
The Effect of Early Application of Synthetic Peptides 19-2.5 and 19-4LF to Improve Survival and Neurological Outcome in a Mouse Model of Cardiac Arrest and Resuscitation
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
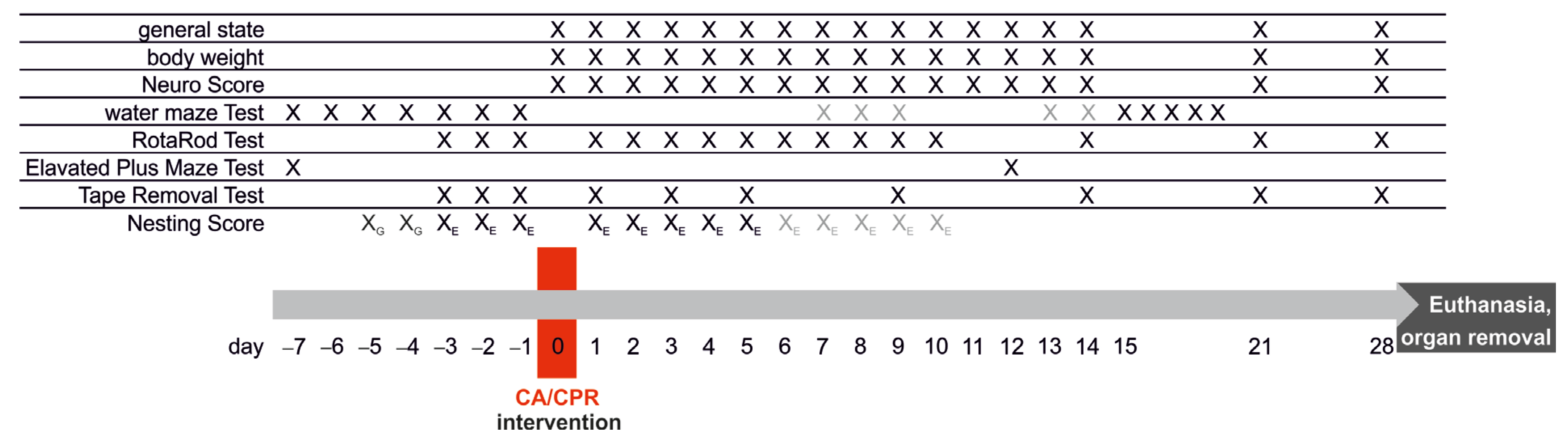
2.2. Study Groups and Experimental Protocol
2.3. Anesthesia
2.4. Protocol of Cardiac Arrest and Resuscitation
2.5. Parameters of Recovery Level and Well-Being of the Animals
2.6. Synthetic Peptides
2.7. Neurological Assessment
2.8. Blood Sampling and Tissue Preparation
2.9. Analysis of Plasma Samples
2.10. mRNA Expression Analyses from Tissue Samples
2.11. Statistics
3. Results
3.1. Long-Term Survival after Eight-Minute Cardiac Arrest and Resuscitation
3.2. Neurological Assessment
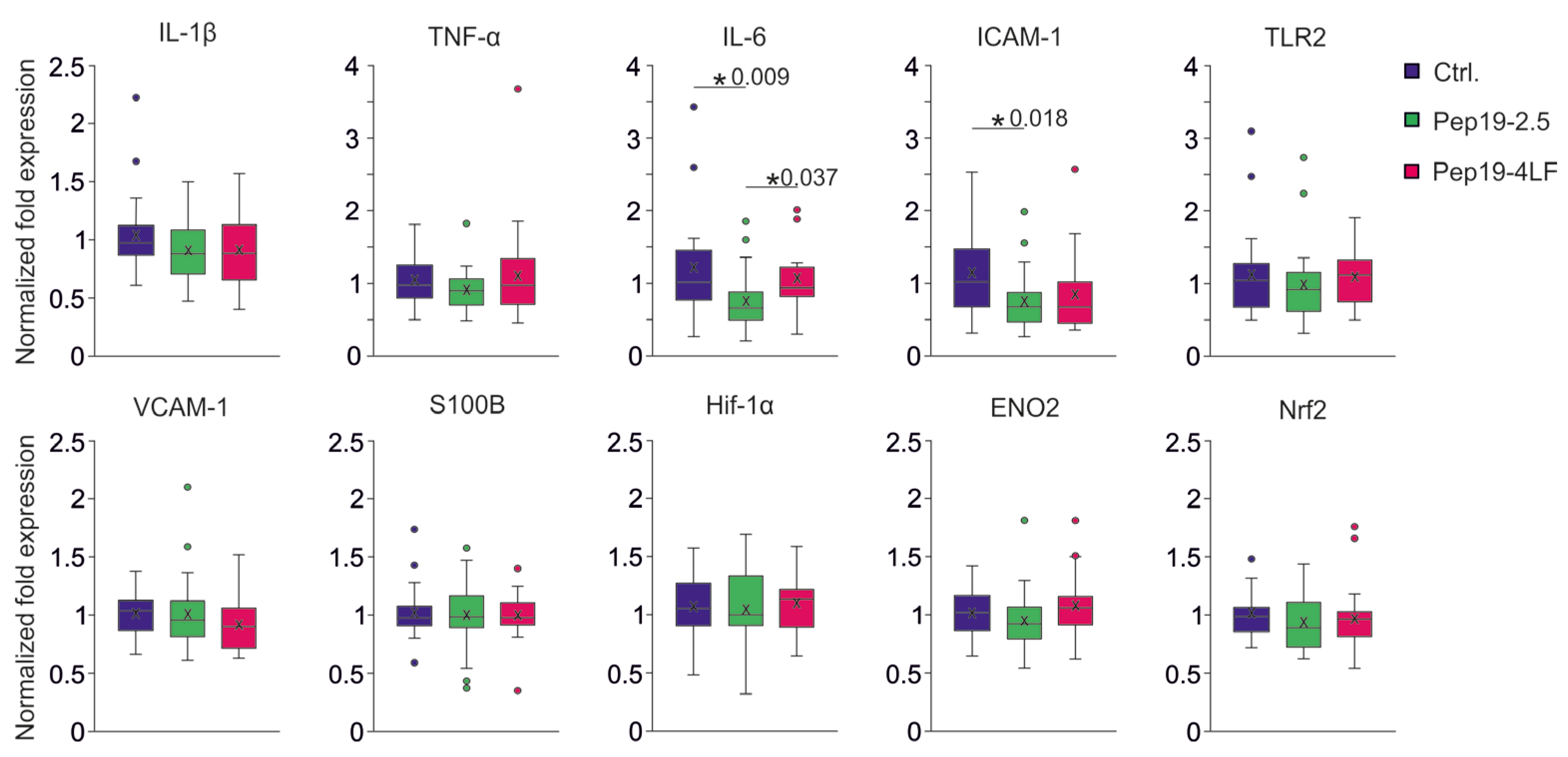
3.3. Plasma Biomarker Levels
3.4. mRNA Expression Levels Four Hours after CA-CPR in Brain Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Name | Primer | Sequence | Primer Length [bp] | Product Length [bp] | Efficiency [%] | Source | Primer Design | Primer Concen-tration | R2 |
|---|---|---|---|---|---|---|---|---|---|---|
| Hif1a | Hypoxia inducible factor 1 subunit alpha | Forward | TCAAGCAGCAGGAATTGGAAC | 21 | 181 | 95 | NCBI Primer- BLAST | Exon junction | 400 nM | 0.99 |
| Reverse | CTCATCCATTGACTGCCCCA | 20 | ||||||||
| s100b | S100 calcium binding protein B | Forward | GGTGACAAGCACAAGCTGAAG | 21 | 91 | 104 | NCBI Primer- BLAST | Exon junction | 400 nM | 0.97 |
| Reverse | CTTCCTGCTCCTTGATTTCCTCCA | 24 | ||||||||
| eno2 | Enolase 2 | Forward | AGGTGGATCTCTATACTGCCAAA | 23 | 98 | 96 | PrimerBank ID14290500a1 | Intron Inclusion | 200 nM | 0.99 |
| Reverse | GTCCCCATCCCTTAGTTCCAG | 21 | ||||||||
| ICAM-1 | Intercellular adhesion | Forward | GTGGGTCGAAGGTGGTTCTT | 20 | 188 | 105 | NCBI Primer- BLAST | Exon junction | 400 nM | 1 |
| molecule 1 | Reverse | CCGAGGACCATACAGCACG | 19 | |||||||
| VCAM-1 | Vascular cell adhesion molecule 1 | Forward | CAAAAAGGGACGATTCCGGC | 20 | 187 | 123 | NCBI Primer- BLAST | Intron Inclusion | 400 nM | 0.96 |
| Reverse | GTTTCGGGCACATTTCCACA | 20 | ||||||||
| Nrf2 | Nuclear factor, erythroid 2 like 2 | Forward | AGCAGGACATGGAGCAAGTT | 20 | 96 | 117 | NCBI Primer- BLAST | Intron Inclusion | 400 nM | 0.98 |
| Reverse | CAGCGGTAGTATCAGCCAGC | 20 | ||||||||
| Tnf | Tumor necrosis factor | Forward | CCTGTAGCCCACGTCGTAG | 19 | 148 | 120 | PrimerBank ID133892368c3 | Intron Inclusion | 500 nM | 0.98 |
| Reverse | GGGAGTAGACAAGGTACAACCC | 22 | ||||||||
| Il1β | Interleukin 1 beta | Forward | ATGAAAGACGGCACACCCAC | 20 | 175 | 129 | NCBI Primer- BLAST | Intron Inclusion | 400 nM | 0.99 |
| Reverse | GCTTGTGCTCTGCTTGTGAG | 20 | ||||||||
| Il6 | Interleukin 6 | Forward | GAGGATACCACTCCCAACAGACC | 23 | 141 | 106 | Aachen 078 | Intron Inclusion | 400 nM | 0.99 |
| Reverse | AAGTGCATCATCGTTGTTCATACA | 24 | ||||||||
| Tlr2 | Toll like receptor 2 | Forward | ACCTGAGAATGATGTGGGCG | 20 | 202 | 103 | NCBI Primer- BLAST | Non-Exon-junction | 400 nM | 0.99 |
| Reverse | CATTTGCCCGGAACGAAGTC | 20 | ||||||||
| Ppia | Peptidylprolyl isomerase A | Forward | GGCAAATGCTGGACCAAAC | 19 | 110 | 98 | Sundaram et al., PLoS one 2019. | Intron Inclusion | 400 nM | 0.99 |
| Reverse | CATTCCTGGACCCAAAACG | 19 | ||||||||
| s7 | Ribosomal protein S7 | Forward | AAAGTTCAGTGGCAAGCACG | 20 | 99 | 99 | NCBI Primer- BLAST | Intron Inclusion | 400 nM | 0.99 |
| Reverse | CTGGGGCGCTTCTGCTTATT | 20 |
References
- Lehrer, R.I.; Ganz, T. Antimicrobial peptides in mammalian and insect host defence. Curr. Opin. Immunol. 1999, 11 (Suppl. 1), 23–27. [Google Scholar] [CrossRef]
- Easton, D.M.; Nijnik, A.; Mayer, M.L.; Hancock, R.E.W. Potential of immunomodulatory host defense peptides as novel anti-infectives. Trends Biotechnol. 2009, 27, 582–590. [Google Scholar] [CrossRef]
- Gutsmann, T.; Razquin-Olazarán, I.; Kowalski, I.; Kaconis, Y.; Howe, J.; Bartels, R.; Hornef, M.; Schürholz, T.; Rössle, M.; Sanchez-Gómez, S.; et al. New antiseptic peptides to protect against endotoxin-mediated shock. Antimicrob. Agents Chemother. 2010, 54, 3817–3824. [Google Scholar] [CrossRef] [Green Version]
- Schuerholz, T.; Doemming, S.; Hornef, M.; Martin, L.; Simon, T.-P.; Heinbockel, L.; Brandenburg, K.; Marx, G. The anti-inflammatory effect of the synthetic antimicrobial peptide 19-2.5 in a murine sepsis model: A prospective randomized study. Crit. Care 2013, 17, R3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, L.; Van Meegern, A.; Doemming, S.; Schuerholz, T. Antimicrobial Peptides in Human Sepsis. Front. Immunol. 2015, 6, 404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfalzgraff, A.; Heinbockel, L.; Su, Q.; Gutsmann, T.; Brandenburg, K.; Weindl, G. Synthetic antimicrobial and LPS-neutralising peptides suppress inflammatory and immune responses in skin cells and promote keratinocyte migration. Sci. Rep. 2016, 6, 31577. [Google Scholar] [CrossRef] [Green Version]
- Yamada, N.; Martin, L.B.; Zechendorf, E.; Purvis, G.S.D.; Chiazza, F.; Varrone, B.; Collino, M.; Shepherd, J.; Heinbockel, L.; Gutsmann, T.; et al. Novel Synthetic, Host-defense Peptide Protects Against Organ Injury/Dysfunction in a Rat Model of Severe Hemorrhagic Shock. Ann. Surg. 2018, 268, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, S.; Al Zoubi, S.; Collotta, D.; Krieg, N.; Wissuwa, B.; Alves, G.F.; Purvis, G.S.D.; Norata, G.D.; Baragetti, A.; Catapano, A.L.; et al. A Synthetic Peptide Designed to Neutralize Lipopolysaccharides Attenuates Metaflammation and Diet-Induced Metabolic Derangements in Mice. Front. Immunol. 2021, 12, 701275. [Google Scholar] [CrossRef]
- Sandroni, C.; Cronberg, T.; Sekhon, M. Brain injury after cardiac arrest: Pathophysiology, treatment, and prognosis. Intensiv. Care Med. 2021, 47, 1393–1414. [Google Scholar] [CrossRef] [PubMed]
- Sekhon, M.S.; Ainslie, P.N.; Griesdale, D.E. Clinical pathophysiology of hypoxic ischemic brain injury after cardiac arrest: A “two-hit” model. Crit. Care 2017, 21, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Zhu, P.; Fujino, M.; Zhuang, J.; Guo, H.; Sheikh, I.; Zhao, L.; Li, X.-K. Oxidative Stress in Hypoxic-Ischemic Encephalopathy: Molecular Mechanisms and Therapeutic Strategies. Int. J. Mol. Sci. 2016, 17, 2078. [Google Scholar] [CrossRef] [Green Version]
- Meyer, A.S.; Johansson, P.I.; Kjaergaard, J.; Frydland, M.; Meyer, M.A.; Henriksen, H.H.; Thomsen, J.H.; Wiberg, S.C.; Hassager, C.; Ostrowski, S.R. Endothelial Dysfunction in Resuscitated Cardiac Arrest (ENDO-RCA): Safety and efficacy of low-dose Iloprost, a prostacyclin analogue, in addition to standard therapy, as compared to standard therapy alone, in post-cardiac-arrest-syndrome patients. Am. Heart J. 2020, 219, 9–20. [Google Scholar] [CrossRef]
- Jentzer, J.C.; Clements, C.M.; Murphy, J.G.; Wright, R.S. Recent developments in the management of patients resuscitated from cardiac arrest. J. Crit. Care 2017, 39, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Arslan, F.; Keogh, B.; McGuirk, P.; Parker, A.E. TLR2 and TLR4 in Ischemia Reperfusion Injury. Mediat. Inflamm. 2010, 2010, 704202. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ge, P.; Zhu, Y. TLR2 and TLR4 in the Brain Injury Caused by Cerebral Ischemia and Reperfusion. Mediat. Inflamm. 2013, 2013, 124614. [Google Scholar] [CrossRef] [Green Version]
- Bergt, S.; Güter, A.; Grub, A.; Wagner, N.-M.; Beltschany, C.; Langner, S.; Wree, A.; Hildebrandt, S.; Nöldge-Schomburg, G.; Vollmar, B.; et al. Impact of Toll-Like Receptor 2 Deficiency on Survival and Neurological Function after Cardiac Arrest: A Murine Model of Cardiopulmonary Resuscitation. PLoS ONE 2013, 8, e74944. [Google Scholar] [CrossRef]
- Ma, Q.; Zhang, Z.; Shim, J.-K.; Venkatraman, T.N.; Lascola, C.D.; Quinones, Q.J.; Mathew, J.P.; Terrando, N.; Podgoreanu, M.V. Annexin A1 Bioactive Peptide Promotes Resolution of Neuroinflammation in a Rat Model of Exsanguinating Cardiac Arrest Treated by Emergency Preservation and Resuscitation. Front. Neurosci. 2019, 13, 608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perkins, G.D.; Callaway, C.W.; Haywood, K.; Neumar, R.W.; Lilja, G.; Rowland, M.J.; Sawyer, K.N.; Skrifvars, M.B.; Nolan, J.P. Brain injury after cardiac arrest. Lancet 2021, 398, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Kirdajova, D.B.; Kriska, J.; Tureckova, J.; Anderova, M. Ischemia-Triggered Glutamate Excitotoxicity from the Perspective of Glial Cells. Front. Cell. Neurosci. 2020, 14, 51. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.H.Y.; Crack, P.J. Modulation of neuro-inflammation and vascular response by oxidative stress following cerebral ischemia-reperfusion injury. Curr. Med. Chem. 2008, 15, 1–14. [Google Scholar]
- Tsao, C.W.; Aday, A.W.; Almarzooq, Z.I.; Alonso, A.; Beaton, A.Z.; Bittencourt, M.S.; Boehme, A.K.; Buxton, A.E.; Carson, A.P.; Commodore-Mensah, Y.; et al. Heart Disease and Stroke Statistics—2022 Update: A Report From the American Heart Association. Circulation 2022, 145, e153–e639. [Google Scholar] [CrossRef] [PubMed]
- Gräsner, J.-T.; Wnent, J.; Herlitz, J.; Perkins, G.D.; Lefering, R.; Tjelmeland, I.; Koster, R.W.; Masterson, S.; Rossell-Ortiz, F.; Maurer, H.; et al. Survival after out-of-hospital cardiac arrest in Europe—Results of the EuReCa TWO study. Resuscitation 2020, 148, 218–226. [Google Scholar] [CrossRef]
- Gräsner, J.-T.; Herlitz, J.; Tjelmeland, I.B.; Wnent, J.; Masterson, S.; Lilja, G.; Bein, B.; Böttiger, B.W.; Rosell-Ortiz, F.; Nolan, J.P.; et al. European Resuscitation Council Guidelines 2021: Epidemiology of cardiac arrest in Europe. Resuscitation 2021, 161, 61–79. [Google Scholar] [CrossRef] [PubMed]
- Laver, S.; Farrow, C.; Turner, D.; Nolan, J. Mode of death after admission to an intensive care unit following cardiac arrest. Intensive Care Med. 2004, 30, 2126–2128. [Google Scholar] [CrossRef] [PubMed]
- Menzebach, A.; Bergt, S.; von Waldthausen, P.; Dinu, C.; Nöldge-Schomburg, G.; Vollmar, B. A comprehensive study of survival, tissue damage, and neurological dysfunction in a murine model of cardiopulmonary resuscitation after potassium induced cardiac arrest. Shock 2010, 33, 189–196. [Google Scholar] [CrossRef]
- Bergt, S.; Grub, A.; Mueller, M.; Bajorat, R.; Barilar, I.; Vollmar, B.; Roesner, J.P.; Wagner, N.-M. Toll-like receptor 4 deficiency or inhibition does not modulate survival and neurofunctional outcome in a murine model of cardiac arrest and resuscitation. PLoS ONE 2019, 14, e0220404. [Google Scholar] [CrossRef] [Green Version]
- Bajorat, R.; Kurth, J.; Stenzel, J.; Vollmar, B.; Krause, B.J.; Reuter, D.A.; Schuerholz, T.; Bergt, S. Early Post-ischemic Brain Glucose Metabolism Is Dependent on Function of TLR2: A Study Using [18F]F-FDG PET-CT in a Mouse Model of Cardiac Arrest and Cardiopulmonary Resuscitation. Mol. Imaging Biol. 2021, 24, 466–478. [Google Scholar] [CrossRef]
- Kumstel, S.; Wendt, E.H.U.; Eichberg, J.; Talbot, S.R.; Häger, C.; Zhang, X.; Abdelrahman, A.; Schönrogge, M.; Palme, R.; Bleich, A.; et al. Grading animal distress and side effects of therapies. Ann. N. Y. Acad. Sci. 2020, 1473, 20–34. [Google Scholar] [CrossRef] [Green Version]
- Abella, B.S.; Zhao, D.; Alvarado, J.; Hamann, K.; Hoek, T.L.V.; Becker, L.B. Intra-Arrest Cooling Improves Outcomes in a Murine Cardiac Arrest Model. Circulation 2004, 109, 2786–2791. [Google Scholar] [CrossRef] [Green Version]
- Neigh, G.N.; Glasper, E.R.; Kofler, J.; Traystman, R.J.; Mervis, R.F.; Bachstetter, A.; Devries, A.C. Cardiac arrest with cardiopulmonary resuscitation reduces dendritic spine density in CA1 pyramidal cells and selectively alters acquisition of spatial memory. Eur. J. Neurosci. 2004, 20, 1865–1872. [Google Scholar] [CrossRef]
- Hutchens, M.P.; Nakano, T.; Dunlap, J.; Traystman, R.J.; Hurn, P.D.; Alkayed, N.J. Soluble epoxide hydrolase gene deletion reduces survival after cardiac arrest and cardiopulmonary resuscitation. Resuscitation 2008, 76, 89–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neigh, G.N.; Kofler, J.; Meyers, J.L.; Bergdall, V.; La Perle, K.M.D.; Traystman, R.J.; Devries, A.C. Cardiac Arrest/Cardiopulmonary Resuscitation Increases Anxiety-Like Behavior and Decreases Social Interaction. J. Cereb. Blood Flow Metab. 2004, 24, 372–382. [Google Scholar] [CrossRef]
- Bouet, V.; Boulouard, M.; Toutain, J.; Divoux, D.; Bernaudin, M.; Schumann-Bard, P.; Freret, T. The adhesive removal test: A sensitive method to assess sensorimotor deficits in mice. Nat. Protoc. 2009, 4, 1560–1564. [Google Scholar] [CrossRef]
- Albertsmeier, M.; Teschendorf, P.; Popp, E.; Galmbacher, R.; Vogel, P.; Böttiger, B.W. Evaluation of a tape removal test to assess neurological deficit after cardiac arrest in rats. Resuscitation 2007, 74, 552–558. [Google Scholar] [CrossRef]
- Bajorat, R.; Porath, K.; Kuhn, J.; Goßla, E.; Goerss, D.; Sellmann, T.; Köhling, R.; Kirschstein, T. Oral administration of the casein kinase 2 inhibitor TBB leads to persistent KCa2.2 channel up-regulation in the epileptic CA1 area and cortex, but lacks anti-seizure efficacy in the pilocarpine epilepsy model. Epilepsy Res. 2018, 147, 42–50. [Google Scholar] [CrossRef]
- Klatte, K.; Kirschstein, T.; Otte, D.; Pothmann, L.; Müller, L.; Tokay, T.; Kober, M.; Uebachs, M.; Zimmer, A.; Beck, H. Impaired D-Serine-Mediated Cotransmission Mediates Cognitive Dysfunction in Epilepsy. J. Neurosci. 2013, 33, 13066–13080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.C.; Akinyi, L.; Scharf, D.; Mo, J.; Larner, S.F.; Muller, U.; Oli, M.W.; Zheng, W.; Kobeissy, F.; Papa, L.; et al. Ubiquitin C-terminal hydrolase-L1 as a biomarker for ischemic and traumatic brain injury in rats. Eur. J. Neurosci. 2010, 31, 722–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [Green Version]
- Andersen, C.L.; Ledet-Jensen, J.; Ørntoft, T. Normalization of real-time quantitative RT-PCR data: A model based variance estimation approach to identify genes suited for normalization—Applied to bladder- and colon-cancer data-sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [Green Version]
- Huber-Lang, M.; Lambris, J.D.; Ward, P.A. Innate immune responses to trauma. Nat. Immunol. 2018, 19, 327–341. [Google Scholar] [CrossRef]
- Xiang, Y.; Zhao, H.; Wang, J.; Zhang, L.; Liu, A.; Chen, Y. Inflammatory mechanisms involved in brain injury following cardiac arrest and cardiopulmonary resuscitation. Biomed. Rep. 2016, 5, 11–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, L.; Schmitz, S.; De Santis, R.; Doemming, S.; Haase, H.; Hoeger, J.; Heinbockel, L.; Brandenburg, K.; Marx, G.; Schuerholz, T. Peptide 19-2.5 Inhibits Heparan Sulfate-Triggered Inflammation in Murine Cardiomyocytes Stimulated with Human Sepsis Serum. PLoS ONE 2015, 10, e0127584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katz, L.; Ebmeyer, U.; Safar, P.; Radovsky, A.; Neumar, R. Outcome Model of Asphyxial Cardiac Arrest in Rats. J. Cereb. Blood Flow Metab. 1995, 15, 1032–1039. [Google Scholar] [CrossRef]
- Zhu, Y.; Yang, G.Y.; Ahlemeyer, B.; Pang, L.; Che, X.M.; Culmsee, C.; Klumpp, S.; Krieglstein, J. Transforming growth factor-beta 1 increases bad phosphorylation and protects neurons against damage. J. Neurosci. 2002, 22, 3898–3909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spera, P.A.; Ellison, J.A.; Feuerstein, G.Z.; Barone, F.C. IL-10 reduces rat brain injury following focal stroke. Neurosci. Lett. 1998, 251, 189–192. [Google Scholar] [CrossRef]
- Huusko, J.; Merentie, M.; Dijkstra, M.H.; Ryhänen, M.-M.; Karvinen, H.; Rissanen, T.T.; Vanwildemeersch, M.; Hedman, M.; Lipponen, J.; Heinonen, S.E.; et al. The effects of VEGF-R1 and VEGF-R2 ligands on angiogenic responses and left ventricular function in mice. Cardiovasc. Res. 2009, 86, 122–130. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, K.M.; Le, A.; Duong, H.; Wu, Y.; Zhang, Q.; Messadi, D.V. Correlation between VEGF and HIF-1α expression in human oral squamous cell carcinoma. Exp. Mol. Pathol. 2004, 76, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Shweiki, D.; Itin, A.; Soffer, D.; Keshet, E. Vascular endothelial growth factor induced by hypoxia may mediate hypoxia-initiated angiogenesis. Nature 1992, 359, 843–845. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.-J.; Glushakova, O.; Mondello, S.; Van, K.; Hayes, R.L.; Lyeth, B.G. Acute Temporal Profiles of Serum Levels of UCH-L1 and GFAP and Relationships to Neuronal and Astroglial Pathology following Traumatic Brain Injury in Rats. J. Neurotrauma 2015, 32, 1179–1189. [Google Scholar] [CrossRef]
- Osier, N.D.; Bramlett, H.M.; Shear, D.A.; Mondello, S.; Carlson, S.W.; Dietrich, W.D.; Deng-Bryant, Y.; Wang, K.K.; Hayes, R.L.; Yang, Z.; et al. Kollidon VA64 Treatment in Traumatic Brain Injury: Operation Brain Trauma Therapy. J. Neurotrauma 2021, 38, 2454–2472. [Google Scholar] [CrossRef]
- Li, X.; Tang, Y.; Yao, Z.; Hu, S.; Zhou, H.; Mo, X.; She, C.; Lu, X.; Huang, G. FDG-PET/CT Assessment of the Cerebral Protective Effects of Hydrogen in Rabbits with Cardiac Arrest. Curr. Med. Imaging Former. Curr. Med. Imaging Rev. 2022, 18, 977–985. [Google Scholar] [CrossRef]
- Ebner, F.; Moseby-Knappe, M.; Mattsson-Carlgren, N.; Lilja, G.; Dragancea, I.; Undén, J.; Friberg, H.; Erlinge, D.; Kjaergaard, J.; Hassager, C.; et al. Serum GFAP and UCH-L1 for the prediction of neurological outcome in comatose cardiac arrest patients. Resuscitation 2020, 154, 61–68. [Google Scholar] [CrossRef]
- Hirano, T. Interleukin-6 and its relation to inflammation and disease. Clin. Immunol. Immunopathol. 1992, 62, S60–S65. [Google Scholar] [CrossRef]
- Akira, S.; Taga, T.; Kishimoto, T. Interleukin-6 in Biology and Medicine. Adv. Immunol. 1993, 54, 1–78. [Google Scholar] [CrossRef]
- Müller, N. The Role of Intercellular Adhesion Molecule-1 in the Pathogenesis of Psychiatric Disorders. Front. Pharmacol. 2019, 10, 1251. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, M.; Cooper, D.; Russell, J.; Salter, J.W.; Zhang, J.H.; Nanda, A.; Granger, D.N. Molecular Determinants of the Prothrombogenic and Inflammatory Phenotype Assumed by the Postischemic Cerebral Microcirculation. Stroke 2003, 34, 1777–1782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsberg, P.J.; Carpe’n, O.; Paetau, A.; Karjalainen-Lindsberg, M.-L.; Kaste, M. Endothelial ICAM-1 Expression Associated with Inflammatory Cell Response in Human Ischemic Stroke. Circulation 1996, 94, 939–945. [Google Scholar] [CrossRef] [PubMed]
- Shenkar, R.; Cohen, A.J.; Vestweber, D.; Miller, Y.E.; Tuder, R.; Abraham, E. Hemorrhage and resuscitation alter the expression of ICAM-1 and P-selectin in mice. J. Inflamm. 1995, 45, 248–259. [Google Scholar] [PubMed]
- Hofer, S.; Bopp, C.; Hoerner, C.; Plaschke, K.; Faden, R.M.; Martin, E.; Bardenheuer, H.J.; Weigand, M.A. Injury of the Blood Brain Barrier and Up-Regulation of ICAM-1 in Polymicrobial Sepsis. J. Surg. Res. 2008, 146, 276–281. [Google Scholar] [CrossRef]
- Jinxiang, Z.; Heshui, W.; Lin, W.; Jinhui, Z.; Hui, W.; Qichang, Z. TLR2 mRNA upregulation in ischemic lobes in mouse partial hepatic ischemia/reperfusion injury model. J. Huazhong Univ. Sci. Technol. 2004, 24, 144–146. [Google Scholar] [CrossRef]




| Experimental Groups | ||||
|---|---|---|---|---|
| Intervention: | Ctrl. (NaCl 0,9%) | Pep19-2.5 | Pep19-4LF | |
| n = 37 | n = 34 | n = 37 | ||
| Parameter | Mean ± SD | Mean ± SD | Mean ± SD | p Value |
| Baseline before CA | ||||
| heart rate (1/min) MAP (mm Hg) body temperature (°C) | 210.78 ± 23.81 | 220.71 ± 23.52 | 220.34 ± 25.15 | 0.141 |
| 64.95 ± 8.26 | 68.79 ± 9.03 | 70.32 ± 7.67 | 0.021 * | |
| 36.21 ± 0.28 | 36.06 ± 0.20 | 36.10 ± 0.20 | 0.033 † | |
| CA | ||||
| ROSC time (s) dosage epinephrine (μg) extubation (min) | 63.32 ± 27.74 | 70.76 ± 37.43 | 64.39 ± 43.85 | 0.375 |
| 13.38 ± 3.14 | 14.04 ± 3.84 | 12.89 ± 3.61 | 0.276 | |
| 169.53 ± 17.81 | 171.31 ± 14.22 | 175.06 ± 17.86 | 0.376 | |
| 1 h after CA | ||||
| heart rate (1/min) | 328.56 ± 75.80 | 352.00 ± 88.82 | 337.91 ± 78.92 | 0.763 |
| MAP (mm Hg) | 57.33 ± 8.98 | 56.00 ± 4.86 | 52.67 ± 2.92 | 0.592 |
| body temperature (°C) | 36.25 ± 0.26 | 36.30 ± 0.35 | 36.33 ± 0.27 | 0.379 |
| 2 h after CA | ||||
| heart rate (1/min) | 273.57 ± 71.99 | 298.64 ± 75.87 | 300.11 ± 76.86 | 0.448 |
| MAP (mm Hg) | 57.80 ± 7.52 | 55.00 ± 0.00 | 53.00 ± 2.00 | 0.664 |
| body temperature (°C) | 36.36 ± 0.36 | 36.39 ± 0.34 | 36.44 ± 0.31 | 0.356 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bajorat, R.; Danckert, L.; Ebert, F.; Bancken, T.; Bergt, S.; Klawitter, F.; Vollmar, B.; Reuter, D.A.; Schürholz, T.; Ehler, J. The Effect of Early Application of Synthetic Peptides 19-2.5 and 19-4LF to Improve Survival and Neurological Outcome in a Mouse Model of Cardiac Arrest and Resuscitation. Biomedicines 2023, 11, 855. https://doi.org/10.3390/biomedicines11030855
Bajorat R, Danckert L, Ebert F, Bancken T, Bergt S, Klawitter F, Vollmar B, Reuter DA, Schürholz T, Ehler J. The Effect of Early Application of Synthetic Peptides 19-2.5 and 19-4LF to Improve Survival and Neurological Outcome in a Mouse Model of Cardiac Arrest and Resuscitation. Biomedicines. 2023; 11(3):855. https://doi.org/10.3390/biomedicines11030855
Chicago/Turabian StyleBajorat, Rika, Lena Danckert, Florian Ebert, Theresa Bancken, Stefan Bergt, Felix Klawitter, Brigitte Vollmar, Daniel A. Reuter, Tobias Schürholz, and Johannes Ehler. 2023. "The Effect of Early Application of Synthetic Peptides 19-2.5 and 19-4LF to Improve Survival and Neurological Outcome in a Mouse Model of Cardiac Arrest and Resuscitation" Biomedicines 11, no. 3: 855. https://doi.org/10.3390/biomedicines11030855