
Circulating Microvesicles in Association with the NLRP3 Inflammasome in Coronary Thrombi from STEMI Patients
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Sample Preparation
2.2. Laboratory Analyses
2.3. Statistical Analyses
3. Results
3.1. Clinical Characteristics
3.2. Circulating Microvesicles
3.3. Correlations between Circulating Microvesicles and Inflammasome Gene Expression in Thrombi
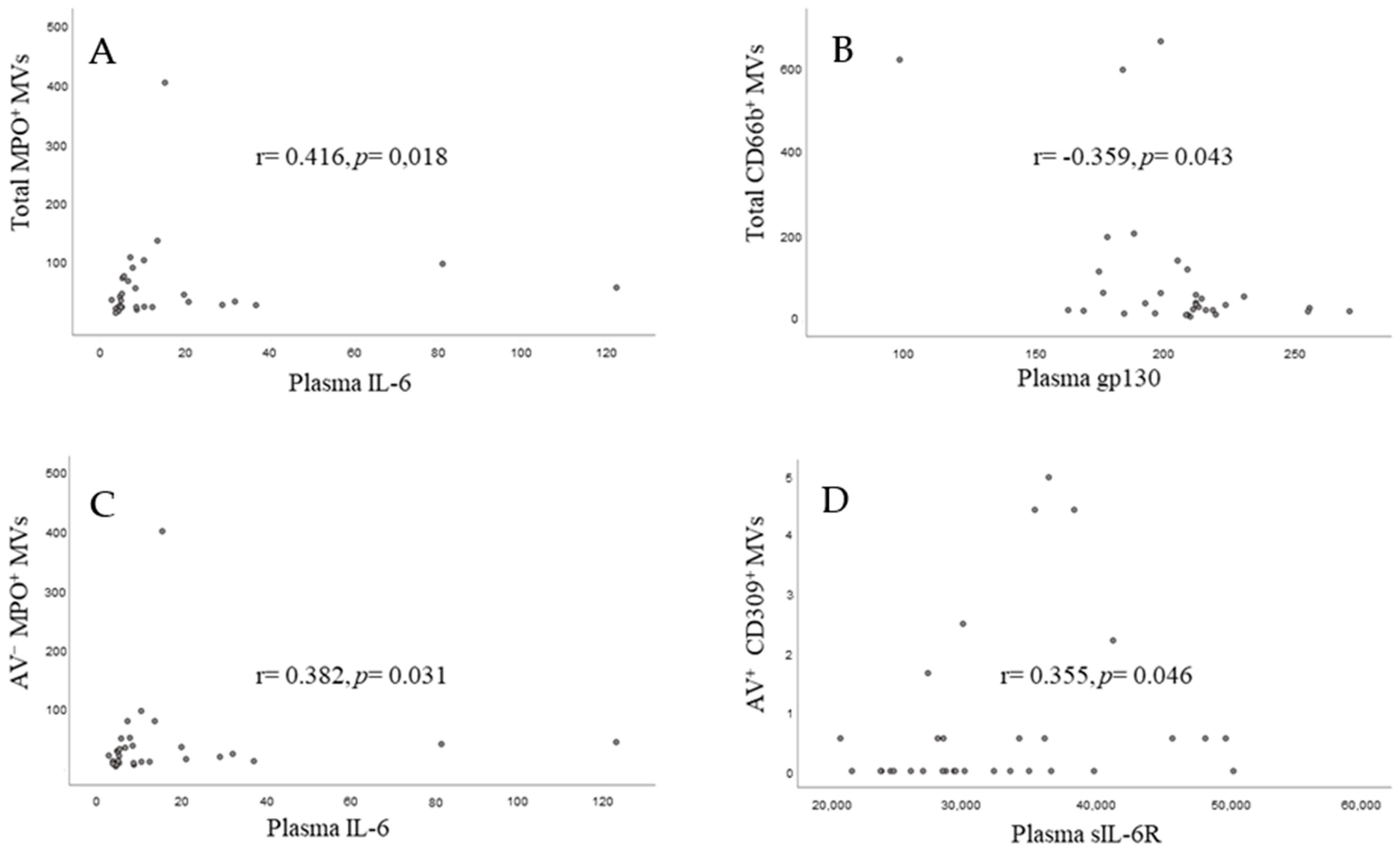
3.4. Correlations between Circulating Microvesicles and Corresponding Circulating Markers
3.5. PS Externalization on the MVs and the NLRP3 Inflammasome
3.6. MVs and Clinical Characteristics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Badimon, L.; Suades, R.; Vilella-Figuerola, A.; Crespo, J.; Vilahur, G.; Escate, R.; Padro, T.; Chiva-Blanch, G. Liquid Biopsies: Microvesicles in Cardiovascular Disease. Antioxid Redox Signal 2020, 33, 645–662. [Google Scholar] [CrossRef] [PubMed]
- Boulanger, C.M.; Loyer, X.; Rautou, P.-E.; Amabile, N. Extracellular vesicles in coronary artery disease. Nat. Rev. Cardiol. 2017, 14, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Meldolesi, J. Exosomes and Ectosomes in Intercellular Communication. Curr. Biol. 2018, 28, R435–R444. [Google Scholar] [CrossRef] [PubMed]
- Franco, A.T.; Corken, A.; Ware, J. Platelets at the interface of thrombosis, inflammation, and cancer. Blood 2015, 126, 582–588. [Google Scholar] [CrossRef]
- Wolf, P. The nature and significance of platelet products in human plasma. Br. J. Haematol. 1967, 13, 269–288. [Google Scholar] [CrossRef]
- Ridger, V.C.; Boulanger, C.M.; Angelillo-Scherrer, A.; Badimon, L.; Blanc-Brude, O.; Bochaton-Piallat, M.L.; Boilard, E.; Buzas, E.I.; Caporali, A.; Dignat-George, F.; et al. Microvesicles in vascular homeostasis and diseases. Position Paper of the European Society of Cardiology (ESC) Working Group on Atherosclerosis and Vascular Biology. Thromb. Haemost. 2017, 117, 1296–1316. [Google Scholar] [CrossRef]
- Arraud, N.; Linares, R.; Tan, S.; Gounou, C.; Pasquet, J.M.; Mornet, S.; Brisson, A.R. Extracellular vesicles from blood plasma: Determination of their morphology, size, phenotype and concentration. J. Thromb. Haemost. 2014, 12, 614–627. [Google Scholar] [CrossRef]
- Boilard, E.; Duchez, A.-C.; Brisson, A. The diversity of platelet microparticles. Curr. Opin. Hematol. 2015, 22, 437–444. [Google Scholar] [CrossRef]
- Connor, D.E.; Exner, T.; Ma, D.D.; Joseph, J.E. The majority of circulating platelet-derived microparticles fail to bind annexin V, lack phospholipid-dependent procoagulant activity and demonstrate greater expression of glycoprotein Ib. Thromb. Haemost. 2010, 103, 1044–1052. [Google Scholar] [CrossRef]
- Hansson, G.K. Inflammation, atherosclerosis, and coronary artery disease. N. Engl. J. Med. 2005, 352, 1685–1695. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Cui, J.; Zhang, G.; Wu, C.; Abdel-Latif, A.; Smyth, S.S.; Shiroishi, T.; Mackman, N.; Wei, Y.; Tao, M.; et al. Inflammasome activation promotes venous thrombosis through pyroptosis. Blood Adv. 2021, 5, 2619–2623. [Google Scholar] [CrossRef] [PubMed]
- Grebe, A.; Hoss, F.; Latz, E. NLRP3 Inflammasome and the IL-1 Pathway in Atherosclerosis. Circ. Res. 2018, 122, 1722–1740. [Google Scholar] [CrossRef] [PubMed]
- Paramel Varghese, G.; Folkersen, L.; Strawbridge, R.J.; Halvorsen, B.; Yndestad, A.; Ranheim, T.; Krohg-Sørensen, K.; Skjelland, M.; Espevik, T.; Aukrust, P.; et al. NLRP3 Inflammasome Expression and Activation in Human Atherosclerosis. J. Am. Heart Assoc. 2016, 5, e003031. [Google Scholar] [CrossRef]
- Curtiss, L.K.; Tobias, P.S. Emerging role of Toll-like receptors in atherosclerosis. J. Lipid Res. 2009, 50, S340–S345. [Google Scholar] [CrossRef]
- Broz, P.; Dixit, V.M. Inflammasomes: Mechanism of assembly, regulation and signalling. Nat. Rev. Immunol. 2016, 16, 407–420. [Google Scholar] [CrossRef]
- Wu, C.; Lu, W.; Zhang, Y.; Zhang, G.; Shi, X.; Hisada, Y.; Grover, S.P.; Zhang, X.; Li, L.; Xiang, B.; et al. Inflammasome Activation Triggers Blood Clotting and Host Death through Pyroptosis. Immunity 2019, 50, 1401–1411.e1404. [Google Scholar] [CrossRef] [PubMed]
- Chanchal, S.; Mishra, A.; Singh, M.K.; Ashraf, M.Z. Understanding Inflammatory Responses in the Manifestation of Prothrombotic Phenotypes. Front. Cell Dev. Biol. 2020, 8, 73. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.H.; Conway, E.M. Cross Talk Pathways Between Coagulation and Inflammation. Circ. Res. 2016, 118, 1392–1408. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Masuda, K.; Kishimoto, T. Regulation of IL-6 in Immunity and Diseases. Adv. Exp. Med. Biol. 2016, 941, 79–88. [Google Scholar] [CrossRef]
- Wolf, J.; Rose-John, S.; Garbers, C. Interleukin-6 and its receptors: A highly regulated and dynamic system. Cytokine 2014, 70, 11–20. [Google Scholar] [CrossRef]
- Ibanez, B.; James, S.; Agewall, S.; Antunes, M.J.; Bucciarelli-Ducci, C.; Bueno, H.; Caforio, A.L.P.; Crea, F.; Goudevenos, J.A.; Halvorsen, S.; et al. 2017 ESC Guidelines for the management of acute myocardial infarction in patients presenting with ST-segment elevation: The Task Force for the management of acute myocardial infarction in patients presenting with ST-segment elevation of the European Society of Cardiology (ESC). Eur. Heart J. 2018, 39, 119–177. [Google Scholar] [CrossRef] [PubMed]
- Nordeng, J.; Schandiz, H.; Solheim, S.; Åkra, S.; Hoffman, P.; Roald, B.; Bendz, B.; Arnesen, H.; Helseth, R.; Seljeflot, I. The Inflammasome Signaling Pathway Is Actively Regulated and Related to Myocardial Damage in Coronary Thrombi from Patients with STEMI. Mediat. Inflamm. 2021, 2021, 5525917. [Google Scholar] [CrossRef]
- Cypryk, W.; Nyman, T.A.; Matikainen, S. From Inflammasome to Exosome-Does Extracellular Vesicle Secretion Constitute an Inflammasome-Dependent Immune Response? Front. Immunol. 2018, 9, 2188. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, F.; Yuan, Y.; Jin, C.; Chang, C.; Zhu, Y.; Zhang, X.; Tian, C.; He, F.; Wang, J. Inflammasome-Derived Exosomes Activate NF-κB Signaling in Macrophages. J. Proteome Res. 2017, 16, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Nieuwland, R.; Berckmans, R.J.; McGregor, S.; Boing, A.N.; Romijn, F.P.; Westendorp, R.G.; Hack, C.E.; Sturk, A. Cellular origin and procoagulant properties of microparticles in meningococcal sepsis. Blood 2000, 95, 930–935. [Google Scholar] [CrossRef]
- Gedefaw, L.; Ullah, S.; Leung, P.H.M.; Cai, Y.; Yip, S.P.; Huang, C.L. Inflammasome Activation-Induced Hypercoagulopathy: Impact on Cardiovascular Dysfunction Triggered in COVID-19 Patients. Cells 2021, 10, 916. [Google Scholar] [CrossRef]
- Rothmeier, A.S.; Marchese, P.; Petrich, B.G.; Furlan-Freguia, C.; Ginsberg, M.H.; Ruggeri, Z.M.; Ruf, W. Caspase-1-mediated pathway promotes generation of thromboinflammatory microparticles. J. Clin. Invest. 2015, 125, 1471–1484. [Google Scholar] [CrossRef]
- Budden, C.F.; Gearing, L.J.; Kaiser, R.; Standke, L.; Hertzog, P.J.; Latz, E. Inflammasome-induced extracellular vesicles harbour distinct RNA signatures and alter bystander macrophage responses. J. Extracell Vesicles 2021, 10, e12127. [Google Scholar] [CrossRef]
- Döring, Y.; Drechsler, M.; Soehnlein, O.; Weber, C. Neutrophils in atherosclerosis: From mice to man. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 288–295. [Google Scholar] [CrossRef] [Green Version]
- Warnatsch, A.; Ioannou, M.; Wang, Q.; Papayannopoulos, V. Inflammation. Neutrophil extracellular traps license macrophages for cytokine production in atherosclerosis. Science 2015, 349, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Martínez, G.J.; Barraclough, J.Y.; Nakhla, S.; Kienzle, V.; Robertson, S.; Mallat, Z.; Celermajer, D.S.; Patel, S. Neutrophil-derived microparticles are released into the coronary circulation following percutaneous coronary intervention in acute coronary syndrome patients. Biosci. Rep. 2017, 37, BSR20160430. [Google Scholar] [CrossRef]
- Sionis, A.; Suades, R.; Sans-Roselló, J.; Sánchez-Martínez, M.; Crespo, J.; Padró, T.; Cubedo, J.; Ferrero-Gregori, A.; Vila-Perales, M.; Duran-Cambra, A.; et al. Circulating microparticles are associated with clinical severity of persistent ST-segment elevation myocardial infarction complicated with cardiogenic shock. Int. J. Cardiol. 2018, 258, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Chiva-Blanch, G.; Bratseth, V.; Ritschel, V.; Andersen, G.O.; Halvorsen, S.; Eritsland, J.; Arnesen, H.; Badimon, L.; Seljeflot, I. Monocyte-derived circulating microparticles (CD14(+), CD14(+)/CD11b(+) and CD14(+)/CD142(+)) are related to long-term prognosis for cardiovascular mortality in STEMI patients. Int. J. Cardiol. 2017, 227, 876–881. [Google Scholar] [CrossRef]
- Mavroudis, C.A.; Eleftheriou, D.; Hong, Y.; Majumder, B.; Koganti, S.; Sapsford, R.; North, J.; Lowdell, M.; Klein, N.; Brogan, P.; et al. Microparticles in acute coronary syndrome. Thromb. Res. 2017, 156, 109–116. [Google Scholar] [CrossRef]
- Haller, P.M.; Jäger, B.; Piackova, E.; Sztulman, L.; Wegberger, C.; Wojta, J.; Gyöngyösi, M.; Kiss, A.; Podesser, B.K.; Spittler, A.; et al. Changes in Circulating Extracellular Vesicles in Patients with ST-Elevation Myocardial Infarction and Potential Effects of Remote Ischemic Conditioning-A Randomized Controlled Trial. Biomedicines 2020, 8, 218. [Google Scholar] [CrossRef]
- Suades, R.; Padró, T.; Crespo, J.; Ramaiola, I.; Martin-Yuste, V.; Sabaté, M.; Sans-Roselló, J.; Sionis, A.; Badimon, L. Circulating microparticle signature in coronary and peripheral blood of ST elevation myocardial infarction patients in relation to pain-to-PCI elapsed time. Int. J. Cardiol. 2016, 202, 378–387. [Google Scholar] [CrossRef]
- Lacroix, R.; Robert, S.; Poncelet, P.; Dignat-George, F. Overcoming limitations of microparticle measurement by flow cytometry. Semin. Thromb. Hemost. 2010, 36, 807–818. [Google Scholar] [CrossRef]
- Bergen, K.; Mobarrez, F.; Jorneskog, G.; Wallen, H.; Tehrani, S. Phosphatidylserine expressing microvesicles in relation to microvascular complications in type 1 diabetes. Thromb. Res. 2018, 172, 158–164. [Google Scholar] [CrossRef]
- Lundström, A.; Mobarrez, F.; Rooth, E.; Thålin, C.; von Arbin, M.; Henriksson, P.; Gigante, B.; Laska, A.-C.; Wallén, H. Prognostic Value of Circulating Microvesicle Subpopulations in Ischemic Stroke and TIA. Transl. Stroke Res. 2020, 11, 708–719. [Google Scholar] [CrossRef] [Green Version]
- Mobarrez, F.; Gunnarsson, I.; Svenungsson, E. Altered β(2) -glycoprotein I expression on microparticles in the presence of antiphospholipid antibodies. J. Thromb. Haemost. 2017, 15, 1799–1806. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Shi, Y.; Waehrens, L.N.; Rasmussen, J.T.; Heegaard, C.W.; Gilbert, G.E. Lactadherin detects early phosphatidylserine exposure on immortalized leukemia cells undergoing programmed cell death. Cytom. A 2006, 69, 1193–1201. [Google Scholar] [CrossRef] [PubMed]
- Mobarrez, F.; Vikerfors, A.; Gustafsson, J.T.; Gunnarsson, I.; Zickert, A.; Larsson, A.; Pisetsky, D.S.; Wallén, H.; Svenungsson, E. Microparticles in the blood of patients with systemic lupus erythematosus (SLE): Phenotypic characterization and clinical associations. Sci. Rep. 2016, 6, 36025. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Baseline Characteristics | |
|---|---|
| Age, years (mean (range)) | 60 (38, 83) |
| Females | 3 (9%) |
| BMI (kg/m2) | 27.7 (23.4, 28.6) |
| Hypertension | 11 (33%) |
| T2DM | 4 (12%) |
| Previous CAD | 1 (3%) |
| Current smokers | 16 (49%) |
| Medication: | |
| ASA | 6 (18%) |
| Plavix | 2 (6%) |
| Anticoagulation | 3 (9%) |
| NOAC | 2 (6%) |
| Betablocker | 4 (12%) |
| AT-II-blocker | 5 (15%) |
| Statins | 6 (18%) |
| Diuretics | 4 (12%) |
| SBP a (mmHg) | 127 (±31) |
| DBP a (mmHg) | 82 (±19) |
| Ischaemic time (min) | 152 (121, 375) |
| cTnT after PCI (µg/L) | 354 (119, 748) |
| cTnT peak (µg/L) | 3434 (1179, 7652) |
| CRP (mg/L) | 2.71 (0.96, 6.03) |
| Culprit lesion: | |
| LAD | 16 (49%) |
| CX | 6 (18%) |
| RCA | 11 (33%) |
| Retrograde flow | 12 (36%) |
| Three-vessel disease | 6 (18%) |
| In-stent thrombus | 1 (3%) |
| Total MVs/µL PPP | AV−/µL PPP | AV+/µL PPP | |
|---|---|---|---|
| Total | 5112.83 (2807.52, 7139.93) | 4917.04 (2660.95, 7112.14) | 51.16 (28.07, 92.37) |
| TF+ | 14.66 (6.91, 26.41) | 0.55 (0.00, 1.52) | 11.34 (5.67, 25.72) |
| Platelet | |||
| CD61 | 125.41 (59.60, 250.83) | 79.37 (33.88, 138.69) | 25.17 (10.92, 71.21) |
| CD62P | 23.51 (15.49, 38.16) | 21.02 (14.38, 34.02) | 1.11 (0.55, 2.77) |
| CD42b | 28.76 (15.49, 63.33) | 18.53 (10.51, 35.40) | 6.36 (1.24, 18.53) |
| Endothelial | |||
| CD309 | 5.53 (2.90, 9.40) | 4.98 (2.90, 9.26 | 0.00 (0.00, 0.55) |
| CD62E | 27.10 (19.50, 36.64) | 24.34 (17.98, 32.91) | 0.00 (0.00, 0.55) |
| CD31 | 35.40 (27.79, 68.45) | 29.31 (21.43, 46.18) | 5.81 (2.35, 11.75) |
| Leukocytes | |||
| CD14 | 8.57 (4.98, 16.04) | 8.30 (4.98, 14.66) | 0.00 (0.00, 0.55) |
| CD11b | 64.99 (43.32, 100.39) | 63.33 (45.08, 97.83) | 0.55 (0.55, 1.11) |
| CD62L | 2.49 (0.69, 5.91) | 2.21 (0.69, 4.84) | 0.00 (0.00, 0.55) |
| CD66b | 36.50 (21.29, 102.46) | 15.49 (13.0, 34.15) | 10.79 (5.81, 43.83) |
| MPO | 30.14 (19.91, 67.89) | 21.85 (9.96, 40.79) | 9.40 (4.42, 19.91) |
| CD45 | 15.49 (11.06, 27.65) | 13.27 (9.96, 26.69) | 0.55 (0.14, 1.66) |
| CD15 | 1.66 (0.55, 3.32) | 1.11 (0.55, 3.32) | 0.00 (0.00, 0.41) |
| NLRP3 | CASPASE-1 | IL-1 β | IL-18 | IL-6 | sIL-6R | gp130 | ||
|---|---|---|---|---|---|---|---|---|
| Total | r | −0.322 | −0.053 | −0.399 | −0.170 | −0.186 | −0.298 | −0.457 |
| p | 0.143 | 0.791 | 0.032 | 0.428 | 0.396 | 0.124 | 0.019 | |
| AV− | r | −0.362 | −0.062 | −0.438 | −0.190 | −0.129 | −0.340 | −0.502 |
| p | 0.098 | 0.752 | 0.017 | 0.373 | 0.558 | 0.076 | 0.009 | |
| AV+ | r | 0.545 | −1.06 | 0.114 | 0.108 | −0.186 | 0.113 | 0.003 |
| p | 0.009 | 0.592 | 0.557 | 0.615 | 0.395 | 0.568 | 0.988 |
| NLRP3 | IL-1β | Caspase-1 | IL-18 | IL-6 | sIL-6R | gp130 | ||
|---|---|---|---|---|---|---|---|---|
| Thrombi genes | r | 0.592 | 0.309 | −0.018 | 0.165 | −0.005 | 0.279 | 0.279 |
| p | 0.004 a | 0.103 | 0.927 | 0.440 | 0.980 | 0.151 | 0.151 | |
| Plasma | r | 0.073 | −0.139 | 0.034 | −0.034 | |||
| p | 0.690 | 0.449 | 0.855 | 0.854 |
| Ischaemic Time | Peak cTnT | ||
|---|---|---|---|
| Total MVs | r | −0.137 | 0.208 |
| p | 0.454 | 0.254 | |
| AV− MVs | r | −0.096 | 0.228 |
| p | 0.600 | 0.210 | |
| AV+ MVs | r | −0.139 | −0.007 |
| p | 0.448 | 0.969 |
| Total MVs (µL/PPP) | AV− MVs (µL/PPP) | AV+ MVs (µL/PPP) | ||
|---|---|---|---|---|
| Smoke a | No smoke (n = 16) | 5249.17 (2571.49, 7844.72) | 5144.91 (2525.17, 7776.41) | 45.77 (24.34, 82.96) |
| Current smoker (n = 15) | 5068.58 (2976.77, 5841.26) | 4122.23 (2712.39, 5310.84) | 51.99 (30.42, 95.13) | |
| p-value | 0.797 | 0.621 | 0.374 | |
| BMI b | <median (n = 13) | 4850.11 (2751.66, 6594.30) | 4822.46 (2600.39, 6564.44) | 43.69 (27.10, 68.86) |
| ≥median (n = 14) | 5414.55 (2742.95, 7370.44) | 5414.55 (2742.95, 7370.44) | 62.09 (25.44, 146.99) | |
| p-value | 0.577 | 0.680 | 0.452 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bratseth, V.; Nordeng, J.; Helseth, R.; Solheim, S.; Åkra, S.; Arnesen, H.; Chiva-Blanch, G.; Seljeflot, I. Circulating Microvesicles in Association with the NLRP3 Inflammasome in Coronary Thrombi from STEMI Patients. Biomedicines 2022, 10, 2196. https://doi.org/10.3390/biomedicines10092196
Bratseth V, Nordeng J, Helseth R, Solheim S, Åkra S, Arnesen H, Chiva-Blanch G, Seljeflot I. Circulating Microvesicles in Association with the NLRP3 Inflammasome in Coronary Thrombi from STEMI Patients. Biomedicines. 2022; 10(9):2196. https://doi.org/10.3390/biomedicines10092196
Chicago/Turabian StyleBratseth, Vibeke, Jostein Nordeng, Ragnhild Helseth, Svein Solheim, Sissel Åkra, Harald Arnesen, Gemma Chiva-Blanch, and Ingebjørg Seljeflot. 2022. "Circulating Microvesicles in Association with the NLRP3 Inflammasome in Coronary Thrombi from STEMI Patients" Biomedicines 10, no. 9: 2196. https://doi.org/10.3390/biomedicines10092196
APA StyleBratseth, V., Nordeng, J., Helseth, R., Solheim, S., Åkra, S., Arnesen, H., Chiva-Blanch, G., & Seljeflot, I. (2022). Circulating Microvesicles in Association with the NLRP3 Inflammasome in Coronary Thrombi from STEMI Patients. Biomedicines, 10(9), 2196. https://doi.org/10.3390/biomedicines10092196