, O.A.; Bogomolova, A.E.; Smirnov, I.V.; Gabibov, A.G. DNA hydrolyzing autoantibodies. Science 1992, 256, 665–667. [Google Scholar] [CrossRef] [PubMed]
- Andrievskaya, O.A.; Buneva, V.N.; Naumov, V.A.; Nevinsky, G.A. Catalytic heterogenity of polyclonal RNA-hydrolyzing IgM from sera of patients with lupus erythematosus. Med. Sci. Monit. 2000, 6, 460–470. [Google Scholar] [PubMed]
- Andrievskaya, O.A.; Buneva, V.N.; Baranovskii, A.G.; Gal’vita, A.V.; Benzo, E.S.; Naumov, V.A.; Nevinsky, G.A. Catalytic diversity of polyclonal RNA-hydrolyzing IgG antibodies from the sera of patients with systemic lupus erythematosus. Immunol. Lett. 2002, 81, 191–198. [Google Scholar] [CrossRef]
- Baranovskii, A.G.; Kanyshkova, T.G.; Mogelnitskii, A.S.; Naumov, V.A.; Buneva, V.N.; Gusev, E.I.; Boiko, A.N.; Zargarova, T.A.; Favorova, O.O.; Nevinsky, G.A. Polyclonal antibodies from blood and cerebrospinal fluid of patients with multiple sclerosis effectively hydrolyze DNA and RNA. Biochemistry 1998, 63, 1239–1248. [Google Scholar]
- Baranovsky, A.G.; Matushin, V.G.; Vlassov, A.V.; Zabara, V.G.; Naumov, V.A.; Giege, R.; Buneva, V.N.; Nevinsky, G.A. DNA- and RNA-hydrolyzing antibodies from the blood of patients with various forms of viral hepatitis. Biochemistry 1997, 62, 1358–1366. [Google Scholar]
- Savel’ev, A.N.; Eneyskaya, E.V.; Shabalin, K.A.; Filatov, M.V.; Neustroev, K.N. Antibodies with amylolytic activity. Protein Peptide Lett. 1999, 6, 179–181. [Google Scholar]
- Savel’ev, A.N.; Kanyshkova, T.G.; Kulminskaya, A.A.; Buneva, V.N.; Eneyskaya, E.V.; Filatov, M.V.; Nevinsky, G.A.; Neustroev, K.N. Amylolytic activity of IgG and sIgA immunoglobulins from human milk. Clin. Chim. Acta 2001, 314, 141–152. [Google Scholar] [CrossRef]
- Neustoev, K.N.; Ivanen, D.R.; Kulminskaya, A.A.; Brumer, I.H.; Saveliev, A.N.; Nevinsky, G.A. Amylolytic activity and catalytic properties of IgM and IgG antibodies from patients with systemic lupus erythematosus. Hum. Antibodies 2003, 12, 31–34. [Google Scholar]
- Paul, S.; Volle, D.J.; Beach, C.M.; Johnson, D.R.; Powell, M.J.; Massey, R.J. Catalytic hydrolysis of vasoactive intestinal peptide by human autoantibody. Science 1989, 244, 1158–1162. [Google Scholar] [CrossRef]
- Li, L.; Paul, S.; Tyutyulkova, S.; Kazatchkine, M.D.; Kaveri, S. Catalytic activity of anti-thyroglobulin antibodies. J. Immunol. 1995, 154, 3328–3332. [Google Scholar]
- Kalaga, R.; Li, L.; O’Dell, J.R.; Paul, S. Unexpected presence of polyreactive catalytic antibodies in IgG from unimmunized donors and decreased levels in rheumatoid arthritis. J. Immunol. 1995, 155, 2695–2702. [Google Scholar]
- Thiagarajan, P.; Dannenbring, R.; Matsuura, K.; Tramontane, A.; Gololobov, G.; Paul, S. Monoclonal antibody light chain with prothrombinase activity. Biochemistry 2000, 39, 6459–6465. [Google Scholar] [CrossRef]
- Lacroix-Desmazes, S.; Moreau, A.; Sooryanarayana; Bonnemain, C.; Stieltjes, N.; Pashov, A.; Sultan, Y.; Hoebeke, J.M.; Kazatchkine, D.; Kaveri, S.V. Catalytic activity of antibodies against factor VIII in patients with hemophilia A. Nat. Med. 1999, 5, 1044–1047. [Google Scholar] [CrossRef]
- Polosukhina, D.I.; Kanyshkova, T.G.; Doronin, B.M.; Tyshkevich, O.B.; Buneva, V.N.; Boiko, A.N.; Gusev, E.I.; Favorova, O.O.; Nevinsky, G.A. Hydrolysis of myelin basic protein by polyclonal catalytic IgGs from the sera of patients with multiple sclerosis. J. Cell Mol. Med. 2004, 8, 359–368. [Google Scholar] [CrossRef]
- Polosukhina, D.I.; Kanyshkova, T.G.; Doronin, B.M.; Tyshkevich, O.B.; Buneva, V.N.; Boiko, A.N.; Gusev, E.I.; Nevinsky, G.A.; Favorova, O.O. Metal-dependent hydrolysis of myelin basic protein by IgGs from the sera of patients with multiple sclerosis. Immunol. Lett. 2006, 103, 75–81. [Google Scholar] [CrossRef]
- Baranova, S.V.; Buneva, V.N.; Kharitonova, M.A.; Sizyakina, L.P.; Calmels, C.; Andreola, M.L.; Parissi, V.; Nevinsky, G.A. HIV-1 integrase-hydrolyzing antibodies from sera of HIV-infected patients. Biochimie 2009, 91, 1081–1086. [Google Scholar] [CrossRef]
- Paul, S.; Planque, S.A.; Nishiyama, Y.; Hanson, C.V.; Massey, R.J. Nature and nurture of catalytic antibodies. Adv. Exp. Med. Biol. 2012, 750, 56–75. [Google Scholar]
- Planque, S.A.; Nishiyama, Y.; Hara, M.; Sonoda, S.; Murphy, S.K.; Watanabe, K.; Mitsuda, Y.; Brown, E.L.; Massey, R.J.; Primmer, S.R.; et al. Physiological IgM class catalytic antibodies selective for transthyretin amyloid. J. Biol. Chem. 2014, 289, 13243–13258. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.; Kang, R.; Fan, X.-G.; Tang, D. Release and activity of histone in diseases. Cell Death. Dis. 2014, 5, e1370. [Google Scholar] [CrossRef] [Green Version]
- Baranova, S.V.; Buneva, V.N.; Nevinsky, G.A. Antibodies from the sera of HIV-infected patients efficiently hydrolyze all human histones. J. Mol. Recognit. 2016, 29, 346–362. [Google Scholar] [CrossRef] [PubMed]
- Baranova, S.V.; Dmitrienok, P.S.; Ivanisenko, N.V.; Buneva, V.N.; Nevinsky, G.A. Antibodies to H1 histone from the sera of HIV-infected patients recognize and catalyze site-specific degradation of this histone. J. Mol. Recognit. 2017, 30, e2588. [Google Scholar] [CrossRef] [PubMed]
- Baranova, S.V.; Dmitrienok, P.S.; Ivanisenko, N.V.; Buneva, V.N.; Nevinsky, G.A. Antibodies to H2a and H2b histones from the sera of HIV-infected patients catalyze site-specific degradation of these histones. Mol. Biosyst. 2017, 13, 1090–1101. [Google Scholar] [CrossRef] [PubMed]
- Baranova, S.V.; Dmitrenok, P.S.; Zubkova, A.D.; Ivanisenko, N.V.; Odintsova, E.S.; Buneva, V.N.; Nevinsky, G.A. Antibodies against H3 and H4 histones from the sera of HIV-infected patients catalyze site-specific degradation of these histones. J. Mol. Recognit. 2018, 31, e2703. [Google Scholar] [CrossRef]
- Baranova, S.V.; Mikheeva, E.V.; Buneva, V.N.; Nevinsky, G.A. Antibodies from the Sera of Multiple Sclerosis Patients Efficiently Hydrolyze Five Histones. Biomolecules 2019, 9, 741. [Google Scholar] [CrossRef] [Green Version]
- Baranova, S.V.; Dmitrienok, P.S.; Buneva, V.N.; Nevinsky, G.A. Autoantibodies in HIV-infected patients: Cross site-specific hydrolysis of H1 histone and myelin basic protein. Biofactors 2019, 45, 211–222. [Google Scholar] [CrossRef]
- Baranova, S.V.; Dmitrienok, P.S.; Buneva, V.N.; Nevinsky, G.A. HIV-Infected Patients: Cross Site-Specific Hydrolysis of H2a and H2b Histones and Myelin Basic Protein with Antibodies against These Three Proteins. Biomolecules 2020, 10, 1501. [Google Scholar] [CrossRef]
- Baranova, S.V.; Dmitrienok, P.S.; Buneva, V.N.; Sedykh, S.E.; Nevinsky, G.A. HIV-Infected Patients: Cross Site-Specific Hydrolysis of H3 and H4 Histones and Myelin Basic Protein with Antibodies against These Three Proteins. Molecules 2021, 26, 316. [Google Scholar] [CrossRef]
- Bezuglova, A.V.; Konenkova, L.P.; Doronin, B.M.; Buneva, V.N.; Nevinsky, G.A. Affinity and catalytic heterogeneity and metal-dependence of polyclonal myelin basic protein-hydrolyzing IgGs from sera of patients with systemic lupus erythematosus. J. Mol. Recognit. 2011, 24, 960–974. [Google Scholar] [CrossRef]
- Parshukova, D.L.; Smirnova, P.; Ermakov, E.A.; Bokhan, N.A.; Semke, A.V.; Ivanova, S.A.; Buneva, V.N.; Nevinsky, G.A. Autoimmunity and immune system dysregulation in schizophrenia: IgGs from sera of patients hydrolyze myelin basic protein. J. Mol. Recognit. 2019, 32, e2759. [Google Scholar] [CrossRef]
- Aulova, K.S.; Toporkova, L.B.; Lopatnikova, J.A.; Alshevskaya, A.A.; Sedykh, S.E.; Buneva, V.N.; Budde, T.; Meuth, S.G.; Popova, N.A.; Orlovskaya, I.A.; et al. Changes in cell differentiation and proliferation lead to production of abzymes in EAE mice treated with DNA-Histone complexes. J. Cell Mol. Med. 2018, 22, 5816–5832. [Google Scholar] [CrossRef] [Green Version]
- Founel, S.; Muller, S. Antinucleosome antibodies and T-cell response in systemic lupus erythematosus. Ann. Med. Interne 2002, 153, 513–519. [Google Scholar]
- Nevinsky, G.A. Structural, thermodynamic, and kinetic basis of DNA- and RNA-dependent enzymes functioning: Important role of weak nonspecific additive interactions between enzymes and long nucleic acids for their recognition and transformation. In Protein Structures: Kaleidoscope of Structural Properties and Functions; Uversky, V.N., Ed.; Research Signpost: Kerala, India, 2003; pp. 133–222. [Google Scholar]
- Nevinsky, G.A. Structural, thermodynamic, and kinetic basis for the activities of some nucleic acid repair enzymes. J. Mol. Recognit. 2011, 24, 656–677. [Google Scholar] [CrossRef]
- Nevinsky, G.A. How Enzymes, Proteins, and Antibodies Recognize Extended DNAs, General Regularities. Int. J. Mol. Sci. 2021, 22, 1369. [Google Scholar] [CrossRef]
- Nemazee, D. Receptor editing in lymphocyte development and central tolerance. Nat. Rev. Immunol. 2006, 6, 728–740. [Google Scholar] [CrossRef]
- Diaz, M.; Casali, P. Somatic immunoglobulin hypermutation. Curr. Opin. Immunol. 2002, 14, 235–240. [Google Scholar] [CrossRef] [Green Version]
- Honjo, T.; Habu, S. Origin of immune diversity: Genetic variation and selection. Annu. Rev. Bioch. 1985, 54, 803–830. [Google Scholar] [CrossRef]
- Nevinsky, G.A. The extreme diversity of autoantibodies and abzymes against different antigens in patients with various autoimmune diseases. In Advances in Medicine and Biology; Nova Science Publishers, Inc.: New York, NY, USA, 2021; pp. 1–130. [Google Scholar]
- Zhou, Z.H.; Tzioufas, A.G.; Notkins, A.L. Properties and function of polyreactive antibodies and polyreactive antigen-binding B cells. J. Autoimmun. 2007, 29, 219–228. [Google Scholar] [CrossRef] [Green Version]
- James, L.C.; Roversi, P.; Tawfik, D.S. Antibody multispecificity mediated by conformational diversity. Science 2003, 299, 1362–1367. [Google Scholar] [CrossRef] [Green Version]
- James, L.C.; Tawfik, D.S. Conformational diversity and protein evolution—A 60-year-old hypothesis revisited. Trends Biochem. Sci. 2003, 28, 361–368. [Google Scholar] [CrossRef]
- James, L.C.; Tawfik, D.S. The specificity of cross-reactivity: Promiscuous antibody binding involves specific hydrogen bonds rather than nonspecific hydrophobic stickiness. Protein Sci. 2003, 12, 2183–2193. [Google Scholar] [CrossRef] [Green Version]
- Timofeeva, A.M.; Buneva, V.N.; Nevinsky, G.A. Systemic lupus erythematosus: Molecular cloning and analysis of 22 individual recombinant monoclonal kappa light chains specifically hydrolyzing human myelin basic protein. J. Mol. Recognit. 2015, 28, 614–627. [Google Scholar] [CrossRef]
- Timofeeva, A.M.; Buneva, V.N.; Nevinsky, G.A. Systemic lupus erythematosus: Molecular cloning and analysis of recombinant monoclonal kappa light chain NGTA1-Me-pro with two metalloprotease active centers. Mol. Biosyst. 2016, 12, 3556–3566. [Google Scholar] [CrossRef]
- Timofeeva, A.M.; Ivanisenko, N.V.; Buneva, V.N.; Nevinsky, G.A. Systemic lupus erythematosus: Molecular cloning and analysis of recombinant monoclonal kappa light chain NGTA2-Me-pro-Tr possessing two different activities-trypsin-like and metalloprotease. Int. Immunol. 2015, 27, 633–645. [Google Scholar] [CrossRef] [Green Version]
- Bezuglova, A.M.; Buneva, V.N.; Nevinsky, G.A. Systemic lupus erythematosus: Monoclonal light chains of immunoglobulins against myelin basic protein, have proteolytic and DNase activity. Rus. J. Immunol. 2011, 5, 215–225. [Google Scholar]
- Timofeeva, A.M.; Nevinsky, G.A. Systemic lupus erythematosus: Possible localization of trypsin-like and metalloprotease active centers in the protein sequence of the monoclonal light chain (NGTA2-Me-pro-Tr). Biotechnol. Appl. Biochem. 2020, 67, 946–959. [Google Scholar] [CrossRef]
- Timofeeva, A.M.; Dmitrenok, P.S.; Konenkova, L.P.; Buneva, V.N.; Nevinsky, G.A. Multiple sites of the cleavage of 21- and 25-mer encephalytogenic oligopeptides corresponding to human myelin basic protein (MBP) by specific anti-MBP antibodies from patients with systemic lupus erythematosus. PLoS ONE 2013, 8, e51600. [Google Scholar] [CrossRef]
- Timofeeva, A.M.; Buneva, V.N.; Nevinsky, G.A. Systemic Lupus Erythematosus: Localization of DNase, Trypsin-Like, and Metalloprotease Active Centers in the Protein Sequence of NGTA3-Pro-Dnase Monoclonal Light Chain of Human Antibodies. J. Mol. Biol. Mol. Imaging 2020, 6, 1031. [Google Scholar]
- McDonald, W.I.; Compston, A.; Edan, G.; Goodkin, D.; Hartung, H.P.; Lublin, F.D.; McFarland, H.F.; Paty, D.W.; Polman, C.H.; Reingold, S.C.; et al. Recommended diagnostic criteria for multiple sclerosis: Guidelines from the International Panel on the Diagnosis of Multiple Sclerosis. Ann. Neurol. 2001, 50, 121–127. [Google Scholar] [CrossRef]
- Kurtzke, J.F. Rating neurological impairment in multiple sclerosis: An expanded disability scale (EDSS). Neurology 1983, 33, 1444–1452. [Google Scholar] [CrossRef] [Green Version]
- Krasitskaya, V.V.; Chaukina, V.V.; Abroskina, M.V.; Vorobyeva, M.A.; Ilminskaya, A.A.; Kabilov, M.R.; Prokopenko, S.V.; Nevinsky, G.A.; Venyaminova, A.G.; Frank, L.A. Bioluminescent aptamer-based sandwich-type assay of anti-myelin basic protein autoantibodies associated with multiple sclerosis. Anal. Chim. Acta 2019, 1064, 112–118. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of IgGs | ||||||
|---|---|---|---|---|---|---|
| Anti-H2A | Anti-H1 | Anti-H2B | Anti-H3 | Anti-H4 | Anti-MBP | Anti-DNA |
| 12 Sites * | 19 Sites | 11 Sites | 12 Sites | 9 Sites | 10 Sites | 23 Sites |
| - | - | - | - | - | - | K5-Q6 |
| K9-A10 | - | - | - | - | - | K9-10A |
| R11-A12 | - | - | R11-A12 | R11-A12 | - | R11-12A |
| K13-14A | - | - | K13-A14 | - | - | K13-14A |
| K15-16S | - | K15-S16 | K15-S16 | - | - | K15-S16 |
| - | S16-R17 | - | - | - | - | - |
| S16-R17 | - | - | - | - | - | - |
| - | - | R17-S18 | R17-S18 | - | - | R17-S18 |
| - | G22-L23 | - | - | - | - | - |
| R20-A21 | - | R20-A21 | R20-A21 | - | - | R20-A21 |
| R29-30V | - | - | - | - | - | R29-30V |
| R32-L33 | R32-L33 | R32-L33 | R32-L33 | R32-L33 | R32-L33 | R32-L33 |
| - | - | - | - | - | - | L34-R33 |
| - | - | - | - | L34-R35 | - | |
| R35-K36 | - | R35-K36 | R35-K36 | R35-K36 | R35-R36 | R35-K36 |
| - | - | - | - | - | - | K36-37G |
| - | E41-R42 | - | - | - | E41-R42 | - |
| R42-43V | - | - | - | - | R42-V43 | R42-43V |
| - | V43-G44 | - | - | - | - | - |
| - | - | G46-A47 | G46-A47 | G46-A47 | - | G46-A47 |
| - | - | Y50-L51 | - | - | - | - |
| - | L55-E56 | - | - | - | - | - |
| - | E56-Y57 | - | - | - | - | - |
| - | Y57-L58 | - | - | - | - | - |
| - | - | - | - | - | A60-E61 | |
| - | - | - | - | - | E61-I62 | E61-I62 |
| - | E64-L65 | - | - | - | E64-L65 | - |
| - | G67-N68 | - | - | - | - | - |
| - | - | - | - | - | R71-D72 | |
| - | K74-K75 | - | K74-K75 | - | - | K74-K75 |
| - | K75-T76 | - | - | - | - | K75-T76 |
| R77-I78 | R77-I78 | R77-I78 | R77-I78 | R77-I78 | R77-I78 | R77-I78 |
| - | - | - | - | - | - | - |
| - | P80-R81 | - | - | - | - | - |
| R81-H82 | - | R81-H82 | R81-H82 | R81-H82 | R81-Y82 | R81-H82 |
| - | H82-L83 | - | - | - | - | - |
| - | Q84-L85 | - | - | - | - | - |
| - | A86-I87 | - | - | - | - | |
| - | - | - | R88-N89 | R88-N89 | - | - |
| - | - | - | - | G98-R99 | ||
| - | - | T101-I102 | T101-I102 | T101-I102 | - | - |
| - | - | K119-T120 | - | - | - | - |
| - | T120-E121 | - | T120-E121 | - | - | - |
| - | - | E121-S122 | E121-S122 | E121-S122 | - | - |
| - | A126-K127 | - | - | - | - | A126-K127 |
| - | - | - | - | - | - | K127-G128 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nevinsky, G.A.; Buneva, V.N.; Dmitrienok, P.S. Multiple Sclerosis: Enzymatic Cross Site-Specific Recognition and Hydrolysis of H2A Histone by IgGs against H2A, H1, H2B, H3 Histones, Myelin Basic Protein, and DNA. Biomedicines 2022, 10, 1876. https://doi.org/10.3390/biomedicines10081876
Nevinsky GA, Buneva VN, Dmitrienok PS. Multiple Sclerosis: Enzymatic Cross Site-Specific Recognition and Hydrolysis of H2A Histone by IgGs against H2A, H1, H2B, H3 Histones, Myelin Basic Protein, and DNA. Biomedicines. 2022; 10(8):1876. https://doi.org/10.3390/biomedicines10081876
Chicago/Turabian StyleNevinsky, Georgy A., Valentina N. Buneva, and Pavel S. Dmitrienok. 2022. "Multiple Sclerosis: Enzymatic Cross Site-Specific Recognition and Hydrolysis of H2A Histone by IgGs against H2A, H1, H2B, H3 Histones, Myelin Basic Protein, and DNA" Biomedicines 10, no. 8: 1876. https://doi.org/10.3390/biomedicines10081876