
Tephrosin Suppresses the Chemoresistance of Paclitaxel-Resistant Ovarian Cancer via Inhibition of FGFR1 Signaling Pathway
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Culture
2.2. Cell Viability Using the Water-Soluble Tetrazolium 1 (WST-1) Assay and Crystal Violet Assay
2.3. Cell Cytotoxicity Analysis Using the Lactate Dehydrogenase (LDH) Assay
2.4. Reverse Transcription Polymerase Chain Reaction (RT-PCR) and Reverse Transcription Quantitative Polymerase Chain Reaction (RT-qPCR) Tests
2.5. Immunoblotting
2.6. Fluorescence-Activated Cell Sorting (FACS) Analysis
2.7. Fluo-3 Acetoxymethyl (AM) Assay
2.8. Statistical Analysis
3. Results
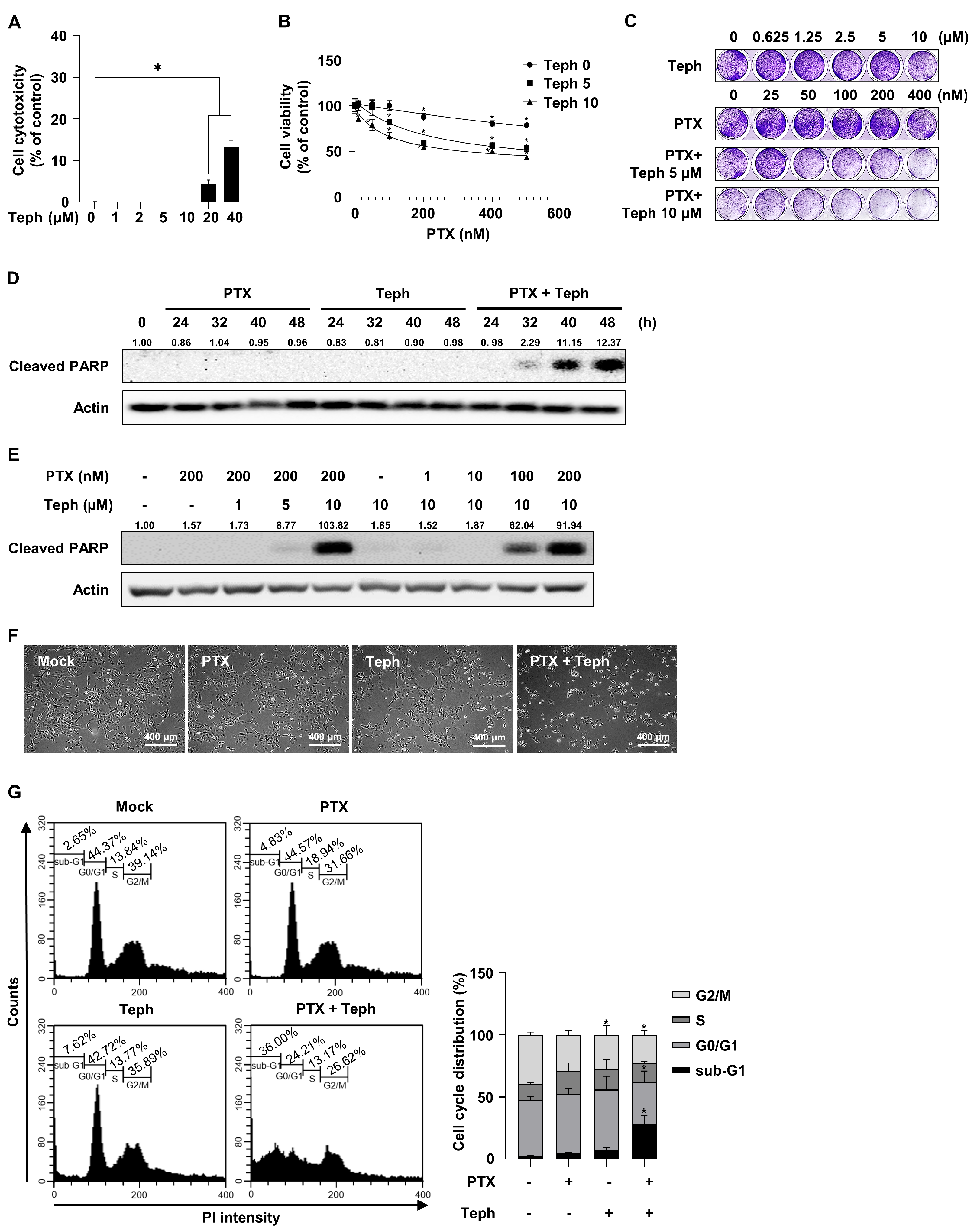
3.1. Combination Treatment with Paclitaxel and Tephrosin Effectively Induced Cytotoxicity in SKOV3-TR Cells
3.2. Tephrosin Inhibited the Phosphorylation of AKT, STAT3, ERK, and p38 MAPK in SKOV3-TR Cells
3.3. Tephrosin Downregulated the Phosphorylation of FGFR1/FRS2 Signaling Pathway in SKOV3-TR Cells
3.4. Tephrosin Suppressed Paclitaxel Resistance Independently of P-Glycoprotein (P-gp) Expression and Function in SKOV3-TR Cells
3.5. Tephrosin Inhibited the Expression of XIAP in SKOV3-TR Cells
3.6. Tephrosin Also Restored Paclitaxel Resistance in Other Paclitaxel-Resistant Ovarian HeyA8-MDR Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sambasivan, S. Epithelial ovarian cancer: Review article. Cancer Treat. Res. Commun. 2022, 33, 100629. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.R.; Shih, I.M. Ovarian cancer. Annu. Rev. Pathol. 2009, 4, 287–313. [Google Scholar] [CrossRef] [PubMed]
- Doubeni, C.A.; Doubeni, A.R.; Myers, A.E. Diagnosis and Management of Ovarian Cancer. Am. Fam. Physician 2016, 93, 937–944. [Google Scholar] [PubMed]
- Jayson, G.C.; Kohn, E.C.; Kitchener, H.C.; Ledermann, J.A. Ovarian cancer. Lancet 2014, 384, 1376–1388. [Google Scholar] [CrossRef] [PubMed]
- Bookman, M.A. Optimal primary therapy of ovarian cancer. Ann. Oncol. 2016, 27 (Suppl. 1), i58–i62. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Ueda, Y.; Naka, T.; Enomoto, T. Therapeutic strategies in epithelial ovarian cancer. J. Exp. Clin. Cancer Res. 2012, 31, 14. [Google Scholar] [CrossRef] [PubMed]
- Kampan, N.C.; Madondo, M.T.; McNally, O.M.; Quinn, M.; Plebanski, M. Paclitaxel and Its Evolving Role in the Management of Ovarian Cancer. BioMed Res. Int. 2015, 2015, 413076. [Google Scholar] [CrossRef]
- Ortiz, M.; Wabel, E.; Mitchell, K.; Horibata, S. Mechanisms of chemotherapy resistance in ovarian cancer. Cancer Drug Resist. 2022, 5, 304–316. [Google Scholar] [CrossRef]
- Das, T.; Anand, U.; Pandey, S.K.; Ashby, C.R., Jr.; Assaraf, Y.G.; Chen, Z.S.; Dey, A. Therapeutic strategies to overcome taxane resistance in cancer. Drug Resist. Updates 2021, 55, 100754. [Google Scholar] [CrossRef]
- Weaver, B.A. How Taxol/paclitaxel kills cancer cells. Mol. Biol. Cell 2014, 25, 2677–2681. [Google Scholar] [CrossRef]
- Kannampuzha, S.; Gopalakrishnan, A.V. Cancer chemoresistance and its mechanisms: Associated molecular factors and its regulatory role. Med. Oncol. 2023, 40, 264. [Google Scholar] [CrossRef] [PubMed]
- Adil, M.; Kanwal, S.; Rasheed, S.; Iqbal, M.; Abbas, G. Cancer Chemoresistance; Recent Challenges and Future Considerations. Cancer Treat. Res. 2023, 185, 237–253. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, S.; Coward, J.I.; Bast, R.C., Jr.; Berchuck, A.; Berek, J.S.; Brenton, J.D.; Coukos, G.; Crum, C.C.; Drapkin, R.; Etemadmoghadam, D.; et al. Rethinking ovarian cancer: Recommendations for improving outcomes. Nat. Rev. Cancer 2011, 11, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Freimund, A.E.; Beach, J.A.; Christie, E.L.; Bowtell, D.D.L. Mechanisms of Drug Resistance in High-Grade Serous Ovarian Cancer. Hematol. Oncol. Clin. N. Am. 2018, 32, 983–996. [Google Scholar] [CrossRef] [PubMed]
- Mosca, L.; Ilari, A.; Fazi, F.; Assaraf, Y.G.; Colotti, G. Taxanes in cancer treatment: Activity, chemoresistance and its overcoming. Drug Resist. Updates 2021, 54, 100742. [Google Scholar] [CrossRef] [PubMed]
- Adamska, A.; Elaskalani, O.; Emmanouilidi, A.; Kim, M.; Abdol Razak, N.B.; Metharom, P.; Falasca, M. Molecular and cellular mechanisms of chemoresistance in pancreatic cancer. Adv. Biol. Regul. 2018, 68, 77–87. [Google Scholar] [CrossRef]
- Zhao, S.; Tang, Y.; Wang, R.; Najafi, M. Mechanisms of cancer cell death induction by paclitaxel: An updated review. Apoptosis 2022, 27, 647–667. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Juhnn, Y.S.; Song, Y.S. Akt involvement in paclitaxel chemoresistance of human ovarian cancer cells. Ann. N. Y. Acad. Sci. 2007, 1095, 82–89. [Google Scholar] [CrossRef]
- Fan, Z.; Cui, H.; Yu, H.; Ji, Q.; Kang, L.; Han, B.; Wang, J.; Dong, Q.; Li, Y.; Yan, Z.; et al. MiR-125a promotes paclitaxel sensitivity in cervical cancer through altering STAT3 expression. Oncogenesis 2016, 5, e223. [Google Scholar] [CrossRef]
- Zhang, R.; Yang, X.; Roque, D.M.; Li, C.; Lin, J. A novel small molecule LLL12B inhibits STAT3 signaling and sensitizes ovarian cancer cell to paclitaxel and cisplatin. PLoS ONE 2021, 16, e0240145. [Google Scholar] [CrossRef]
- Shih, P.C.; Mei, K.C. Role of STAT3 signaling transduction pathways in cancer stem cell-associated chemoresistance. Drug Discov. Today 2021, 26, 1450–1458. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zha, J.; Lei, M. Inhibiting ERK/Mnk/eIF4E broadly sensitizes ovarian cancer response to chemotherapy. Clin. Transl. Oncol. 2018, 20, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhao, R.; Qin, X.; Mao, X.; Li, Q.; Fang, S. Cobimetinib Sensitizes Cervical Cancer to Paclitaxel via Suppressing Paclitaxel-Induced ERK Activation. Pharmacology 2022, 107, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Kudaravalli, S.; den Hollander, P.; Mani, S.A. Role of p38 MAP kinase in cancer stem cells and metastasis. Oncogene 2022, 41, 3177–3185. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Xiao, L.; Li, Z. The relationship between p38MAPK and apoptosis during paclitaxel resistance of ovarian cancer cells. J. Huazhong Univ. Sci. Technol. Med. Sci. 2007, 27, 725–728. [Google Scholar] [CrossRef] [PubMed]
- Revathidevi, S.; Munirajan, A.K. Akt in cancer: Mediator and more. Semin. Cancer Biol. 2019, 59, 80–91. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Sun, M.M.; Zhang, G.G.; Yang, J.; Chen, K.S.; Xu, W.W.; Li, B. Targeting PI3K/Akt signal transduction for cancer therapy. Signal Transduct. Target. Ther. 2021, 6, 425. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Song, H.; Yang, P.; Dai, X.; Li, Y.; Wang, L.; Du, J.; Pan, K.; Zhang, T. Silencing dishevelled-1 sensitizes paclitaxel-resistant human ovarian cancer cells via AKT/GSK-3beta/beta-catenin signalling. Cell Prolif. 2015, 48, 249–258. [Google Scholar] [CrossRef]
- Yang, Y.I.; Lee, K.T.; Park, H.J.; Kim, T.J.; Choi, Y.S.; Shih Ie, M.; Choi, J.H. Tectorigenin sensitizes paclitaxel-resistant human ovarian cancer cells through downregulation of the Akt and NFkappaB pathway. Carcinogenesis 2012, 33, 2488–2498. [Google Scholar] [CrossRef]
- Ou, C.; Peng, C.; Wang, Y.; Lu, S.; Yu, X.; He, Q.; He, A.; Zhang, L. CRTC2 promotes paclitaxel resistance by inducing autophagy in ovarian cancer in part via the PI3K-AKT signaling axis. J. Cancer 2023, 14, 1011–1023. [Google Scholar] [CrossRef]
- Suyama, H.; Igishi, T.; Sano, H.; Matsumoto, S.; Shigeoka, Y.; Nakanishi, H.; Endo, M.; Burioka, N.; Hitsuda, Y.; Shimizu, E. ERK activation and subsequent RB phosphorylation are important determinants of the sensitivity to paclitaxel in lung adenocarcinoma cells. Int. J. Oncol. 2004, 24, 1499–1504. [Google Scholar]
- Wu, G.; Qin, X.Q.; Guo, J.J.; Li, T.Y.; Chen, J.H. AKT/ERK activation is associated with gastric cancer cell resistance to paclitaxel. Int. J. Clin. Exp. Pathol. 2014, 7, 1449–1458. [Google Scholar]
- Kawiak, A.; Domachowska, A.; Lojkowska, E. Plumbagin Increases Paclitaxel-Induced Cell Death and Overcomes Paclitaxel Resistance in Breast Cancer Cells through ERK-Mediated Apoptosis Induction. J. Nat. Prod. 2019, 82, 878–885. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Zhao, S.; Shi, Z.; Cao, L.; Liu, J.; Pan, T.; Zhou, D.; Zhang, J. Chemotherapy-elicited exosomal miR-378a-3p and miR-378d promote breast cancer stemness and chemoresistance via the activation of EZH2/STAT3 signaling. J. Exp. Clin. Cancer Res. 2021, 40, 120. [Google Scholar] [CrossRef] [PubMed]
- Park, S.A.; Kim, L.K.; Park, H.M.; Kim, H.J.; Heo, T.H. Inhibition of GP130/STAT3 and EMT by combined bazedoxifene and paclitaxel treatment in ovarian cancer. Oncol. Rep. 2022, 47, 52. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.L.; Chen, X.; Zhang, X.Y.; Li, Z.M.; Fan, X.M.; Shen, Y. Octreotide-Paclitaxel Conjugate Reverses Paclitaxel Resistance by p38 Mitogen-Activated Protein Kinase (MAPK) Signaling Pathway in A2780/Taxol Human Ovarian Cancer Cells. Med. Sci. Monit. 2020, 26, e922612. [Google Scholar] [CrossRef]
- Stieg, D.C.; Wang, Y.; Liu, L.Z.; Jiang, B.H. ROS and miRNA Dysregulation in Ovarian Cancer Development, Angiogenesis and Therapeutic Resistance. Int. J. Mol. Sci. 2022, 23, 6702. [Google Scholar] [CrossRef]
- Wu, J.; Yeung, S.J.; Liu, S.; Qdaisat, A.; Jiang, D.; Liu, W.; Cheng, Z.; Liu, W.; Wang, H.; Li, L.; et al. Cyst(e)ine in nutrition formulation promotes colon cancer growth and chemoresistance by activating mTORC1 and scavenging ROS. Signal Transduct. Target. Ther. 2021, 6, 188. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; An, B.; Lin, Y.; Ni, Y.; Zhao, X.; Liang, X. Molecular mechanisms of ROS-modulated cancer chemoresistance and therapeutic strategies. Biomed. Pharmacother. 2023, 165, 115036. [Google Scholar] [CrossRef]
- Li, J.; Liu, P.; Mao, H.; Wanga, A.; Zhang, X. Emodin sensitizes paclitaxel-resistant human ovarian cancer cells to paclitaxel-induced apoptosis in vitro. Oncol. Rep. 2009, 21, 1605–1610. [Google Scholar] [CrossRef]
- Sharifi, S.; Barar, J.; Hejazi, M.S.; Samadi, N. Roles of the Bcl-2/Bax ratio, caspase-8 and 9 in resistance of breast cancer cells to paclitaxel. Asian Pac. J. Cancer Prev. 2014, 15, 8617–8622. [Google Scholar] [CrossRef]
- Williams, J.; Lucas, P.C.; Griffith, K.A.; Choi, M.; Fogoros, S.; Hu, Y.Y.; Liu, J.R. Expression of Bcl-xL in ovarian carcinoma is associated with chemoresistance and recurrent disease. Gynecol. Oncol. 2005, 96, 287–295. [Google Scholar] [CrossRef]
- Vergara, D.; Tinelli, A.; Iannone, A.; Maffia, M. The impact of proteomics in the understanding of the molecular basis of Paclitaxel-resistance in ovarian tumors. Curr. Cancer Drug Targets 2012, 12, 987–997. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Gao, M.; Li, Z.; Song, L.; Gao, X.; Han, J.; Wang, F.; Chen, Y.; Li, W.; Yang, J. p53 and P-glycoprotein influence chemoresistance in hepatocellular carcinoma. Front. Biosci. (Elite Ed.) 2018, 10, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Z.; Mousavi, M.; Moliani, A.; Bahman, Y.; Bagheri, H. Resveratrol inhibits glioblastoma cells and chemoresistance progression through blockade P-glycoprotein and targeting AKT/PTEN signaling pathway. Chem. Biol. Interact. 2023, 376, 110409. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Yin, Z.; Tan, L.; Zhu, W.; Tao, K.; Wang, G.; Shi, W.; Gao, J. Interferon regulatory factor-1 reverses chemoresistance by downregulating the expression of P-glycoprotein in gastric cancer. Cancer Lett. 2019, 457, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Su, N.; Yang, J.; Tan, Q.; Huang, S.; Jin, M.; Ni, Z.; Zhang, B.; Zhang, D.; Luo, F.; et al. FGF/FGFR signaling in health and disease. Signal Transduct. Target. Ther. 2020, 5, 181. [Google Scholar] [CrossRef] [PubMed]
- De Luca, A.; Esposito Abate, R.; Rachiglio, A.M.; Maiello, M.R.; Esposito, C.; Schettino, C.; Izzo, F.; Nasti, G.; Normanno, N. FGFR Fusions in Cancer: From Diagnostic Approaches to Therapeutic Intervention. Int. J. Mol. Sci. 2020, 21, 6856. [Google Scholar] [CrossRef] [PubMed]
- Kouhara, H.; Hadari, Y.R.; Spivak-Kroizman, T.; Schilling, J.; Bar-Sagi, D.; Lax, I.; Schlessinger, J. A lipid-anchored Grb2-binding protein that links FGF-receptor activation to the Ras/MAPK signaling pathway. Cell 1997, 89, 693–702. [Google Scholar] [CrossRef]
- Lamothe, B.; Yamada, M.; Schaeper, U.; Birchmeier, W.; Lax, I.; Schlessinger, J. The docking protein Gab1 is an essential component of an indirect mechanism for fibroblast growth factor stimulation of the phosphatidylinositol 3-kinase/Akt antiapoptotic pathway. Mol. Cell. Biol. 2004, 24, 5657–5666. [Google Scholar] [CrossRef]
- Dudka, A.A.; Sweet, S.M.; Heath, J.K. Signal transducers and activators of transcription-3 binding to the fibroblast growth factor receptor is activated by receptor amplification. Cancer Res. 2010, 70, 3391–3401. [Google Scholar] [CrossRef] [PubMed]
- Ornitz, D.M.; Itoh, N. The Fibroblast Growth Factor signaling pathway. Wiley Interdiscip. Rev. Dev. Biol. 2015, 4, 215–266. [Google Scholar] [CrossRef]
- Liu, G.; Chen, T.; Ding, Z.; Wang, Y.; Wei, Y.; Wei, X. Inhibition of FGF-FGFR and VEGF-VEGFR signalling in cancer treatment. Cell Prolif. 2021, 54, e13009. [Google Scholar] [CrossRef] [PubMed]
- Helsten, T.; Elkin, S.; Arthur, E.; Tomson, B.N.; Carter, J.; Kurzrock, R. The FGFR Landscape in Cancer: Analysis of 4,853 Tumors by Next-Generation Sequencing. Clin. Cancer Res. 2016, 22, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.Y.; Kim, E.; Cheung, H.W.; Weir, B.A.; Dunn, G.P.; Shen, R.R.; Hahn, W.C. The Tyrosine Kinase Adaptor Protein FRS2 Is Oncogenic and Amplified in High-Grade Serous Ovarian Cancer. Mol. Cancer Res. 2015, 13, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.Y.; Hahn, W.C. Oncogenic Signaling Adaptor Proteins. J. Genet. Genom. 2015, 42, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Xie, G.; Zhang, Z.; Wang, Z.; Wang, Y.; Wang, Y.; Qiu, Y.; Li, L.; Bu, H.; Li, J.; et al. MiR-4653-3p and its target gene FRS2 are prognostic biomarkers for hormone receptor positive breast cancer patients receiving tamoxifen as adjuvant endocrine therapy. Oncotarget 2016, 7, 61166–61182. [Google Scholar] [CrossRef]
- Fan, G.; Jiao, J.; Shen, F.; Chu, F. Upregulation of lncRNA ZFAS1 promotes lung adenocarcinoma progression by sponging miR-1271-5p and upregulating FRS2. Thorac. Cancer 2020, 11, 2178–2187. [Google Scholar] [CrossRef]
- Liu, J.; You, P.; Chen, G.; Fu, X.; Zeng, X.; Wang, C.; Huang, Y.; An, L.; Wan, X.; Navone, N.; et al. Hyperactivated FRS2alpha-mediated signaling in prostate cancer cells promotes tumor angiogenesis and predicts poor clinical outcome of patients. Oncogene 2016, 35, 1750–1759. [Google Scholar] [CrossRef]
- Aytatli, A.; Barlak, N.; Sanli, F.; Caglar, H.O.; Gundogdu, B.; Tatar, A.; Ittmann, M.; Karatas, O.F. AZD4547 targets the FGFR/Akt/SOX2 axis to overcome paclitaxel resistance in head and neck cancer. Cell. Oncol. 2022, 45, 41–56. [Google Scholar] [CrossRef]
- Crawford, K.; Bontrager, E.; Schwarz, M.A.; Chaturvedi, A.; Lee, D.D.; Md Sazzad, H.; von Holzen, U.; Zhang, C.; Schwarz, R.E.; Awasthi, N. Targeted FGFR/VEGFR/PDGFR inhibition with dovitinib enhances the effects of nab-paclitaxel in preclinical gastric cancer models. Cancer Biol. Ther. 2021, 22, 619–629. [Google Scholar] [CrossRef] [PubMed]
- McSheehy, P.M.J.; Forster-Gross, N.; El Shemerly, M.; Bachmann, F.; Roceri, M.; Hermann, N.; Spickermann, J.; Kellenberger, L.; Lane, H.A. The fibroblast growth factor receptor inhibitor, derazantinib, has strong efficacy in human gastric tumor models and synergizes with paclitaxel in vivo. Anticancer Drugs 2023, 34, 532–543. [Google Scholar] [CrossRef] [PubMed]
- Boichuk, S.; Dunaev, P.; Mustafin, I.; Mani, S.; Syuzov, K.; Valeeva, E.; Bikinieva, F.; Galembikova, A. Infigratinib (BGJ 398), a Pan-FGFR Inhibitor, Targets P-Glycoprotein and Increases Chemotherapeutic-Induced Mortality of Multidrug-Resistant Tumor Cells. Biomedicines 2022, 10, 601. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Tian, H.; Yue, W.; Li, L.; Li, S.; Gao, C.; Si, L.; Qi, L.; Lu, M.; Hao, B.; et al. Rotenone induces apoptosis in human lung cancer cells by regulating autophagic flux. IUBMB Life 2016, 68, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Liu, Y.; Dai, M.; Li, Y.; Peng, R.; Yu, S.; Liu, H. Rotenone restrains colon cancer cell viability, motility and epithelial-mesenchymal transition and tumorigenesis in nude mice via the PI3K/AKT pathway. Int. J. Mol. Med. 2020, 46, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.T.; Huang, H.C.; Lin, J.K. Rotenone induces apoptosis in MCF-7 human breast cancer cell-mediated ROS through JNK and p38 signaling. Mol. Carcinog. 2010, 49, 141–151. [Google Scholar] [CrossRef]
- Du, J.; Jiang, F.; Xu, S.S.; Huang, Z.F.; Chen, L.L.; Li, L. Tephrosin induces apoptosis of human pancreatic cancer cells through the generation of reactive oxygen species. J. Cancer 2021, 12, 270–280. [Google Scholar] [CrossRef]
- Li, J.; Wang, X.L.; Fang, Y.C.; Wang, C.Y. Tephrosin-induced autophagic cell death in A549 non-small cell lung cancer cells. J. Asian Nat. Prod. Res. 2010, 12, 992–1000. [Google Scholar] [CrossRef]
- Maloney, S.M.; Hoover, C.A.; Morejon-Lasso, L.V.; Prosperi, J.R. Mechanisms of Taxane Resistance. Cancers 2020, 12, 3323. [Google Scholar] [CrossRef]
- Abedini, M.R.; Muller, E.J.; Bergeron, R.; Gray, D.A.; Tsang, B.K. Akt promotes chemoresistance in human ovarian cancer cells by modulating cisplatin-induced, p53-dependent ubiquitination of FLICE-like inhibitory protein. Oncogene 2010, 29, 11–25. [Google Scholar] [CrossRef]
- Salaroglio, I.C.; Mungo, E.; Gazzano, E.; Kopecka, J.; Riganti, C. ERK is a Pivotal Player of Chemo-Immune-Resistance in Cancer. Int. J. Mol. Sci. 2019, 20, 2505. [Google Scholar] [CrossRef]
- Du, Z.; Lovly, C.M. Mechanisms of receptor tyrosine kinase activation in cancer. Mol. Cancer 2018, 17, 58. [Google Scholar] [CrossRef]
- Yamaoka, T.; Kusumoto, S.; Ando, K.; Ohba, M.; Ohmori, T. Receptor Tyrosine Kinase-Targeted Cancer Therapy. Int. J. Mol. Sci. 2018, 19, 3491. [Google Scholar] [CrossRef]
- Butti, R.; Das, S.; Gunasekaran, V.P.; Yadav, A.S.; Kumar, D.; Kundu, G.C. Receptor tyrosine kinases (RTKs) in breast cancer: Signaling, therapeutic implications and challenges. Mol. Cancer 2018, 17, 34. [Google Scholar] [CrossRef]
- Wei, Y.; Erfani, S.; Schweer, D.; de Gouvea, R.; Qadir, J.; Shi, J.; Cheng, K.; Wu, D.; Craven, R.; Wu, Y.; et al. Targeting receptor tyrosine kinases in ovarian cancer: Genomic dysregulation, clinical evaluation of inhibitors, and potential for combinatorial therapies. Mol. Ther. Oncolytics 2023, 28, 293–306. [Google Scholar] [CrossRef]
- Orlicky, J.; Sulova, Z.; Dovinova, I.; Fiala, R.; Zahradnikova, A., Jr.; Breier, A. Functional fluo-3/AM assay on P-glycoprotein transport activity in L1210/VCR cells by confocal microscopy. Gen. Physiol. Biophys. 2004, 23, 357–366. [Google Scholar]
- Goto, T.; Takano, M.; Hirata, J.; Tsuda, H. The involvement of FOXO1 in cytotoxic stress and drug-resistance induced by paclitaxel in ovarian cancers. Br. J. Cancer 2008, 98, 1068–1075. [Google Scholar] [CrossRef]
- Varbiro, G.; Veres, B.; Gallyas, F., Jr.; Sumegi, B. Direct effect of Taxol on free radical formation and mitochondrial permeability transition. Free Radic. Biol. Med. 2001, 31, 548–558. [Google Scholar] [CrossRef]
- Zhang, P.; Qin, D.; Chen, J.; Zhang, Z. Plants in the Genus Tephrosia: Valuable Resources for Botanical Insecticides. Insects 2020, 11, 721. [Google Scholar] [CrossRef]
- Innos, J.; Hickey, M.A. Using Rotenone to Model Parkinson’s Disease in Mice: A Review of the Role of Pharmacokinetics. Chem. Res. Toxicol. 2021, 34, 1223–1239. [Google Scholar] [CrossRef]
- Choi, S.; Choi, Y.; Dat, N.T.; Hwangbo, C.; Lee, J.J.; Lee, J.H. Tephrosin induces internalization and degradation of EGFR and ErbB2 in HT-29 human colon cancer cells. Cancer Lett. 2010, 293, 23–30. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Yu, J.Y.; Leng, Y.L.; Zhu, R.R.; Liu, H.X.; Wang, X.Y.; Yang, T.T.; Guo, Y.N.; Tang, J.L.; Zhang, X.C. MiR-181c sensitizes ovarian cancer cells to paclitaxel by targeting GRP78 through the PI3K/Akt pathway. Cancer Gene Ther. 2022, 29, 770–783. [Google Scholar] [CrossRef]
- Park, G.B.; Kim, D. MicroRNA-503-5p Inhibits the CD97-Mediated JAK2/STAT3 Pathway in Metastatic or Paclitaxel-Resistant Ovarian Cancer Cells. Neoplasia 2019, 21, 206–215. [Google Scholar] [CrossRef]
- Hossein, G.; Halvaei, S.; Heidarian, Y.; Dehghani-Ghobadi, Z.; Hassani, M.; Hosseini, H.; Naderi, N.; Sheikh Hassani, S. Pectasol-C Modified Citrus Pectin targets Galectin-3-induced STAT3 activation and synergize paclitaxel cytotoxic effect on ovarian cancer spheroids. Cancer Med. 2019, 8, 4315–4329. [Google Scholar] [CrossRef]
- Sheng, H.; Feng, Q.; Quan, Q.; Sheng, X.; Zhang, P. Inhibition of STAT3 reverses Taxol-resistance in ovarian cancer by down-regulating G6PD expression in vitro. Biochem. Biophys. Res. Commun. 2022, 617, 62–68. [Google Scholar] [CrossRef]
- Sanchez-Carranza, J.N.; Diaz, J.F.; Redondo-Horcajo, M.; Barasoain, I.; Alvarez, L.; Lastres, P.; Romero-Estrada, A.; Aller, P.; Gonzalez-Maya, L. Gallic acid sensitizes paclitaxel-resistant human ovarian carcinoma cells through an increase in reactive oxygen species and subsequent downregulation of ERK activation. Oncol. Rep. 2018, 39, 3007–3014. [Google Scholar] [CrossRef]
- Mao, Y.; Xu, R. Circular RNA CDR1-AS contributes to pemetrexed and cisplatin chemoresistance through EGFR/PI3K signaling pathway in lung adenocarcinoma. Biomed. Pharmacother. 2020, 123, 109771. [Google Scholar] [CrossRef]
- Dong, M.; Li, P.; Xie, Y.; Wang, Z.; Wang, R. CircMYBL2 regulates the resistance of cervical cancer cells to paclitaxel via miR-665-dependent regulation of EGFR. Drug Dev. Res. 2021, 82, 1193–1205. [Google Scholar] [CrossRef]
- Qiu, L.; Di, W.; Jiang, Q.; Scheffler, E.; Derby, S.; Yang, J.; Kouttab, N.; Wanebo, H.; Yan, B.; Wan, Y. Targeted inhibition of transient activation of the EGFR-mediated cell survival pathway enhances paclitaxel-induced ovarian cancer cell death. Int. J. Oncol. 2005, 27, 1441–1448. [Google Scholar] [CrossRef]
- Szymczyk, J.; Sluzalska, K.D.; Materla, I.; Opalinski, L.; Otlewski, J.; Zakrzewska, M. FGF/FGFR-Dependent Molecular Mechanisms Underlying Anti-Cancer Drug Resistance. Cancers 2021, 13, 5796. [Google Scholar] [CrossRef]
- McDermott, S.C.; Rodriguez-Ramirez, C.; McDermott, S.P.; Wicha, M.S.; Nor, J.E. FGFR signaling regulates resistance of head and neck cancer stem cells to cisplatin. Oncotarget 2018, 9, 25148–25165. [Google Scholar] [CrossRef]
- Byron, S.A.; Loch, D.C.; Pollock, P.M. Fibroblast growth factor receptor inhibition synergizes with Paclitaxel and Doxorubicin in endometrial cancer cells. Int. J. Gynecol. Cancer 2012, 22, 1517–1526. [Google Scholar] [CrossRef]
- Na, Y.R.; Kim, J.Y.; Song, C.H.; Kim, M.; Do, Y.T.; Vo, T.T.L.; Choi, E.; Ha, E.; Seo, J.H.; Shin, S.J. The FGFR Family Inhibitor AZD4547 Exerts an Antitumor Effect in Ovarian Cancer Cells. Int. J. Mol. Sci. 2021, 22, 10817. [Google Scholar] [CrossRef]
- Cha, H.J.; Choi, J.H.; Park, I.C.; Kim, C.H.; An, S.K.; Kim, T.J.; Lee, J.H. Selective FGFR inhibitor BGJ398 inhibits phosphorylation of AKT and STAT3 and induces cytotoxicity in sphere-cultured ovarian cancer cells. Int. J. Oncol. 2017, 50, 1279–1288. [Google Scholar] [CrossRef]
- Szymczyk, J.; Sochacka, M.; Chudy, P.; Opalinski, L.; Otlewski, J.; Zakrzewska, M. FGF1 protects FGFR1-overexpressing cancer cells against drugs targeting tubulin polymerization by activating AKT via two independent mechanisms. Front. Oncol. 2022, 12, 1011762. [Google Scholar] [CrossRef]
- Goh, L.K.; Sorkin, A. Endocytosis of receptor tyrosine kinases. Cold Spring Harb. Perspect. Biol. 2013, 5, a017459. [Google Scholar] [CrossRef]
- Szybowska, P.; Kostas, M.; Wesche, J.; Haugsten, E.M.; Wiedlocha, A. Negative Regulation of FGFR (Fibroblast Growth Factor Receptor) Signaling. Cells 2021, 10, 1342. [Google Scholar] [CrossRef]
- Opalinski, L.; Sokolowska-Wedzina, A.; Szczepara, M.; Zakrzewska, M.; Otlewski, J. Antibody-induced dimerization of FGFR1 promotes receptor endocytosis independently of its kinase activity. Sci. Rep. 2017, 7, 7121. [Google Scholar] [CrossRef]
- Guo, W.; Dong, W.; Li, M.; Shen, Y. Mitochondria P-glycoprotein confers paclitaxel resistance on ovarian cancer cells. Onco Targets Ther. 2019, 12, 3881–3891. [Google Scholar] [CrossRef]
- Pellegrini, E.; Multari, G.; Gallo, F.R.; Vecchiotti, D.; Zazzeroni, F.; Condello, M.; Meschini, S. A natural product, voacamine, sensitizes paclitaxel-resistant human ovarian cancer cells. Toxicol. Appl. Pharmacol. 2022, 434, 115816. [Google Scholar] [CrossRef]
- Zhang, F.F.; Zhu, Y.F.; Zhao, Q.N.; Yang, D.T.; Dong, Y.P.; Jiang, L.; Xing, W.X.; Li, X.Y.; Xing, H.; Shi, M.; et al. Microvesicles mediate transfer of P-glycoprotein to paclitaxel-sensitive A2780 human ovarian cancer cells, conferring paclitaxel-resistance. Eur. J. Pharmacol. 2014, 738, 83–90. [Google Scholar] [CrossRef]
- Nasimian, A.; Farzaneh, P.; Tamanoi, F.; Bathaie, S.Z. Cytosolic and mitochondrial ROS production resulted in apoptosis induction in breast cancer cells treated with Crocin: The role of FOXO3a, PTEN and AKT signaling. Biochem. Pharmacol. 2020, 177, 113999. [Google Scholar] [CrossRef]
- Essers, M.A.; Weijzen, S.; de Vries-Smits, A.M.; Saarloos, I.; de Ruiter, N.D.; Bos, J.L.; Burgering, B.M. FOXO transcription factor activation by oxidative stress mediated by the small GTPase Ral and JNK. EMBO J. 2004, 23, 4802–4812. [Google Scholar] [CrossRef]
- Wang, T.H.; Wang, H.S.; Soong, Y.K. Paclitaxel-induced cell death: Where the cell cycle and apoptosis come together. Cancer 2000, 88, 2619–2628. [Google Scholar] [CrossRef]
- Ferlini, C.; Raspaglio, G.; Mozzetti, S.; Distefano, M.; Filippetti, F.; Martinelli, E.; Ferrandina, G.; Gallo, D.; Ranelletti, F.O.; Scambia, G. Bcl-2 down-regulation is a novel mechanism of paclitaxel resistance. Mol. Pharmacol. 2003, 64, 51–58. [Google Scholar] [CrossRef]
- Tabuchi, Y.; Matsuoka, J.; Gunduz, M.; Imada, T.; Ono, R.; Ito, M.; Motoki, T.; Yamatsuji, T.; Shirakawa, Y.; Takaoka, M.; et al. Resistance to paclitaxel therapy is related with Bcl-2 expression through an estrogen receptor mediated pathway in breast cancer. Int. J. Oncol. 2009, 34, 313–319. [Google Scholar]
- Chun, E.; Lee, K.Y. Bcl-2 and Bcl-xL are important for the induction of paclitaxel resistance in human hepatocellular carcinoma cells. Biochem. Biophys. Res. Commun. 2004, 315, 771–779. [Google Scholar] [CrossRef]
- Nestal de Moraes, G.; Delbue, D.; Silva, K.L.; Robaina, M.C.; Khongkow, P.; Gomes, A.R.; Zona, S.; Crocamo, S.; Mencalha, A.L.; Magalhaes, L.M.; et al. FOXM1 targets XIAP and Survivin to modulate breast cancer survival and chemoresistance. Cell. Signal. 2015, 27, 2496–2505. [Google Scholar] [CrossRef]
- Connolly, K.; Mitter, R.; Muir, M.; Jodrell, D.; Guichard, S. Stable XIAP knockdown clones of HCT116 colon cancer cells are more sensitive to TRAIL, taxanes and irradiation in vitro. Cancer Chemother. Pharmacol. 2009, 64, 307–316. [Google Scholar] [CrossRef]
- Tong, Q.S.; Zheng, L.D.; Wang, L.; Zeng, F.Q.; Chen, F.M.; Dong, J.H.; Lu, G.C. Downregulation of XIAP expression induces apoptosis and enhances chemotherapeutic sensitivity in human gastric cancer cells. Cancer Gene Ther. 2005, 12, 509–514. [Google Scholar] [CrossRef]
- Lai, S.W.; Bamodu, O.A.; Tsai, W.C.; Chang, Y.M.; Lee, W.H.; Yeh, C.T.; Chao, T.Y. The therapeutic targeting of the FGFR1/Src/NF-kappaB signaling axis inhibits pancreatic ductal adenocarcinoma stemness and oncogenicity. Clin. Exp. Metastasis 2018, 35, 663–677. [Google Scholar] [CrossRef]
- Suh, D.H.; Park, W.H.; Kim, M.; Kim, K.; No, J.H.; Kim, Y.B. HOXB9 Overexpression Confers Chemoresistance to Ovarian Cancer Cells by Inducing ERCC-1, MRP-2, and XIAP. Int. J. Mol. Sci. 2023, 24, 1249. [Google Scholar] [CrossRef]
- He, X.; Khurana, A.; Maguire, J.L.; Chien, J.; Shridhar, V. HtrA1 sensitizes ovarian cancer cells to cisplatin-induced cytotoxicity by targeting XIAP for degradation. Int. J. Cancer 2012, 130, 1029–1035. [Google Scholar] [CrossRef]
- Fraser, M.; Leung, B.; Jahani-Asl, A.; Yan, X.; Thompson, W.E.; Tsang, B.K. Chemoresistance in human ovarian cancer: The role of apoptotic regulators. Reprod. Biol. Endocrinol. 2003, 1, 66. [Google Scholar] [CrossRef]
- Cheng, J.Q.; Jiang, X.; Fraser, M.; Li, M.; Dan, H.C.; Sun, M.; Tsang, B.K. Role of X-linked inhibitor of apoptosis protein in chemoresistance in ovarian cancer: Possible involvement of the phosphoinositide-3 kinase/Akt pathway. Drug Resist. Updates 2002, 5, 131–146. [Google Scholar] [CrossRef] [PubMed]
- Mao, H.L.; Pang, Y.; Zhang, X.; Yang, F.; Zheng, J.; Wang, Y.; Liu, P. Smac peptide potentiates TRAIL- or paclitaxel-mediated ovarian cancer cell death in vitro and in vivo. Oncol. Rep. 2013, 29, 515–522. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.S.; Bae, S.; Lim, Y.J.; So, K.A.; Kim, T.J.; Bae, S.; Lee, J.H. Tephrosin Suppresses the Chemoresistance of Paclitaxel-Resistant Ovarian Cancer via Inhibition of FGFR1 Signaling Pathway. Biomedicines 2023, 11, 3155. https://doi.org/10.3390/biomedicines11123155
Kim HS, Bae S, Lim YJ, So KA, Kim TJ, Bae S, Lee JH. Tephrosin Suppresses the Chemoresistance of Paclitaxel-Resistant Ovarian Cancer via Inhibition of FGFR1 Signaling Pathway. Biomedicines. 2023; 11(12):3155. https://doi.org/10.3390/biomedicines11123155
Chicago/Turabian StyleKim, Hee Su, Sowon Bae, Ye Jin Lim, Kyeong A So, Tae Jin Kim, Seunghee Bae, and Jae Ho Lee. 2023. "Tephrosin Suppresses the Chemoresistance of Paclitaxel-Resistant Ovarian Cancer via Inhibition of FGFR1 Signaling Pathway" Biomedicines 11, no. 12: 3155. https://doi.org/10.3390/biomedicines11123155