
The Relationship between Stress, Inflammation, and Depression



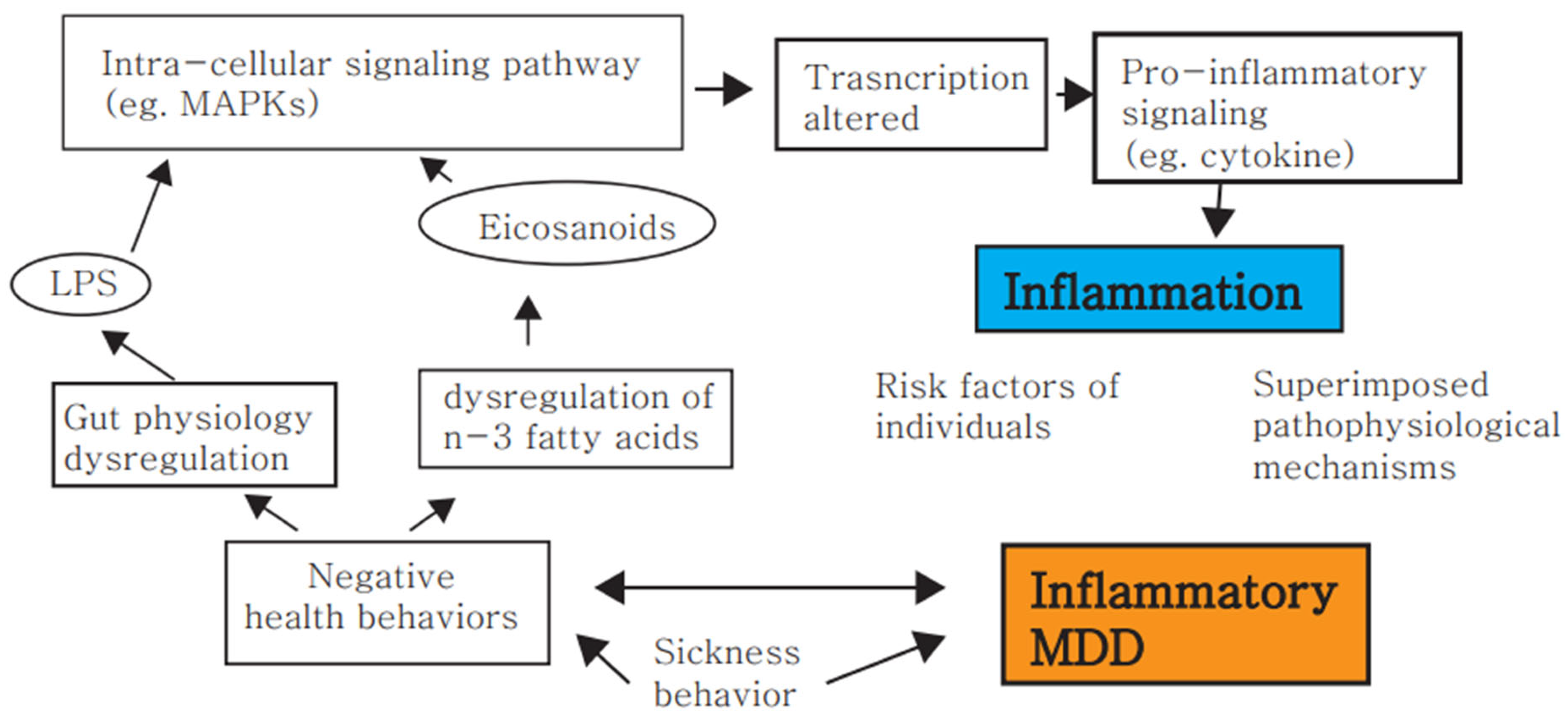{kind=link}
Abstract
:1. Introduction
2. Stress and Inflammation
3. Stress, Inflammation and Depression
4. MDD Is Associated with an Activated Immune System
5. Antidepressants Exert Immunomodulatory Effects
6. Non-Pharmacological Treatment Effects on Inflammatory Depression
7. Limitations and Future Directions
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shen, W.W. Clinical Psychopharmacology for the 21st Century, 3rd ed.; Ho-Chi Publishing Company: Taipei, Taiwan, 2011. [Google Scholar]
- Drazinix, C.M.; Sazbo, S.T. Neurotramitters and Receptors in Psychiatric Disorder (Chapter 2). In The American Psychiatric Association Publishing Textbook of Psychopharmacology; Schatzberg, A.F., Nemeroff, C.B., Eds.; APA Publishing: Washington, DC, USA, 2017. [Google Scholar]
- Jessberger, S.; Kempermann, G. Adult-born hippocampal neurons mature into activity-dependent responsiveness. Eur. J. Neurosci. 2003, 18, 2707–2712. [Google Scholar] [CrossRef]
- Landsbergis, P.A. The changing organization of work and the safety and health of working people: A commentary. J. Occup. Environ Med. 2003, 45, 61–72. [Google Scholar] [CrossRef]
- Molina, P.E. Rethinking Integration of Environmental and Behavioral Stressors; Back to Energy Homeostasis and Function. Function 2022, 3, zqab074. [Google Scholar] [CrossRef]
- Cohen, S.; Janicki-Deverts, D.; Miller, G.E. Psychological stress and disease. JAMA 2007, 298, 1685–1687. [Google Scholar] [CrossRef]
- Cohen, S.; Gianaros, P.J.; Manuck, S.B. A stage model of stress and disease. Perspect. Psychol. Sci. 2016, 11, 456–463. [Google Scholar] [CrossRef]
- Turner, A.I.; Smyth, N.; Hall, S.J.; Torres, S.J.; Hussein, M.; Jayasinghe, S.U.; Ball, K.; Clow, A.J. Psychological stress reactivity and future health and disease outcomes: A systematic review of prospective evidence. Psychoneuroendocrinology 2020, 114, 104599. [Google Scholar] [CrossRef]
- Rohleder, N. Stimulation of systemic low-grade inflammation by psychosocial stress. Psychosom. Med. 2014, 76, 181–189. [Google Scholar] [CrossRef]
- Rohleder, N. Stress and inflammation—The need to address the gap in the transition between acute and chronic stress effects. Psychoneuroendocrinology 2019, 105, 164–171. [Google Scholar] [CrossRef]
- Viljoen, M.; Lee Thomas neé Negrao, B. Low-grade systemic inflammation and the workplace. Work 2021, 69, 903–915. [Google Scholar] [CrossRef]
- Hantsoo, L.; Kornfield, S.; Anguera, M.C.; Epperson, C.N. Inflammation: A proposed intermediary between maternal stress and offspring neuropsychiatric risk. Biol. Psychiatr. 2019, 85, 97–106. [Google Scholar] [CrossRef]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W. Chronic inflammation in the etiology of disease across the life span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.G.; Hendricks, S.E.; Johnson, D.R.; Wieseler, J.; Burke, W.J. Antidepressants augment natural killer cell activity: In vivo and in vitro. Neuropsychobiology 1999, 39, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Calcia, M.A.; Bonsall, D.R.; Bloomfield, P.S.; Selvaraj, S.; Barichello, T.; Howes, O.D. Stress and neuroinflammation: A systematic review of the effects of stress on microglia and the implications for mental illness. Psychopharmacology 2016, 233, 1637–1650. [Google Scholar] [CrossRef] [Green Version]
- Picca, A.; Calvani, R.; Coelho-Junior, H.J.; Landi, F.; Bernabei, R.; Marzetti, E. Mitochondrial dysfunction, oxidative stress, and neuroinflammation: Intertwined roads to neurodegeneration. Antioxidants 2020, 9, 647. [Google Scholar] [CrossRef]
- Black, P.H. Stress and the inflammatory response: A review of neurogenic inflammation. Brain Behav. Immun. 2002, 16, 622–653. [Google Scholar] [CrossRef]
- Liu, Y.-Z.; Wang, Y.-X.; Jiang, C.-L. Inflammation: The common pathway of stress-related diseases. Front. Hum. Neurosci. 2017, 11, 316. [Google Scholar] [CrossRef]
- Liezmann, C.; Klapp, B.; Peters, E. Stress, atopy and allergy: A re-evaluation from a psychoneuroimmunologic persepective. Dermato Endocrinology 2011, 3, 37–40. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.-L.; Lu, C.-L.; Liu, X.-Y. The molecular basis for bidirectional communication between the immune and neuroendocrine systems. Domest. Anim. Endocrinol. 1998, 15, 363–369. [Google Scholar] [CrossRef]
- Quan, N.; Banks, W.A. Brain-immune communication pathways. Brain Behav. Immun. 2007, 21, 727–735. [Google Scholar] [CrossRef]
- Arambula, S.E.; McCarthy, M.M. Neuroendocrine-immune crosstalk shapes sex-specific brain development. Endocrinology 2020, 161, bqaa055. [Google Scholar] [CrossRef] [Green Version]
- Quatrini, L.; Vivier, E.; Ugolini, S. Neuroendocrine regulation of innate lymphoid cells. Immunol. Rev. 2018, 286, 120–136. [Google Scholar] [CrossRef] [Green Version]
- Weigent, D.A.; Blalock, J.E. Associations between the neuroendocrine and immune systems. J. Leukoc. Biol. 1995, 58, 137–150. [Google Scholar] [CrossRef]
- Weigent, D.A.; Carr, D.; Blalock, J.E. Bidirectional communication between the neuroendocrine and immune systems. Common hormones and hormone receptors. Ann. N. Y. Acad. Sci. 1990, 579, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Gallelli, L.; D’Agostino, B.; Vatrella, A.; Fratto, D.; Renda, T.; Galderisi, U.; Piegari, E.; Crimi, N.; Rossi, F.; Caputi, M. Effects of TGF-beta and glucocorticoids on map kinase phosphorylation, IL-6/IL-11 secretion and cell proliferation in primary cultures of human lung fibroblasts. J. Cell. Physiol. 2007, 210, 489–497. [Google Scholar]
- Gallelli, L.; Pelaia, G.; Fratto, D.; Muto, V.; Falcone, D.; Vatrella, A.; Curto, L.; Renda, T.; Busceti, M.; Liberto, M. Effects of budesonide on p38 MAPK activation, apoptosis and IL-8 secretion, induced by TNF-α and Haemophilus influenzae in human bronchial epithelial cells. Int. J. Immunopathol. Pharmacol. 2010, 23, 471–479. [Google Scholar] [CrossRef]
- Sorrells, S.F.; Caso, J.R.; Munhoz, C.D.; Sapolsky, R.M. The stressed CNS: When glucocorticoids aggravate inflammation. Neuron 2009, 64, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Hill, A.R.; Spencer-Segal, J.L. Glucocorticoids and the brain after critical illness. Endocrinology 2021, 162, bqaa242. [Google Scholar] [CrossRef]
- Liberman, A.C.; Trias, E.; da Silva Chagas, L.; Trindade, P.; dos Santos Pereira, M.; Refojo, D.; Hedin-Pereira, C.; Serfaty, C.A. Neuroimmune and inflammatory signals in complex disorders of the central nervous system. Neuroimmunomodulation 2018, 25, 246–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besedovsky, H.O.; Del Rey, A. Regulating inflammation by glucocorticoids. Nat. Immunol. 2006, 7, 537. [Google Scholar] [CrossRef]
- Duque, E.D.A.; Munhoz, C.D. The pro-inflammatory effects of glucocorticoids in the brain. Front. Endocrinol. 2016, 7, 78. [Google Scholar] [CrossRef] [Green Version]
- Sorrells, S.F.; Munhoz, C.D.; Manley, N.C.; Yen, S.; Sapolsky, R.M. Glucocorticoids increase excitotoxic injury and inflammation in the hippocampus of adult male rats. Neuroendocrinology 2014, 100, 129–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elenkov, I.J. Neurohormonal-cytokine interactions: Implications for inflammation, common human diseases and well-being. Neurochem. Int. 2008, 52, 40–51. [Google Scholar] [CrossRef] [PubMed]
- van den Heuvel, L.L.; Suliman, S.; Bröcker, E.; Kilian, S.; Stalder, T.; Kirschbaum, C.; Seedat, S. The association between hair cortisol levels, inflammation and cognitive functioning in females. Psychoneuroendocrinology 2022, 136, 105619. [Google Scholar] [CrossRef]
- Cruz-Topete, D.; Cidlowski, J.A. One hormone, two actions: Anti-and pro-inflammatory effects of glucocorticoids. Neuroimmunomodulation 2015, 22, 20–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oppong, E.; Cato, A.C. Effects of glucocorticoids in the immune system. Adv. Exp. Med. Biol. 2015, 872, 217–233. [Google Scholar]
- Frank, M.G.; Thompson, B.M.; Watkins, L.R.; Maier, S.F. Glucocorticoids mediate stress-induced priming of microglial pro-inflammatory responses. Brain Behav. Immun. 2012, 26, 337–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cain, D.W.; Cidlowski, J.A. Immune regulation by glucocorticoids. Nat. Rev. Immunol. 2017, 17, 233–247. [Google Scholar] [CrossRef]
- Busillo, J.M.; Azzam, K.M.; Cidlowski, J.A. Glucocorticoids sensitize the innate immune system through regulation of the NLRP3 inflammasome. J. Biol. Chem. 2011, 286, 38703–38713. [Google Scholar] [CrossRef] [Green Version]
- Gülke, E.; Gelderblom, M.; Magnus, T. Danger signals in stroke and their role on microglia activation after ischemia. Ther. Adv. Neurol. Disord. 2018, 11, 1756286418774254. [Google Scholar] [CrossRef] [Green Version]
- Alley, D.E.; Seeman, T.E.; Kim, J.K.; Karlamangla, A.; Hu, P.; Crimmins, E.M. Socioeconomic status and C-reactive protein levels in the US population: NHANES IV. Brain Behav. Immun. 2006, 20, 498–504. [Google Scholar] [CrossRef]
- Danese, A.; Pariante, C.M.; Caspi, A.; Taylor, A.; Poulton, R. Childhood maltreatment predicts adult inflammation in a life-course study. Proc. Natl. Acad. Sci. USA 2007, 104, 1319–1324. [Google Scholar] [CrossRef] [Green Version]
- Steptoe, A.; Hamer, M.; Chida, Y. The effects of acute psychological stress on circulating inflammatory factors in humans: A review and meta-analysis. Brain Behav. Immun. 2007, 21, 901–912. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.E.; Chen, E.; Sze, J.; Marin, T.; Arevalo, J.M.; Doll, R.; Ma, R.; Cole, S.W. A functional genomic fingerprint of chronic stress in humans: Blunted glucocorticoid and increased NF-κB signaling. Biol. Psychiatr. 2008, 64, 266–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elenkov, I.J.; Chrousos, G.P. Stress system–organization, physiology and immunoregulation. Neuroimmunomodulation 2006, 13, 257–267. [Google Scholar] [CrossRef]
- Papargyri, P.; Zapanti, E.; Salakos, N.; Papargyris, L.; Bargiota, A.; Mastorakos, G. Links between HPA axis and adipokines: Clinical implications in paradigms of stress-related disorders. Expert Rev. Endocrinol. Metab. 2018, 13, 317–332. [Google Scholar] [CrossRef]
- Borghetti, P.; Saleri, R.; Mocchegiani, E.; Corradi, A.; Martelli, P. Infection, immunity and the neuroendocrine response. Vet. Immunol. Immunopathol. 2009, 130, 141–162. [Google Scholar] [CrossRef] [PubMed]
- Sterling, P. Allostasis: A new paradigm to explain arousal pathology. In Handbook of Life Stress, Cognition and Health; John Wiley & Sons: New York, NY, USA, 1988; pp. 629–649. [Google Scholar]
- Lee, S.W. A Copernican approach to brain advancement: The paradigm of allostatic orchestration. Front. Hum. Neurosci. 2019, 13, 129. [Google Scholar] [CrossRef]
- Bellinger, D.L.; Millar, B.A.; Perez, S.; Carter, J.; Wood, C.; ThyagaRajan, S.; Molinaro, C.; Lubahn, C.; Lorton, D. Sympathetic modulation of immunity: Relevance to disease. Cell Immunol. 2008, 252, 27–56. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.-R.; Xu, Z.; Jiang, C.-L. Neuropeptide Y promotes TGF-β1 production in RAW264. 7 cells by activating PI3K pathway via Y1 receptor. Neurosci. Bull. 2008, 24, 155–159. [Google Scholar] [CrossRef]
- Huang, J.-L.; Zhang, Y.-L.; Wang, C.-C.; Zhou, J.-R.; Ma, Q.; Wang, X.; Shen, X.-H.; Jiang, C.-L. Enhanced phosphorylation of MAPKs by NE promotes TNF-α production by macrophage through α adrenergic receptor. Inflammation 2012, 35, 527–534. [Google Scholar] [CrossRef]
- Bellinger, D.L.; Lorton, D. Sympathetic nerve hyperactivity in the spleen: Causal for nonpathogenic-driven chronic immune-mediated inflammatory diseases (IMIDs)? Int. J. Mol. Sci. 2018, 19, 1188. [Google Scholar] [CrossRef] [Green Version]
- Brinkman, D.J.; Ten Hove, A.S.; Vervoordeldonk, M.J.; Luyer, M.D.; de Jonge, W.J. Neuroimmune interactions in the gut and their significance for intestinal immunity. Cells 2019, 8, 670. [Google Scholar] [CrossRef] [Green Version]
- Song, C.; Kenis, G.; van Gastel, A.; Bosmans, E.; Lin, A.; de Jong, R.; Neels, H.; Scharpé, S.; Janca, A.; Yasukawa, K. Influence of psychological stress on immune-inflammatory variables in normal humans. Part II. Altered serum concentrations of natural anti-inflammatory agents and soluble membrane antigens of monocytes and T lymphocytes. Psychiatr. Res. 1999, 85, 293–303. [Google Scholar] [CrossRef]
- Miller, A.H.; Maletic, V.; Raison, C.L. Inflammation and its discontents: The role of cytokines in the pathophysiology of major depression. Biol. Psychiatr. 2009, 65, 732–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, W.; Feng, R.; Yang, Y. Changes in the serum levels of inflammatory cytokines in antidepressant drug-naïve patients with major depression. PLoS ONE 2018, 13, e0197267. [Google Scholar] [CrossRef] [PubMed]
- Draganov, M.; Arranz, M.J.; Salazar, J.; de Diego-Adeliño, J.; Gallego-Fabrega, C.; Jubero, M.; Carceller-Sindreu, M.; Portella, M.J. Association study of polymorphisms within inflammatory genes and methylation status in treatment response in major depression. Eur. Psychiatr. 2019, 60, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Opel, N.; Cearns, M.; Clark, S.; Toben, C.; Grotegerd, D.; Heindel, W.; Kugel, H.; Teuber, A.; Minnerup, H.; Berger, K. Large-scale evidence for an association between low-grade peripheral inflammation and brain structural alterations in major depression in the BiDirect study. J. Psychiatr. Neurosci. 2019, 44, 423–431. [Google Scholar] [CrossRef] [Green Version]
- Halaris, A. Inflammation and depression but where does the inflammation come from? Curr. Opin. Psychiatr. 2019, 32, 422–428. [Google Scholar] [CrossRef]
- Rhie, S.J.; Jung, E.-Y.; Shim, I. The role of neuroinflammation on pathogenesis of affective disorders. J. Exerc. Rehabil. 2020, 16, 2. [Google Scholar] [CrossRef] [Green Version]
- García-Bueno, B.; Caso, J.R.; Leza, J.C. Stress as a neuroinflammatory condition in brain: Damaging and protective mechanisms. Neurosci. Biobehav. Rev. 2008, 32, 1136–1151. [Google Scholar] [CrossRef]
- Gu, M.; Mei, X.-L.; Zhao, Y.-N. Sepsis and cerebral dysfunction: BBB damage, neuroinflammation, oxidative stress, apoptosis and autophagy as key mediators and the potential therapeutic approaches. Neurotox. Res. 2021, 39, 489–503. [Google Scholar] [CrossRef]
- He, J.; Zhu, G.; Wang, G.; Zhang, F. Oxidative stress and neuroinflammation potentiate each other to promote progression of dopamine neurodegeneration. Oxid. Med. Cell Longev. 2020, 6137521. [Google Scholar] [CrossRef] [PubMed]
- Solleiro-Villavicencio, H.; Rivas-Arancibia, S. Effect of chronic oxidative stress on neuroinflammatory response mediated by CD4+ T cells in neurodegenerative diseases. Front. Cell Neurosci. 2018, 12, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munhoz, C.; Garcia-Bueno, B.; Madrigal, J.; Lepsch, L.; Scavone, C.; Leza, J. Stress-induced neuroinflammation: Mechanisms and new pharmacological targets. Braz. J. Med. Biol. Res. 2008, 41, 1037–1046. [Google Scholar] [CrossRef]
- Gárate, I.; Garcia-Bueno, B.; Madrigal, J.L.M.; Caso, J.R.; Alou, L.; Gomez-Lus, M.L.; Micó, J.A.; Leza, J.C. Stress-induced neuroinflammation: Role of the Toll-like receptor-4 pathway. Biol. Psychiatr. 2013, 73, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.M.; Main, B.S.; Crack, P.J. Neuroinflammation and oxidative stress: Co-conspirators in the pathology of Parkinson’s disease. Neurochem. Int. 2013, 62, 803–819. [Google Scholar] [CrossRef] [PubMed]
- Hori, H.; Kim, Y. Inflammation and post-traumatic stress disorder. Psychiatr. Clin. Neurosci. 2019, 73, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.; Campisi, J.; Sharkey, C.; Kennedy, S.; Nickerson, M.; Greenwood, B.; Fleshner, M. Catecholamines mediate stress-induced increases in peripheral and central inflammatory cytokines. Neuroscience 2005, 135, 1295–1307. [Google Scholar] [CrossRef]
- Johnson, J.D.; Barnard, D.F.; Kulp, A.C.; Mehta, D.M. Neuroendocrine regulation of brain cytokines after psychological stress. J. Endocr. Soc. 2019, 3, 1302–1320. [Google Scholar] [CrossRef]
- Barnard, D.F. The Regulation of Brain Pro-Inflammatory Cytokines: Implications for Stress and Depression. Ph.D. Thesis, Kent State University, Kent, OH, USA, 2020. [Google Scholar]
- Foertsch, S.; Reber, S.O. The role of physical trauma in social stress-induced immune activation. Neurosci. Biobehav. Rev. 2020, 113, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Wohleb, E.S.; Franklin, T.; Iwata, M.; Duman, R.S. Integrating neuroimmune systems in the neurobiology of depression. Nat. Rev. Neurosci. 2016, 17, 497–511. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.D.; Godbout, J.P.; Sheridan, J.F. Repeated social defeat, neuroinflammation, and behavior: Monocytes carry the signal. Neuropsychopharmacology 2017, 42, 46–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, K.; Fornaguera-Trías, J.; Sheridan, J.F. Stress-induced microglia activation and monocyte trafficking to the brain underlie the development of anxiety and depression. Inflamm. Assoc. Depress. Evid. Mech. Implic. 2016, 31, 155–172. [Google Scholar]
- Mendiola, A.S.; Ryu, J.K.; Bardehle, S.; Meyer-Franke, A.; Ang, K.K.-H.; Wilson, C.; Baeten, K.M.; Hanspers, K.; Merlini, M.; Thomas, S. Transcriptional profiling and therapeutic targeting of oxidative stress in neuroinflammation. Nat. Immunol. 2020, 21, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Fonken, L.K.; Weber, M.D.; Daut, R.A.; Kitt, M.M.; Frank, M.G.; Watkins, L.R.; Maier, S.F. Stress-induced neuroinflammatory priming is time of day dependent. Psychoneuroendocrinology 2016, 66, 82–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, M.D.; Frank, M.G.; Tracey, K.J.; Watkins, L.R.; Maier, S.F. Stress induces the danger-associated molecular pattern HMGB-1 in the hippocampus of male Sprague Dawley rats: A priming stimulus of microglia and the NLRP3 inflammasome. J. Neurosci. 2015, 35, 316–324. [Google Scholar] [CrossRef]
- Wohleb, E.S.; Delpech, J.-C. Dynamic cross-talk between microglia and peripheral monocytes underlies stress-induced neuroinflammation and behavioral consequences. Prog. Neuropsychopharmacol. Biol. Psychiatr. 2017, 79, 40–48. [Google Scholar] [CrossRef]
- Park, S.-C. Neurogenesis and antidepressant action. Cell Tissue Res. 2019, 377, 95–106. [Google Scholar] [CrossRef]
- Kim, Y.-K.; Park, S.-C. An alternative approach to future diagnostic standards for major depressive disorder. Prog. Neuropsychopharmacol. Biol. Psychiatr. 2021, 105, 110133. [Google Scholar] [CrossRef]
- Kim, I.B.; Park, S.-C. Neural Circuitry–Neurogenesis Coupling Model of Depression. Int. J. Mol. Sci. 2021, 22, 2468. [Google Scholar] [CrossRef]
- Kim, I.B.; Park, S.-C. Machine Learning-Based Definition of Symptom Clusters and Selection of Antidepressants for Depressive Syndrome. Diagnostics 2021, 11, 1631. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.B.; Park, S.-C. The Entorhinal Cortex and Adult Neurogenesis in Major Depression. Int. J. Mol. Sci. 2021, 22, 11725. [Google Scholar] [CrossRef] [PubMed]
- Scrivo, R.; Vasile, M.; Bartosiewicz, I.; Valesini, G. Inflammation as “common soil” of the multifactorial diseases. Autoimmun. Rev. 2011, 10, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Martone, G. The inflammation hypothesis and mental illness. J. Clin. Psychiatr. Neurosci. 2019, 2, 3–12. [Google Scholar]
- Dolsen, M.R.; Prather, A.A.; Lamers, F.; Penninx, B.W. Suicidal ideation and suicide attempts: Associations with sleep duration, insomnia, and inflammation. Psychol. Med. 2021, 51, 2094–2103. [Google Scholar] [CrossRef]
- Brahadeeswaran, S.; Sivagurunathan, N.; Calivarathan, L. Inflammasome Signaling in the Aging Brain and Age-Related Neurodegenerative Diseases. Mol. Neurobiol. 2022, 59, 2288–2304. [Google Scholar] [CrossRef] [PubMed]
- Capuron, L.; Raison, C.L.; Musselman, D.L.; Lawson, D.H.; Nemeroff, C.B.; Miller, A.H. Association of exaggerated HPA axis response to the initial injection of interferon-alpha with development of depression during interferon-alpha therapy. Am. J. Psychiatr. 2003, 160, 1342–1345. [Google Scholar] [CrossRef]
- Davies, K.A.; Cooper, E.; Voon, V.; Tibble, J.; Cercignani, M.; Harrison, N.A. Interferon and anti-TNF therapies differentially modulate amygdala reactivity which predicts associated bidirectional changes in depressive symptoms. Mol. Psychiatr. 2021, 26, 5150–5160. [Google Scholar] [CrossRef]
- Su, K.-P.; Lai, H.-C.; Peng, C.-Y.; Su, W.-P.; Chang, J.P.-C.; Pariante, C.M. Interferon-alpha-induced depression: Comparisons between early-and late-onset subgroups and with patients with major depressive disorder. Brain Behav. Immun. 2019, 80, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Dooley, L.N.; Kuhlman, K.R.; Robles, T.F.; Eisenberger, N.I.; Craske, M.G.; Bower, J.E. The role of inflammation in core features of depression: Insights from paradigms using exogenously-induced inflammation. Neurosci. Biobehav. Rev. 2018, 94, 219–237. [Google Scholar] [CrossRef] [Green Version]
- Ma, K.; Zhang, H.; Baloch, Z. Pathogenetic and therapeutic applications of tumor necrosis factor-α (TNF-α) in major depressive disorder: A systematic review. Int. J. Mol. Sci. 2016, 17, 733. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Hernández, O.T.; Martínez-Mota, L.; Herrera-Pérez, J.J.; Jiménez-Rubio, G. Role of estradiol in the expression of genes involved in serotonin neurotransmission: Implications for female depression. Curr. Neuropharmacol. 2019, 17, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Baudry, A.; Pietri, M.; Launay, J.-M.; Kellermann, O.; Schneider, B. Multifaceted regulations of the serotonin transporter: Impact on antidepressant response. Front. Neurosci. 2019, 13, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Denna, T.H.; Storkersen, J.N.; Gerriets, V.A. Beyond a neurotransmitter: The role of serotonin in inflammation and immunity. Pharmacol. Res. 2019, 140, 100–114. [Google Scholar] [CrossRef]
- Smith, R.S. The macrophage theory of depression. Med. Hypotheses 1991, 35, 298–306. [Google Scholar] [CrossRef]
- Norman, G.; Karelina, K.; Zhang, N.; Walton, J.; Morris, J.; Devries, A. Stress and IL-1β contribute to the development of depressive-like behavior following peripheral nerve injury. Mol. Psychiatr. 2010, 15, 404–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.-L.; Han, Q.-Q.; Gong, W.-Q.; Pan, D.-H.; Wang, L.-Z.; Hu, W.; Yang, M.; Li, B.; Yu, J.; Liu, Q. Microglial activation mediates chronic mild stress-induced depressive-and anxiety-like behavior in adult rats. J. Neuroinflamm. 2018, 15, 198–209. [Google Scholar] [CrossRef] [Green Version]
- Miura, H.; Ozaki, N.; Sawada, M.; Isobe, K.; Ohta, T.; Nagatsu, T. A link between stress and depression: Shifts in the balance between the kynurenine and serotonin pathways of tryptophan metabolism and the etiology and pathophysiology of depression. Stress 2008, 11, 198–209. [Google Scholar] [CrossRef]
- Tanaka, M.; Tóth, F.; Polyák, H.; Szabó, Á.; Mándi, Y.; Vécsei, L. Immune influencers in action: Metabolites and enzymes of the Tryptophan-Kynurenine metabolic pathway. Biomedicines 2021, 9, 734. [Google Scholar] [CrossRef]
- Dowlati, Y.; Herrmann, N.; Swardfager, W.; Liu, H.; Sham, L.; Reim, E.K.; Lanctôt, K.L. A meta-analysis of cytokines in major depression. Biol. Psychiatr. 2010, 67, 446–457. [Google Scholar] [CrossRef]
- Raison, C.; Borisov, A.; Woolwine, B.; Massung, B.; Vogt, G.; Miller, A. Interferon-α effects on diurnal hypothalamic–pituitary–adrenal axis activity: Relationship with proinflammatory cytokines and behavior. Mol. Psychiatr. 2010, 15, 535–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bekhbat, M.; Treadway, M.T.; Felger, J.C. Inflammation as a Pathophysiologic Pathway to Anhedonia: Mechanisms and Therapeutic Implications; Springer: Berlin/Heidelberg, Germany, 2022. [Google Scholar]
- Bauer, M.E.; Teixeira, A.L. Inflammation in psychiatric disorders: What comes first? Ann. N. Y. Acad. Sci. 2019, 1437, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.-L.; Liu, Y.-N.; Liu, L.; Wang, X.; Jiang, C.-L.; Wang, Y.-X. Inducible nitric oxide synthase is involved in the modulation of depressive behaviors induced by unpredictable chronic mild stress. J. Neuroinflamm. 2012, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, C.J.; Huang, Y.; Wynne, A.; Hanke, M.; Himler, J.; Bailey, M.T.; Sheridan, J.F.; Godbout, J.P. Minocycline attenuates lipopolysaccharide (LPS)-induced neuroinflammation, sickness behavior, and anhedonia. J. Neuroinflamm. 2008, 5, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroder, K.; Tschopp, J. The inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ketelut-Carneiro, N.; Fitzgerald, K.A. Inflammasomes. Curr. Biol. 2020, 30, R689–R694. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, L.; Peng, Y.L.; Liu, Y.Z.; Wu, T.Y.; Shen, X.L.; Zhou, J.R.; Sun, D.Y.; Huang, A.J.; Wang, X. Involvement of inflammasome activation in lipopolysaccharide-induced mice depressive-like behaviors. CNS Neurosci. Ther. 2014, 20, 119–124. [Google Scholar] [CrossRef]
- Zhang, L.; Previn, R.; Lu, L.; Liao, R.-F.; Jin, Y.; Wang, R.-K. Crocin, a natural product attenuates lipopolysaccharide-induced anxiety and depressive-like behaviors through suppressing NF-kB and NLRP3 signaling pathway. Brain Res. Bull. 2018, 142, 352–359. [Google Scholar] [CrossRef]
- Wong, M.-L.; Inserra, A.; Lewis, M.; Mastronardi, C.A.; Leong, L.; Choo, J.; Kentish, S.; Xie, P.; Morrison, M.; Wesselingh, S. Inflammasome signaling affects anxiety-and depressive-like behavior and gut microbiome composition. Mol. Psychiatr. 2016, 21, 797–805. [Google Scholar] [CrossRef]
- Inserra, A.; Mastronardi, C.A.; Rogers, G.; Licinio, J.; Wong, M.-L. Neuroimmunomodulation in major depressive disorder: Focus on caspase 1, inducible nitric oxide synthase, and interferon-gamma. Mol. Neurobiol. 2019, 56, 4288–4305. [Google Scholar] [CrossRef] [Green Version]
- Carlessi, A.S.; Borba, L.A.; Zugno, A.I.; Quevedo, J.; Réus, G.Z. Gut microbiota–brain axis in depression: The role of neuroinflammation. Eur. J. Neurosci. 2021, 53, 222–235. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Xing, C.; Long, W.; Wang, H.Y.; Liu, Q.; Wang, R.-F. Impact of microbiota on central nervous system and neurological diseases: The gut-brain axis. J. Neuroinflamm. 2019, 16, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, R.J.; Freedland, K.E.; Clouse, R.E.; Lustman, P.J. The prevalence of comorbid depression in adults with diabetes: A meta-analysis. Diabetes Care. 2001, 24, 1069–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, S.; Stone, M.; Peters, J.; Davies, M.; Khunti, K. The prevalence of co-morbid depression in adults with Type 2 diabetes: A systematic review and meta-analysis. Diabet. Med. 2006, 23, 1165–1173. [Google Scholar] [CrossRef]
- Moulton, C.D.; Pickup, J.C.; Ismail, K. The link between depression and diabetes: The search for shared mechanisms. Lancet Diabetes Endocrinol. 2015, 3, 461–471. [Google Scholar] [CrossRef]
- Laake, J.-P.S.; Stahl, D.; Amiel, S.A.; Petrak, F.; Sherwood, R.A.; Pickup, J.C.; Ismail, K. The association between depressive symptoms and systemic inflammation in people with type 2 diabetes: Findings from the South London Diabetes Study. Diabetes Care 2014, 37, 2186–2192. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Fu, T.; Yin, R.; Zhang, Q.; Shen, B. Prevalence of depression and anxiety in systemic lupus erythematosus: A systematic review and meta-analysis. BMC Psychiatr. 2017, 17, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Van Exel, E.; Jacobs, J.; Korswagen, L.; Voskuyl, A.; Stek, M.; Dekker, J.; Bultink, I. Depression in systemic lupus erythematosus, dependent on or independent of severity of disease. Lupus 2013, 22, 1462–1469. [Google Scholar] [CrossRef]
- Xie, X.; Wu, D.; Chen, H. Prevalence and risk factors of anxiety and depression in patients with systemic lupus erythematosus in Southwest China. Rheumatol. Int. 2016, 36, 1705–1710. [Google Scholar] [CrossRef]
- Schmeding, A.; Schneider, M. Fatigue, health-related quality of life and other patient-reported outcomes in systemic lupus erythematosus. Best Pract. Res. Clin. Rheumatol. 2013, 27, 363–375. [Google Scholar] [CrossRef]
- Mak, A.; Tang, C.; Ho, R.C.-M. Serum tumour necrosis factor-alpha is associated with poor health-related quality of life and depressive symptoms in patients with systemic lupus erythematosus. Lupus 2013, 22, 254–261. [Google Scholar] [CrossRef]
- Quan, W.; An, J.; Li, G.; Qian, G.; Jin, M.; Feng, C.; Li, S.; Li, X.; Xu, Y.; Hu, X. Th cytokine profile in childhood-onset systemic lupus erythematosus. BMC Pediatr. 2021, 21, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.-C.; Guo, H.-R.; Lu, M.-C.; Livneh, H.; Lai, N.-S.; Tsai, T.-Y. Increased risk of depression in patients with rheumatoid arthritis: A seven-year population-based cohort study. Clinics 2015, 70, 91–96. [Google Scholar] [CrossRef]
- Matcham, F.; Rayner, L.; Steer, S.; Hotopf, M. The prevalence of depression in rheumatoid arthritis: A systematic review and meta-analysis. Rheumatology 2013, 52, 2136–2148. [Google Scholar] [CrossRef] [Green Version]
- Stebbings, S.; Treharne, G.J. Fatigue in rheumatic disease: An overview. Int. J. Clin. Rheumatol. 2010, 5, 487–502. [Google Scholar] [CrossRef]
- Kojima, M.; Kojima, T.; Suzuki, S.; Oguchi, T.; Oba, M.; Tsuchiya, H.; Sugiura, F.; Kanayama, Y.; Furukawa, T.A.; Tokudome, S. Depression, inflammation, and pain in patients with rheumatoid arthritis. Arthritis Care Res. 2009, 61, 1018–1024. [Google Scholar] [CrossRef] [PubMed]
- Madsen, S.G.; Danneskiold-Samsøe, B.; Stockmarr, A.; Bartels, E. Correlations between fatigue and disease duration, disease activity, and pain in patients with rheumatoid arthritis: A systematic review. Scand. J. Rheumatol. 2016, 45, 255–261. [Google Scholar] [CrossRef] [Green Version]
- Almeida, C.; Choy, E.H.; Hewlett, S.; Kirwan, J.R.; Cramp, F.; Chalder, T.; Pollock, J.; Christensen, R. Biologic interventions for fatigue in rheumatoid arthritis. Cochrane Database Syst. Rev. 2016, 2016, CD008334. [Google Scholar] [CrossRef]
- Yende, S.; D’Angelo, G.; Kellum, J.A.; Weissfeld, L.; Fine, J.; Welch, R.D.; Kong, L.; Carter, M.; Angus, D.C. Inflammatory markers at hospital discharge predict subsequent mortality after pneumonia and sepsis. Am. J. Respir. Crit. Care Med. 2008, 177, 1242–1247. [Google Scholar] [CrossRef] [Green Version]
- Shukla, P.; Rao, G.M.; Pandey, G.; Sharma, S.; Mittapelly, N.; Shegokar, R.; Mishra, P.R. Therapeutic interventions in sepsis: Current and anticipated pharmacological agents. Br. J. Pharmacol. 2014, 171, 5011–5031. [Google Scholar] [CrossRef] [Green Version]
- Prescott, H.C.; Angus, D.C. Enhancing recovery from sepsis: A review. JAMA 2018, 319, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Davydow, D.S.; Hough, C.L.; Langa, K.M.; Iwashyna, T.J. Symptoms of depression in survivors of severe sepsis: A prospective cohort study of older Americans. Am. J. Geriatr. Psychiatr. 2013, 21, 887–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, M.M.; Connor, T.J.; Harkin, A. Stress-related immune markers in depression: Implications for treatment. Int. J. Neuropsychopharmacol. 2016, 19, pyw001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojard, C.; Donnelly, J.P.; Safford, M.M.; Griffin, R. Psychosocial stress as a risk factor for sepsis: A population-based cohort study. Psychosom. Med. 2015, 77, 93. [Google Scholar] [CrossRef] [Green Version]
- Anderson, S.T.; Commins, S.; Moynagh, P.N.; Coogan, A.N. Lipopolysaccharide-induced sepsis induces long-lasting affective changes in the mouse. Brain Behav. Immun. 2015, 43, 98–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassol-Jr, O.J.; Comim, C.M.; Petronilho, F.; Constantino, L.S.; Streck, E.L.; Quevedo, J.; Dal-Pizzol, F. Low dose dexamethasone reverses depressive-like parameters and memory impairment in rats submitted to sepsis. Neurosci. Lett. 2010, 473, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Chiu, W.; Su, Y.; Su, K.; Chen, P. Recurrence of depressive disorders after interferon-induced depression. Transl. Psychiatr. 2017, 7, e1026. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Xu, S.; Zhang, Z.; Zhang, Y.; Wu, Y.; An, J.; Lin, J.; Yuan, Z.; Shen, L.; Si, T. Identification of key genes and the pathophysiology associated with major depressive disorder patients based on integrated bioinformatics analysis. Front. Psychiatr. 2020, 11, 192. [Google Scholar] [CrossRef] [Green Version]
- Ohgi, Y.; Futamura, T.; Kikuchi, T.; Hashimoto, K. Effects of antidepressants on alternations in serum cytokines and depressive-like behavior in mice after lipopolysaccharide administration. Pharmacol. Biochem. Behav. 2013, 103, 853–859. [Google Scholar] [CrossRef]
- Ramirez, K.; Shea, D.T.; McKim, D.B.; Reader, B.F.; Sheridan, J.F. Imipramine attenuates neuroinflammatory signaling and reverses stress-induced social avoidance. Brain Behav. Immun. 2015, 46, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Qiu, W.; Wu, M.; Liu, S.; Chen, B.; Pan, C.; Yang, M.; Wang, K.-J. Suppressive immunoregulatory effects of three antidepressants via inhibition of the nuclear factor-κB activation assessed using primary macrophages of carp (Cyprinus carpio). Toxicol. Appl. Pharmacol. 2017, 322, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Munzer, A.; Sack, U.; Mergl, R.; Schönherr, J.; Petersein, C.; Bartsch, S.; Kirkby, K.C.; Bauer, K.; Himmerich, H. Impact of antidepressants on cytokine production of depressed patients in vitro. Toxins 2013, 5, 2227–2240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannestad, J.; DellaGioia, N.; Bloch, M. The effect of antidepressant medication treatment on serum levels of inflammatory cytokines: A meta-analysis. Neuropsychopharmacology 2011, 36, 2452–2459. [Google Scholar] [CrossRef] [PubMed]
- Więdłocha, M.; Marcinowicz, P.; Krupa, R.; Janoska-Jaździk, M.; Janus, M.; Dębowska, W.; Mosiołek, A.; Waszkiewicz, N.; Szulc, A. Effect of antidepressant treatment on peripheral inflammation markers—A meta-analysis. Prog. Neuropsychopharmacol. Biol. Psychiatr. 2018, 80, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.; Yeh, Y.-W.; Kuo, S.-C.; Liang, C.-S.; Ho, P.-S.; Huang, C.-C.; Yen, C.-H.; Shyu, J.-F.; Lu, R.-B.; Huang, S.-Y. Differences in immunomodulatory properties between venlafaxine and paroxetine in patients with major depressive disorder. Psychoneuroendocrinology 2018, 87, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Mohr, D.C.; Goodkin, D.E.; Islar, J.; Hauser, S.L.; Genain, C.P. Treatment of depression is associated with suppression of nonspecific and antigen-specific TH1 responses in multiple sclerosis. Arch. Neurol. 2001, 58, 1081–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunoni, A.R.; Machado-Vieira, R.; Zarate, C.A.; Valiengo, L.; Vieira, E.L.; Benseñor, I.M.; Lotufo, P.A.; Gattaz, W.F.; Teixeira, A.L. Cytokines plasma levels during antidepressant treatment with sertraline and transcranial direct current stimulation (tDCS): Results from a factorial, randomized, controlled trial. Psychopharmacology 2014, 231, 1315–1323. [Google Scholar] [CrossRef] [Green Version]
- Rethorst, C.D.; Toups, M.S.; Greer, T.L.; Nakonezny, P.A.; Carmody, T.J.; Grannemann, B.D.; Huebinger, R.M.; Barber, R.C.; Trivedi, M.H. Pro-inflammatory cytokines as predictors of antidepressant effects of exercise in major depressive disorder. Mol. Psychiatr. 2013, 18, 1119–1124. [Google Scholar] [CrossRef]
- Yrondi, A.; Sporer, M.; Peran, P.; Schmitt, L.; Arbus, C.; Sauvaget, A. Electroconvulsive therapy, depression, the immune system and inflammation: A systematic review. Brain Stimul. 2018, 11, 29–51. [Google Scholar] [CrossRef]
- Freire, T.F.V.; da Rocha, N.S.; de Almeida Fleck, M.P. The association of electroconvulsive therapy to pharmacological treatment and its influence on cytokines. J. Psychiatr. Res. 2017, 92, 205–211. [Google Scholar] [CrossRef]
- Kronfol, Z.; Nair, M.P.; Weinberg, V.; Young, E.A.; Aziz, M. Acute effects of electroconvulsive therapy on lymphocyte natural killer cell activity in patients with major depression. J. Affect Disord. 2002, 71, 211–215. [Google Scholar] [CrossRef]
- Kiraly, D.; Horn, S.; Van Dam, N.; Costi, S.; Schwartz, J.; Kim-Schulze, S.; Patel, M.; Hodes, G.E.; Russo, S.; Merad, M. Altered peripheral immune profiles in treatment-resistant depression: Response to ketamine and prediction of treatment outcome. Translational Psychiatr. 2017, 7, e1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myung, W.; Lim, S.-W.; Woo, H.I.; Park, J.H.; Shim, S.; Lee, S.-Y.; Kim, D.K. Serum cytokine levels in major depressive disorder and its role in antidepressant response. Psychiatr. Investig. 2016, 13, 644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uher, R.; Tansey, K.E.; Dew, T.; Maier, W.; Mors, O.; Hauser, J.; Dernovsek, M.Z.; Henigsberg, N.; Souery, D.; Farmer, A. An inflammatory biomarker as a differential predictor of outcome of depression treatment with escitalopram and nortriptyline. Am. J. Psychiatr. 2014, 171, 1278–1286. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, R.; Hori, H.; Ikenouchi-Sugita, A.; Umene-Nakano, W.; Ueda, N.; Nakamura, J. Higher plasma interleukin-6 (IL-6) level is associated with SSRI-or SNRI-refractory depression. Prog. Neuropsychopharmacol. Biol. Psychiatr. 2009, 33, 722–726. [Google Scholar] [CrossRef]
- Eller, T.; Vasar, V.; Shlik, J.; Maron, E. Pro-inflammatory cytokines and treatment response to escitaloprsam in major depressive disorder. Prog. Neuropsychopharmacol. Biol. Psychiatr. 2008, 32, 445–450. [Google Scholar] [CrossRef]
- Benedetti, F.; Lucca, A.; Brambilla, F.; Colombo, C.; Smeraldi, E. Interleukine-6 serum levels correlate with response to antidepressant sleep deprivation and sleep phase advance. Prog. Neuropsychopharmacol. Biol. Psychiatr. 2002, 26, 1167–1170. [Google Scholar] [CrossRef]
- Sorri, A.; Järventausta, K.; Kampman, O.; Lehtimäki, K.; Björkqvist, M.; Tuohimaa, K.; Hämäläinen, M.; Moilanen, E.; Leinonen, E. Low tumor necrosis factor-α levels predict symptom reduction during electroconvulsive therapy in major depressive disorder. Brain Behav. 2018, 8, e00933. [Google Scholar] [CrossRef]
- Yang, T.-T.; Wang, L.; Deng, X.-Y.; Yu, G. Pharmacological treatments for fatigue in patients with multiple sclerosis: A systematic review and meta-analysis. J. Neurol. Sci. 2017, 380, 256–261. [Google Scholar] [CrossRef]
- Rogóż, Z.; Kubera, M.; Rogóż, K.; Basta-Kaim, A.; Budziszewska, B. Effect of co-administration of fluoxetine and amantadine on immunoendocrine parameters in rats subjected to a forced swimming test. Pharmacol. Rep. 2009, 61, 1050–1060. [Google Scholar] [CrossRef]
- Lin, P.-Y.; Huang, S.-Y.; Su, K.-P. A meta-analytic review of polyunsaturated fatty acid compositions in patients with depression. Biol. Psychiatr. 2010, 68, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Appleton, K.M.; Voyias, P.D.; Sallis, H.M.; Dawson, S.; Ness, A.R.; Churchill, R.; Perry, R. Omega-3 fatty acids for depression in adults. Cochrane Database Syst. Rev. 2021, 11, CD004692. [Google Scholar] [PubMed]
- Hallahan, B.; Ryan, T.; Hibbeln, J.R.; Murray, I.T.; Glynn, S.; Ramsden, C.E.; SanGiovanni, J.P.; Davis, J.M. Efficacy of omega-3 highly unsaturated fatty acids in the treatment of depression. Br. J. Psychiatr. 2016, 209, 192–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.-D.; Feng, J.-S.; Yang, Z.; Huang, Q.-T.; Lin, J.-D.; Yang, B.; Su, K.-P.; Pan, J.-Y. High-dose omega-3 polyunsaturated fatty acid supplementation might be more superior than low-dose for major depressive disorder in early therapy period: A network meta-analysis. BMC Psychiatr. 2020, 20, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Ren, H.; Huang, Z.; Peng, Y.; He, B.; Yao, X.; Yuan, T.-F.; Su, H. Fish oil prevents lipopolysaccharide-induced depressive-like behavior by inhibiting neuroinflammation. Mol. Neurobiol. 2017, 54, 7327–7334. [Google Scholar] [CrossRef] [PubMed]
- Su, K.-P.; Lai, H.-C.; Yang, H.-T.; Su, W.-P.; Peng, C.-Y.; Chang, J.P.-C.; Chang, H.-C.; Pariante, C.M. Omega-3 fatty acids in the prevention of interferon-alpha-induced depression: Results from a randomized, controlled trial. Biol. Psychiatr. 2014, 76, 559–566. [Google Scholar] [CrossRef]
- Schuch, F.B.; Vancampfort, D.; Richards, J.; Rosenbaum, S.; Ward, P.B.; Stubbs, B. Exercise as a treatment for depression: A meta-analysis adjusting for publication bias. J. Psychiatr. Res. 2016, 77, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Morres, I.D.; Hatzigeorgiadis, A.; Stathi, A.; Comoutos, N.; Arpin-Cribbie, C.; Krommidas, C.; Theodorakis, Y. Aerobic exercise for adult patients with major depressive disorder in mental health services: A systematic review and meta-analysis. Depress. Anxiety 2019, 36, 39–53. [Google Scholar] [CrossRef] [Green Version]
- Wipfli, B.M.; Rethorst, C.D.; Landers, D.M. The anxiolytic effects of exercise: A meta-analysis of randomized trials and dose–response analysis. J. Sport Exerc. Psychol. 2008, 30, 392–410. [Google Scholar] [CrossRef] [Green Version]
- Phillips, C.; Fahimi, A. Immune and neuroprotective effects of physical activity on the brain in depression. Front. Neurosci. 2018, 12, 498. [Google Scholar] [CrossRef] [Green Version]
- Carrea-Gonzalez, M.D.P.; Canton-Habas, V.; Rich-Ruiz, M. Age, depression and dementia: The inflammatory process. Adv. Clin. Exp. Med. 2022, 31, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Chen, C. Recent advances in the study of the comorbidity of depressive and anxiety disorders. Adv. Clin. Exp. Med. 2022, 31, 355–358. [Google Scholar] [CrossRef]
- Tanaka, M.; Spekker, E.; Szabó, Á.; Polyák, H.; Vécsei, L. Modelling the neurodevelopmental pathogenesis in neuropsychiatric disorders. Bioactive kynurenines and their analogues as neuroprotective agents-in celebration of 80th birthday of Professor Peter Riederer. J. Neural Transm. 2022, 129, 627–642. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Török, N.; Tóth, F.; Szabó, Á.; Vécsei, L. Co-Players in Chronic Pain: Neuroinflammation and the Tryptophan-Kynurenine Metabolic Pathway. Biomedicines 2021, 9, 897. [Google Scholar] [CrossRef] [PubMed]
- Hunt, C.; Macedo ECordeiro, T.; Suchting, R.; de Dios, C.; Cuellar Leal, V.A.; Soares, J.C.; Dantzer, R.; Teixeira, A.L.; Selvaraj, S. Effect of immune activation on the kynurenine pathway and depression symptoms—A systematic review and meta-analysis. Neurosci Biobehav. Rev. 2020, 118, 514–523. [Google Scholar] [CrossRef]
- Sanjay Park, M.; Lee, H.J. Roles of Fatty Acids in Microglial Polarization: Evidence from In Vitro and In Vivo Studies on Neurodegenerative Diseases. Int. J. Mol. Sci. 2022, 23, 7300. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, I.-B.; Lee, J.-H.; Park, S.-C. The Relationship between Stress, Inflammation, and Depression. Biomedicines 2022, 10, 1929. https://doi.org/10.3390/biomedicines10081929
Kim I-B, Lee J-H, Park S-C. The Relationship between Stress, Inflammation, and Depression. Biomedicines. 2022; 10(8):1929. https://doi.org/10.3390/biomedicines10081929
Chicago/Turabian StyleKim, Il-Bin, Jae-Hon Lee, and Seon-Cheol Park. 2022. "The Relationship between Stress, Inflammation, and Depression" Biomedicines 10, no. 8: 1929. https://doi.org/10.3390/biomedicines10081929
APA StyleKim, I.-B., Lee, J.-H., & Park, S.-C. (2022). The Relationship between Stress, Inflammation, and Depression. Biomedicines, 10(8), 1929. https://doi.org/10.3390/biomedicines10081929