


Co-Expression of Chromatin Assembly Factor 1 Subunit A and Proliferating Cell Nuclear Antigen Is a Prognostic Biomarker of Esophageal Cancer
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. SiRNAs and Chemical
2.3. Western Blotting
2.4. Cell Fractionation
2.5. DNA Fiber Assay
2.6. Flow Cytometry
2.7. PLA
2.8. Clonogenic Assay
2.9. Quantification and Statistical Analysis
3. Results
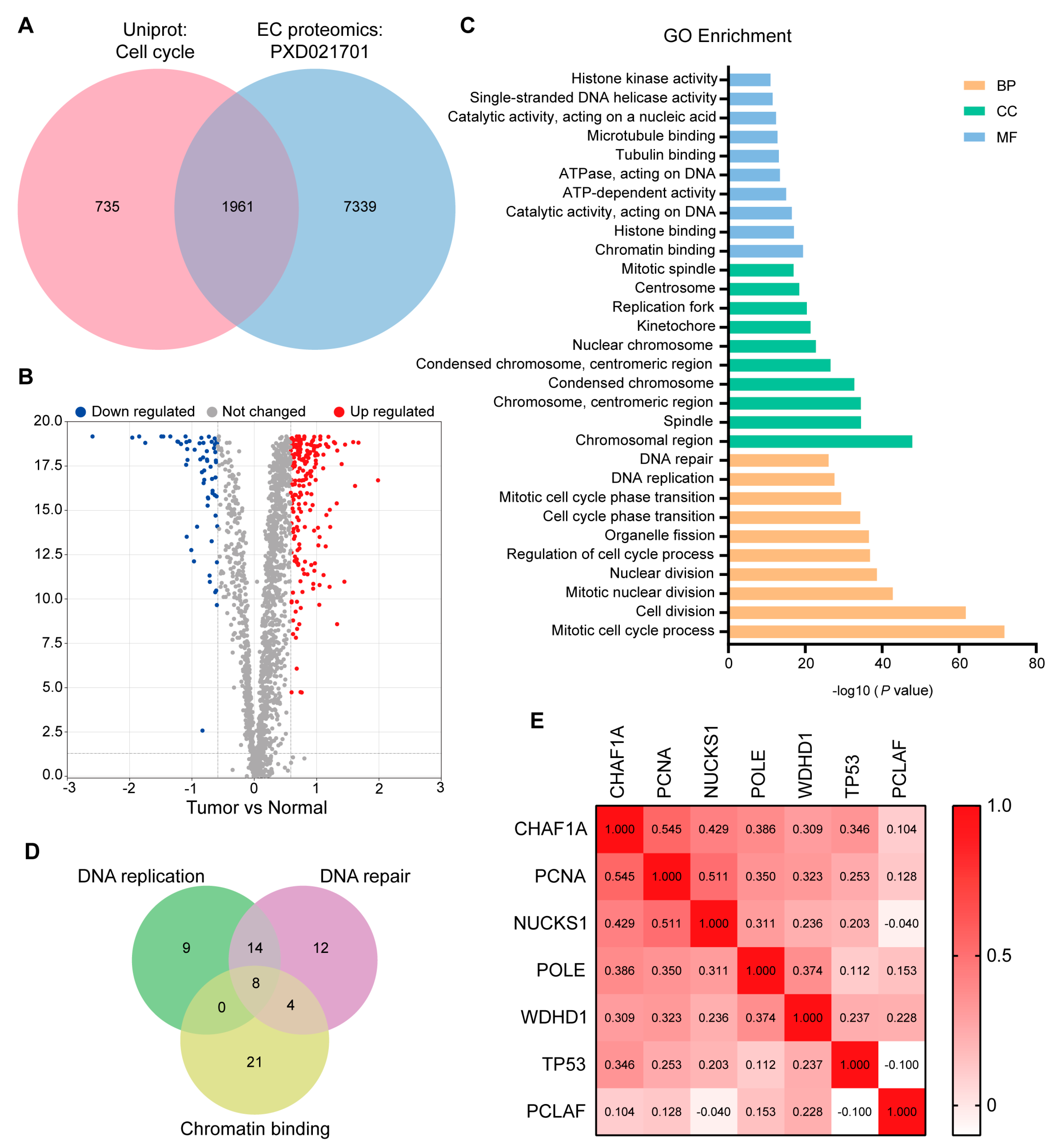
3.1. Screening Key DNA Replication and Repair Proteins in the EC Proteome
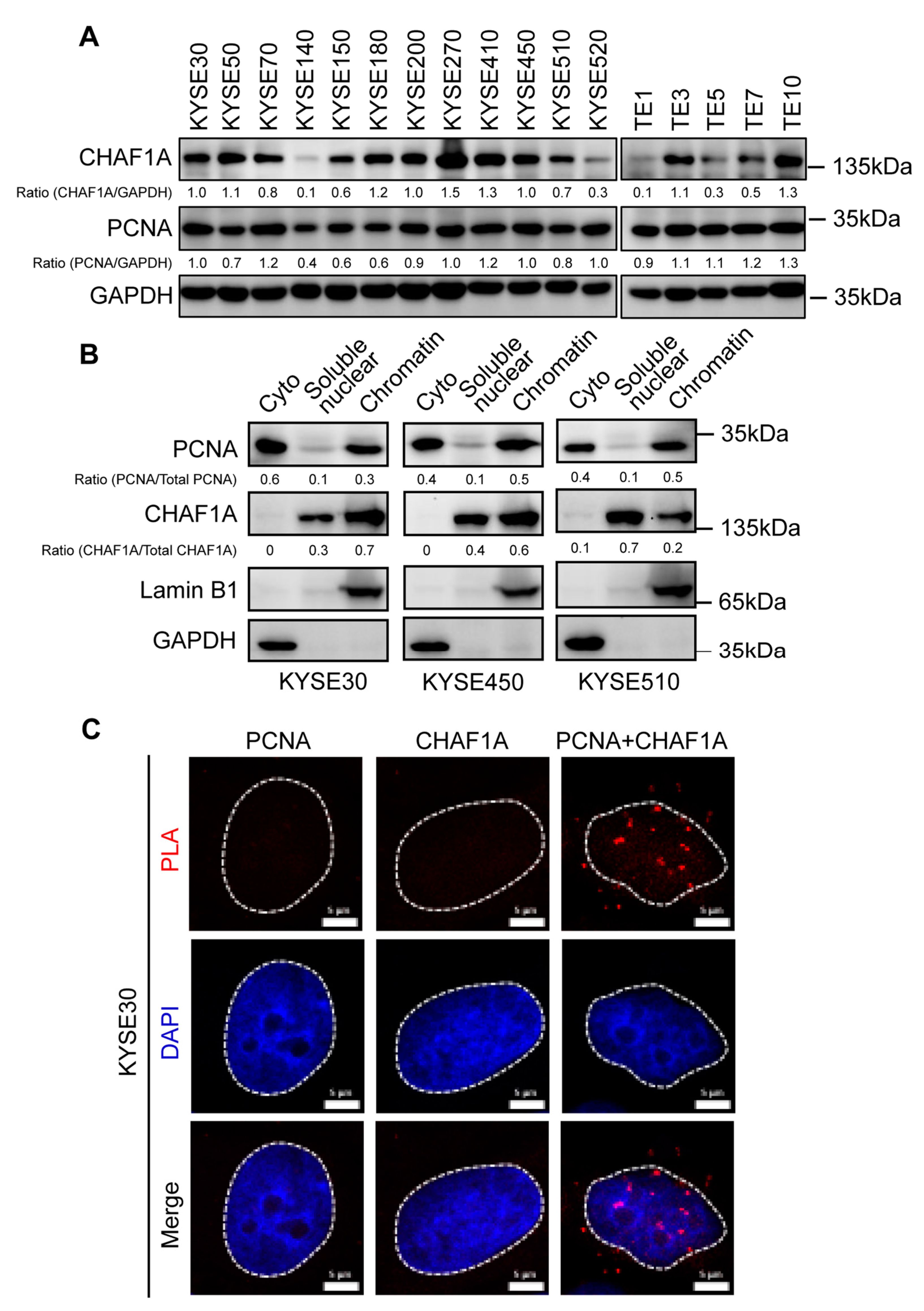
3.2. CHAF1A Interacts with PCNA in the Nucleus of EC Cells
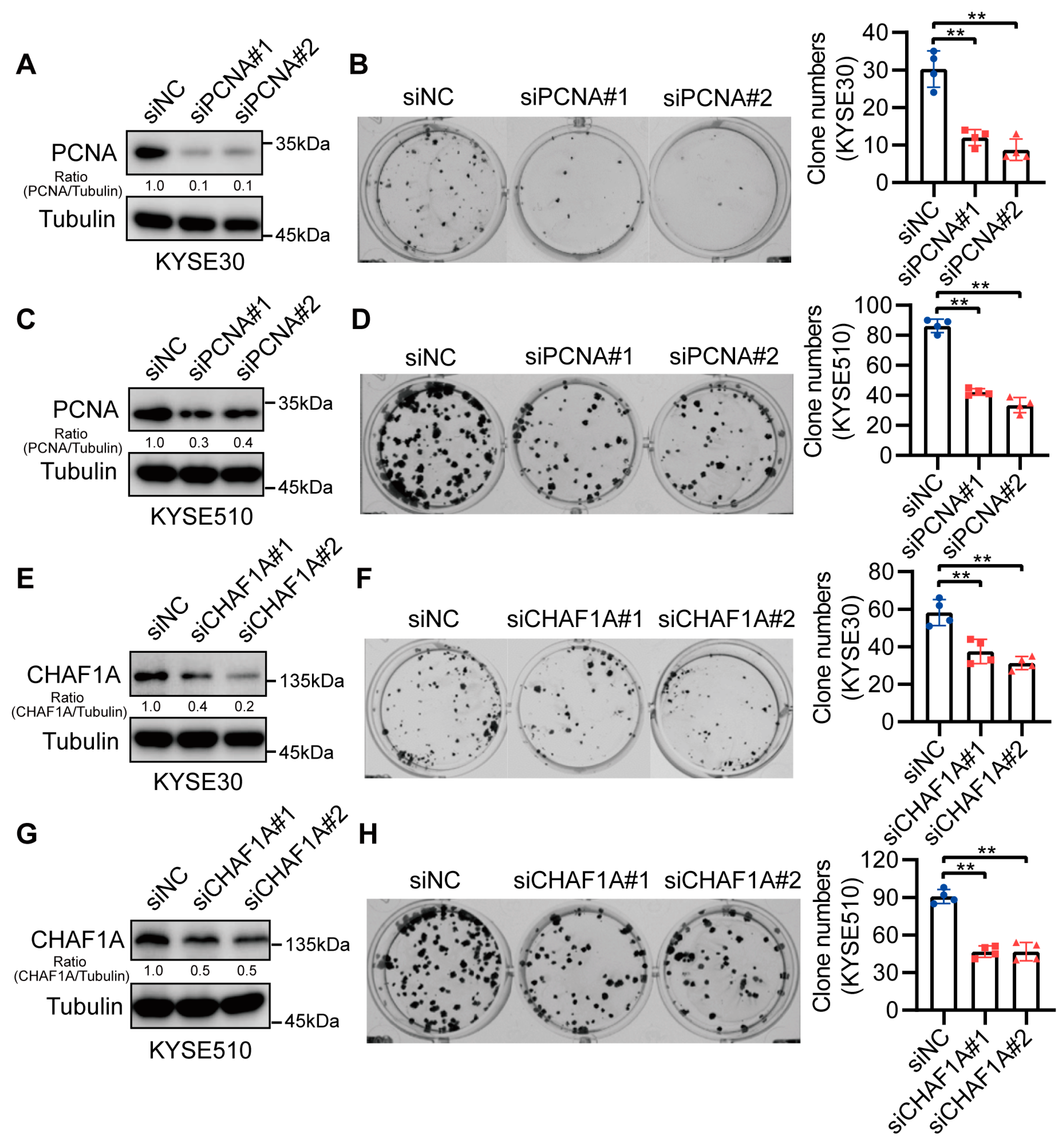
3.3. CHAF1A and PCNA Promote EC Cell Proliferation
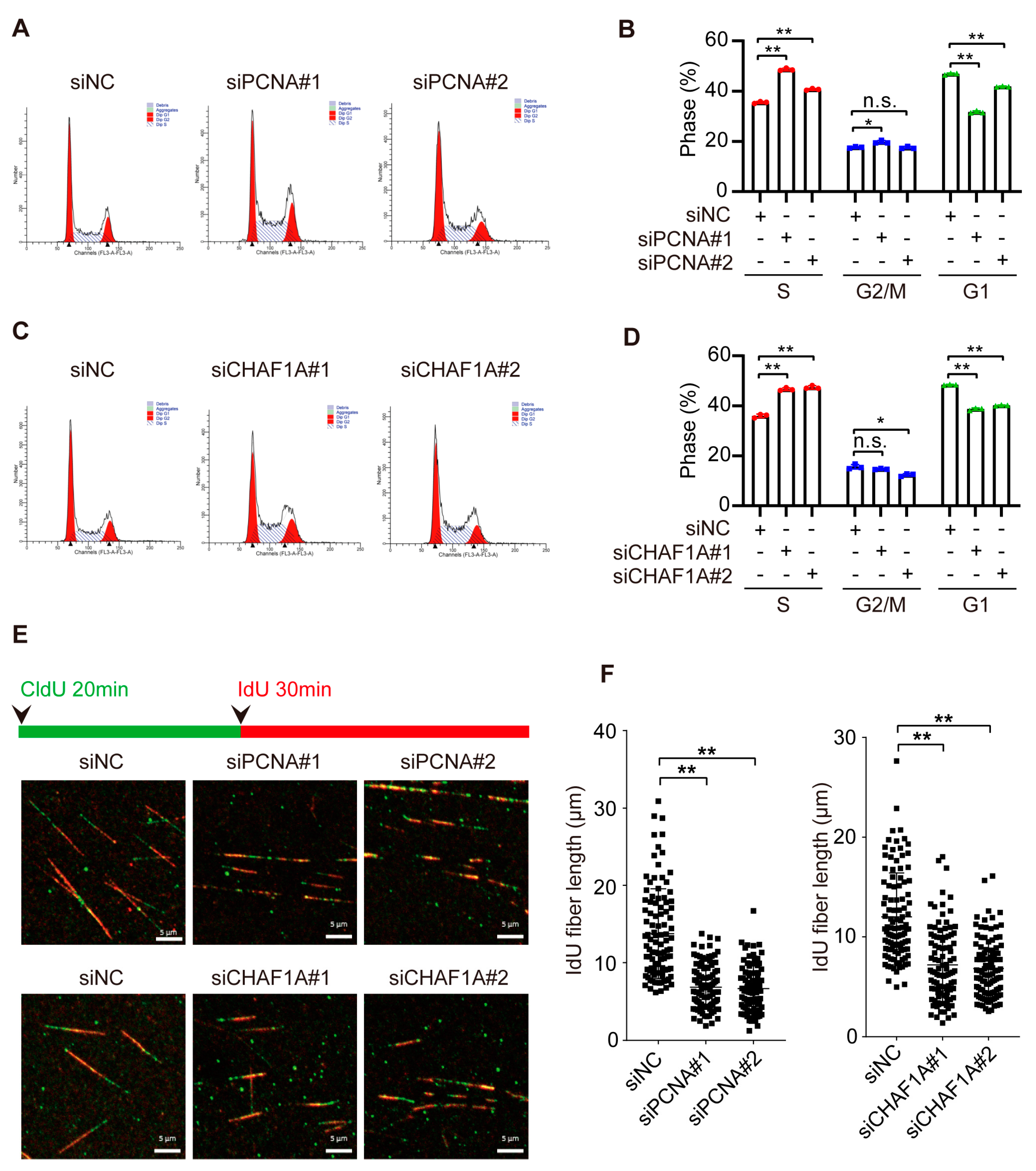
3.4. CHAF1A and PCNA Accelerate DNA Replication in EC Cells
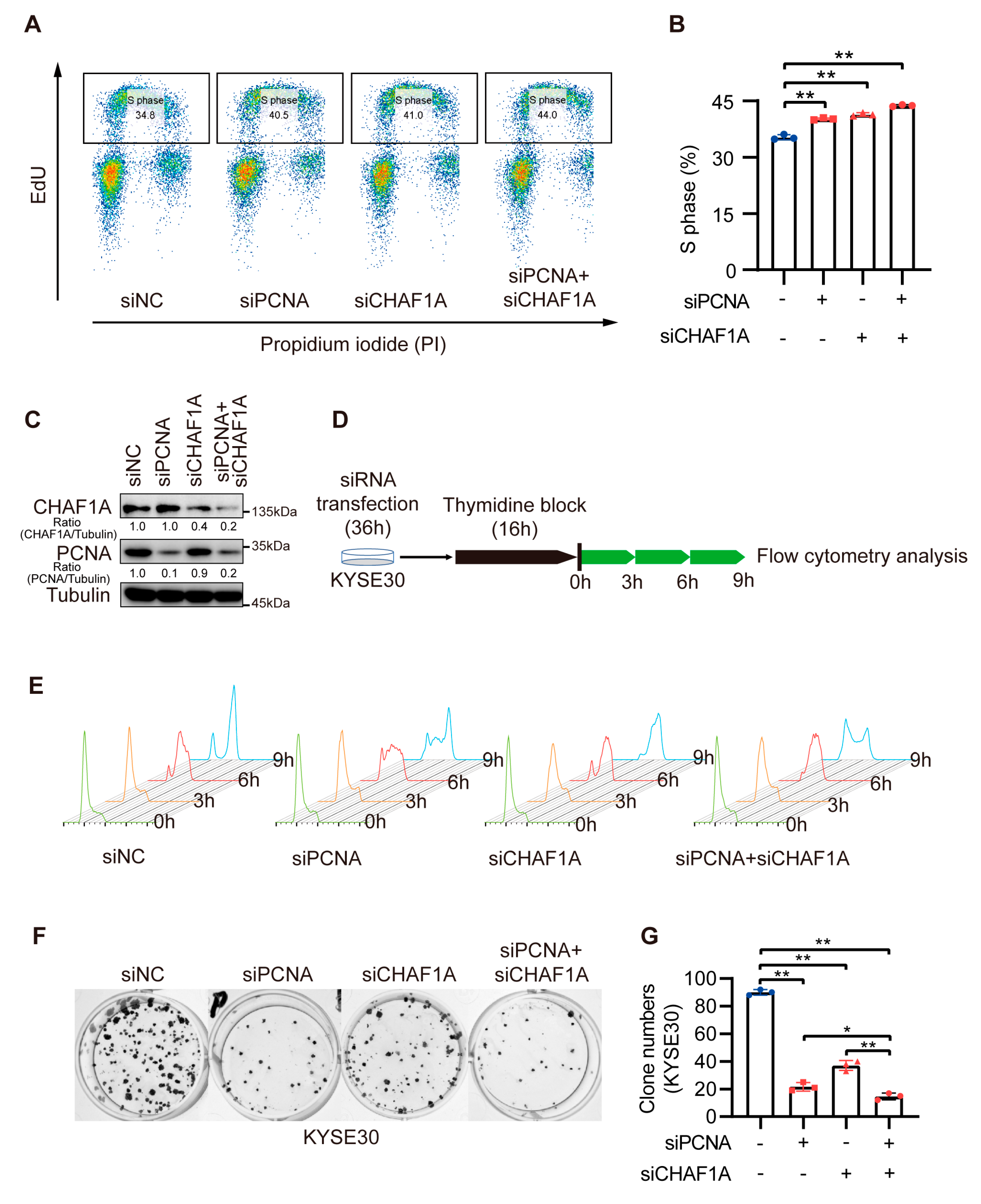
3.5. CHAF1A and PCNA Cooperate to Promote the S-Phase Progression of EC Cells
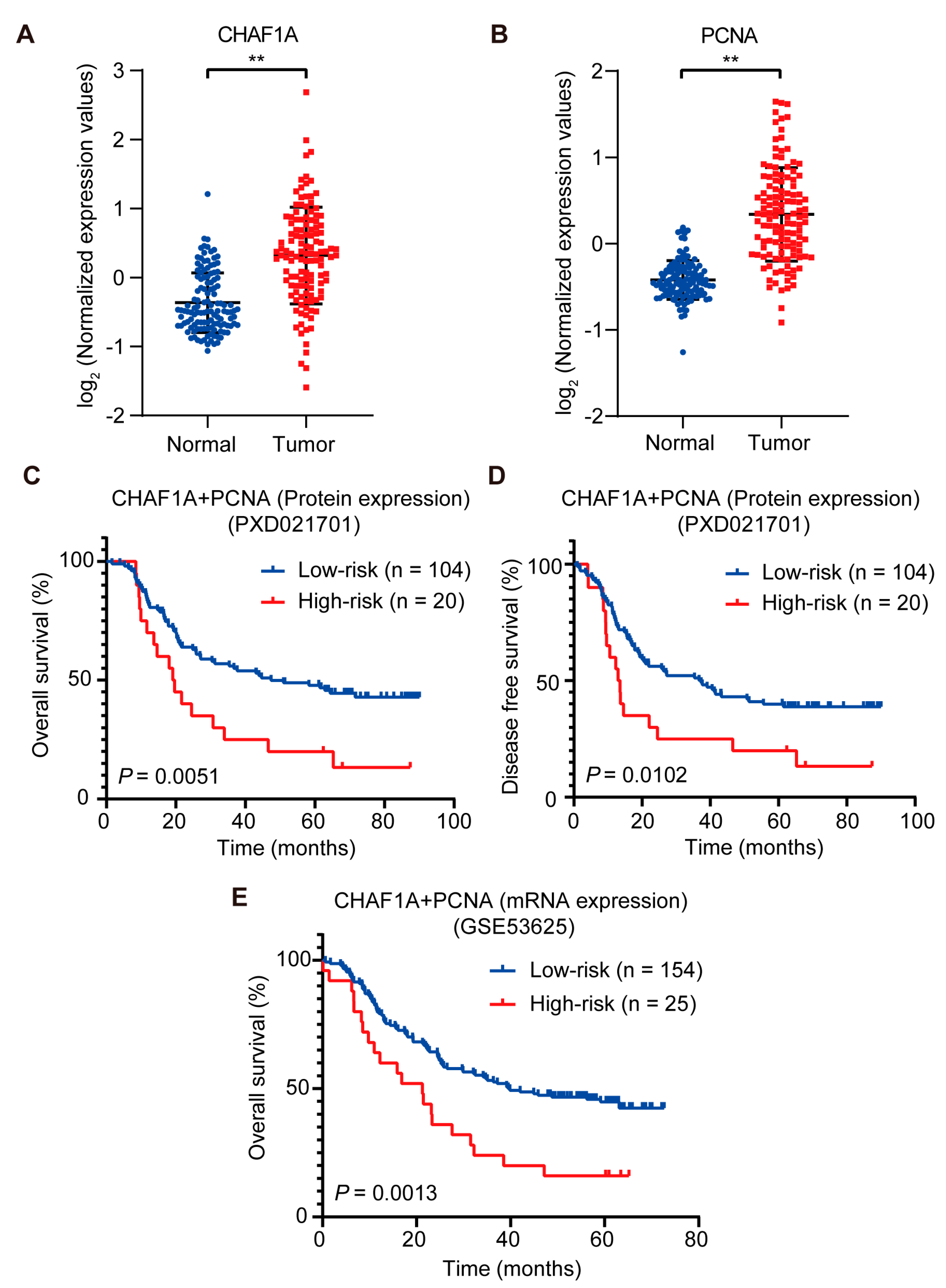
3.6. High CHAF1A and PCNA Expression Are Associated with Poor Prognosis in EC
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Smyth, E.C.; Lagergren, J.; Fitzgerald, R.C.; Lordick, F.; Shah, M.A.; Lagergren, P.; Cunningham, D. Oesophageal cancer. Nat. Rev. Dis. Primers 2017, 3, 17048. [Google Scholar] [CrossRef]
- Gavin, A.T.; Francisci, S.; Foschi, R.; Donnelly, D.W.; Lemmens, V.; Brenner, H.; Anderson, L.A.; Group, E.-W. Oesophageal cancer survival in Europe: A EUROCARE-4 study. Cancer Epidemiol. 2012, 36, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Njei, B.; McCarty, T.R.; Birk, J.W. Trends in esophageal cancer survival in United States adults from 1973 to 2009: A SEER database analysis. J. Gastroenterol. Hepatol. 2016, 31, 1141–1146. [Google Scholar] [CrossRef]
- Zeng, H.; Zheng, R.; Guo, Y.; Zhang, S.; Zou, X.; Wang, N.; Zhang, L.; Tang, J.; Chen, J.; Wei, K.; et al. Cancer survival in China, 2003–2005: A population-based study. Int. J. Cancer 2015, 136, 1921–1930. [Google Scholar] [CrossRef] [PubMed]
- Lagergren, J.; Smyth, E.; Cunningham, D.; Lagergren, P. Oesophageal cancer. Lancet 2017, 390, 2383–2396. [Google Scholar] [CrossRef]
- Neal, C.L.; Yu, D. 14-3-3zeta as a prognostic marker and therapeutic target for cancer. Expert. Opin. Ther. Targets 2010, 14, 1343–1354. [Google Scholar] [CrossRef] [PubMed]
- Perez, S.M.; Brinton, L.T.; Kelly, K.A. Plectin in Cancer: From Biomarker to Therapeutic Target. Cells 2021, 10, 2246. [Google Scholar] [CrossRef]
- Sanchez-Danes, A.; Blanpain, C. Deciphering the cells of origin of squamous cell carcinomas. Nat. Rev. Cancer 2018, 18, 549–561. [Google Scholar] [CrossRef]
- Song, Y.; Li, L.; Ou, Y.; Gao, Z.; Li, E.; Li, X.; Zhang, W.; Wang, J.; Xu, L.; Zhou, Y.; et al. Identification of genomic alterations in oesophageal squamous cell cancer. Nature 2014, 509, 91–95. [Google Scholar] [CrossRef]
- Rodriguez, H.; Zenklusen, J.C.; Staudt, L.M.; Doroshow, J.H.; Lowy, D.R. The next horizon in precision oncology: Proteogenomics to inform cancer diagnosis and treatment. Cell 2021, 184, 1661–1670. [Google Scholar] [CrossRef]
- Liu, W.; Xie, L.; He, Y.H.; Wu, Z.Y.; Liu, L.X.; Bai, X.F.; Deng, D.X.; Xu, X.E.; Liao, L.D.; Lin, W.; et al. Large-scale and high-resolution mass spectrometry-based proteomics profiling defines molecular subtypes of esophageal cancer for therapeutic targeting. Nat. Commun. 2021, 12, 4961. [Google Scholar] [CrossRef]
- Moldovan, G.L.; Pfander, B.; Jentsch, S. PCNA, the maestro of the replication fork. Cell 2007, 129, 665–679. [Google Scholar] [CrossRef] [PubMed]
- Mailand, N.; Gibbs-Seymour, I.; Bekker-Jensen, S. Regulation of PCNA-protein interactions for genome stability. Nat. Rev. Mol. Cell Biol. 2013, 14, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Sykaras, A.G.; Pergaris, A.; Theocharis, S. Challenging, Accurate and Feasible: CAF-1 as a Tumour Proliferation Marker of Diagnostic and Prognostic Value. Cancers 2021, 13, 2575. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Zhang, X.; Wang, Y.; Gan, H.; Xu, X.; Lv, X.; Hua, X.; Que, J.; Ordog, T.; Zhang, Z. Chromatin Assembly Factor 1 (CAF-1) facilitates the establishment of facultative heterochromatin during pluripotency exit. Nucleic Acids Res. 2019, 47, 11114–11131. [Google Scholar] [CrossRef] [PubMed]
- Gerard, A.; Koundrioukoff, S.; Ramillon, V.; Sergere, J.C.; Mailand, N.; Quivy, J.P.; Almouzni, G. The replication kinase Cdc7-Dbf4 promotes the interaction of the p150 subunit of chromatin assembly factor 1 with proliferating cell nuclear antigen. EMBO Rep. 2006, 7, 817–823. [Google Scholar] [CrossRef]
- Shimada, Y.; Imamura, M.; Wagata, T.; Yamaguchi, N.; Tobe, T. Characterization of 21 newly established esophageal cancer cell lines. Cancer 1992, 69, 277–284. [Google Scholar] [CrossRef]
- Nishihira, T.; Hashimoto, Y.; Katayama, M.; Mori, S.; Kuroki, T. Molecular and cellular features of esophageal cancer cells. J. Cancer Res. Clin. Oncol. 1993, 119, 441–449. [Google Scholar] [CrossRef]
- Long, L.; He, J.Z.; Chen, Y.; Xu, X.E.; Liao, L.D.; Xie, Y.M.; Li, E.M.; Xu, L.Y. Riboflavin Depletion Promotes Tumorigenesis in HEK293T and NIH3T3 Cells by Sustaining Cell Proliferation and Regulating Cell Cycle-Related Gene Transcription. J. Nutr. 2018, 148, 834–843. [Google Scholar] [CrossRef]
- Schwab, R.A.; Blackford, A.N.; Niedzwiedz, W. ATR activation and replication fork restart are defective in FANCM-deficient cells. EMBO J. 2010, 29, 806–818. [Google Scholar] [CrossRef]
- Przetocka, S.; Porro, A.; Bolck, H.A.; Walker, C.; Lezaja, A.; Trenner, A.; von Aesch, C.; Himmels, S.F.; D’Andrea, A.D.; Ceccaldi, R.; et al. CtIP-Mediated Fork Protection Synergizes with BRCA1 to Suppress Genomic Instability upon DNA Replication Stress. Mol. Cell 2018, 72, 568–582.e6. [Google Scholar] [CrossRef] [PubMed]
- Quinet, A.; Tirman, S.; Cybulla, E.; Meroni, A.; Vindigni, A. To skip or not to skip: Choosing repriming to tolerate DNA damage. Mol. Cell 2021, 81, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.C.; Hao, J.J.; Nagata, Y.; Xu, L.; Shang, L.; Meng, X.; Sato, Y.; Okuno, Y.; Varela, A.M.; Ding, L.W.; et al. Genomic and molecular characterization of esophageal squamous cell carcinoma. Nat. Genet. 2014, 46, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Research Network Analysis Working Group; Kim, J.; Bowlby, R.; Mungall, A.J.; Robertson, A.G.; Odze, R.D.; Cherniack, A.D.; Shih, J.; Pedamallu, C.S.; Cibulskis, C.; et al. Integrated genomic characterization of oesophageal carcinoma. Nature 2017, 541, 169–175. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, Y.; Cheng, C.; Cui, H.; Cheng, L.; Kong, P.; Wang, J.; Li, Y.; Chen, W.; Song, B.; et al. Genomic analyses reveal mutational signatures and frequently altered genes in esophageal squamous cell carcinoma. Am. J. Hum. Genet. 2015, 96, 597–611. [Google Scholar] [CrossRef]
- Cui, Y.; Chen, H.; Xi, R.; Cui, H.; Zhao, Y.; Xu, E.; Yan, T.; Lu, X.; Huang, F.; Kong, P.; et al. Whole-genome sequencing of 508 patients identifies key molecular features associated with poor prognosis in esophageal squamous cell carcinoma. Cell Res. 2020, 30, 902–913. [Google Scholar] [CrossRef]
- Matthews, H.K.; Bertoli, C.; de Bruin, R.A.M. Cell cycle control in cancer. Nat. Rev. Mol. Cell Biol. 2022, 23, 74–88. [Google Scholar] [CrossRef]
- Techer, H.; Koundrioukoff, S.; Nicolas, A.; Debatisse, M. The impact of replication stress on replication dynamics and DNA damage in vertebrate cells. Nat. Rev. Genet. 2017, 18, 535–550. [Google Scholar] [CrossRef]
- Hong, L.; Han, Y.; Zhang, H.; Fan, D. Prognostic markers in esophageal cancer: From basic research to clinical use. Expert. Rev. Gastroenterol. Hepatol. 2015, 9, 887–889. [Google Scholar] [CrossRef]
- Yang, Y.M.; Hong, P.; Xu, W.W.; He, Q.Y.; Li, B. Advances in targeted therapy for esophageal cancer. Signal Transduct. Target. Ther. 2020, 5, 229. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Hofstetter, W.L. Prognostic and therapeutic molecular markers in the clinical management of esophageal cancer. Expert. Rev. Mol. Diagn. 2020, 20, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Sauer, P.V.; Gu, Y.; Liu, W.H.; Mattiroli, F.; Panne, D.; Luger, K.; Churchill, M.E. Mechanistic insights into histone deposition and nucleosome assembly by the chromatin assembly factor-1. Nucleic Acids Res. 2018, 46, 9907–9917. [Google Scholar] [CrossRef]
- Ng, C.; Aichinger, M.; Nguyen, T.; Au, C.; Najar, T.; Wu, L.; Mesa, K.R.; Liao, W.; Quivy, J.P.; Hubert, B.; et al. The histone chaperone CAF-1 cooperates with the DNA methyltransferases to maintain Cd4 silencing in cytotoxic T cells. Genes Dev. 2019, 33, 669–683. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibodies (Dilution Factor) | Source | Identifier |
|---|---|---|
| Mouse monoclonal anti-PCNA (1:2000) | ZEN BIO | Cat#200947-2E1 |
| Rabbit polyclonal anti-CHAF1A (1:1000) | Proteintech | Cat#17037-1-AP |
| Mouse monoclonal anti-Lamin B1 (1:10,000) | Proteintech | Cat#66095-1-Ig |
| HRP-conjugated monoclonal GAPDH (1:20,000) | Proteintech | Cat#HRP-60004 |
| HRP-conjugated monoclonal alpha tubulin (1:10,000) | Proteintech | Cat#HRP-66031 |
| Goat anti-mouse IgG-HRP (1:5000) | Santa Cruz Biotechnology | Cat#sc-2005 |
| Goat anti-rabbit IgG-HRP (1:5000) | Santa Cruz Biotechnology | Cat#sc-2030 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, B.; Deng, D.-X.; Liao, L.-D.; Zhang, Z.-D.; Zheng, Y.-Q.; Dong, K.; Xu, L.-Y.; Li, E.-M. Co-Expression of Chromatin Assembly Factor 1 Subunit A and Proliferating Cell Nuclear Antigen Is a Prognostic Biomarker of Esophageal Cancer. Biomedicines 2023, 11, 1184. https://doi.org/10.3390/biomedicines11041184
Wen B, Deng D-X, Liao L-D, Zhang Z-D, Zheng Y-Q, Dong K, Xu L-Y, Li E-M. Co-Expression of Chromatin Assembly Factor 1 Subunit A and Proliferating Cell Nuclear Antigen Is a Prognostic Biomarker of Esophageal Cancer. Biomedicines. 2023; 11(4):1184. https://doi.org/10.3390/biomedicines11041184
Chicago/Turabian StyleWen, Bing, Dan-Xia Deng, Lian-Di Liao, Zhi-Da Zhang, Ya-Qi Zheng, Ke Dong, Li-Yan Xu, and En-Min Li. 2023. "Co-Expression of Chromatin Assembly Factor 1 Subunit A and Proliferating Cell Nuclear Antigen Is a Prognostic Biomarker of Esophageal Cancer" Biomedicines 11, no. 4: 1184. https://doi.org/10.3390/biomedicines11041184
APA StyleWen, B., Deng, D.-X., Liao, L.-D., Zhang, Z.-D., Zheng, Y.-Q., Dong, K., Xu, L.-Y., & Li, E.-M. (2023). Co-Expression of Chromatin Assembly Factor 1 Subunit A and Proliferating Cell Nuclear Antigen Is a Prognostic Biomarker of Esophageal Cancer. Biomedicines, 11(4), 1184. https://doi.org/10.3390/biomedicines11041184