
New Insights on Saporin Resistance to Chemical Derivatization with Heterobifunctional Reagents

 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cells
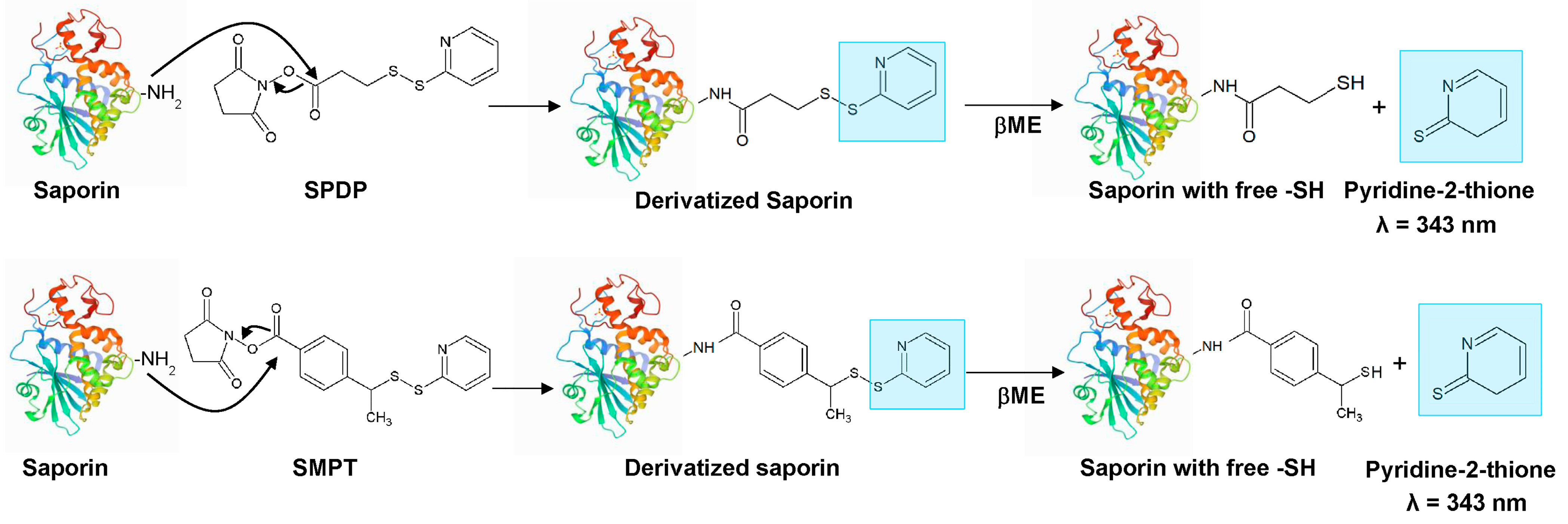
2.3. Derivatization of Saporin with 2-IT
2.4. Derivatization of Saporin with SPDP and SMPT
2.5. Cell-Free Protein Synthesis Inhibition Assay
2.6. Polynucleotide: Adenosine Glycosylase (PNAG) Activity on Herring Sperm DNA Assay
2.7. Cell Viability
2.8. Statistical Analysis
3. Results
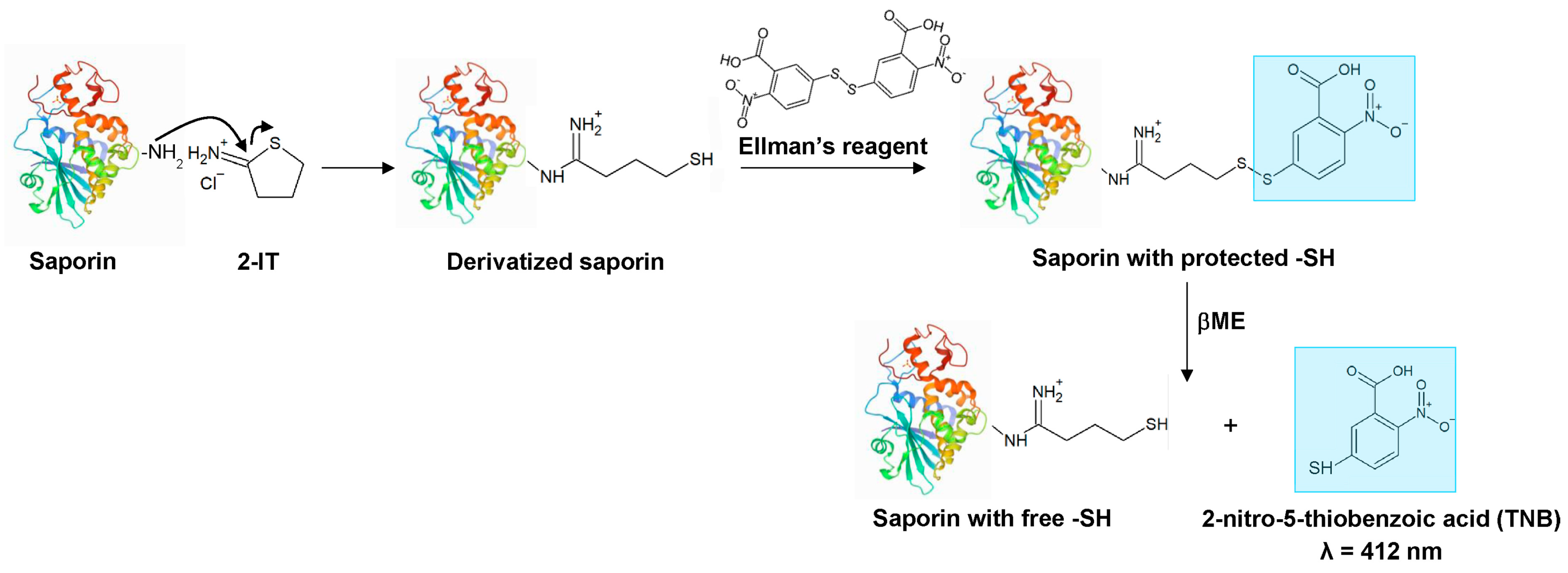
3.1. Derivatization of Saporin
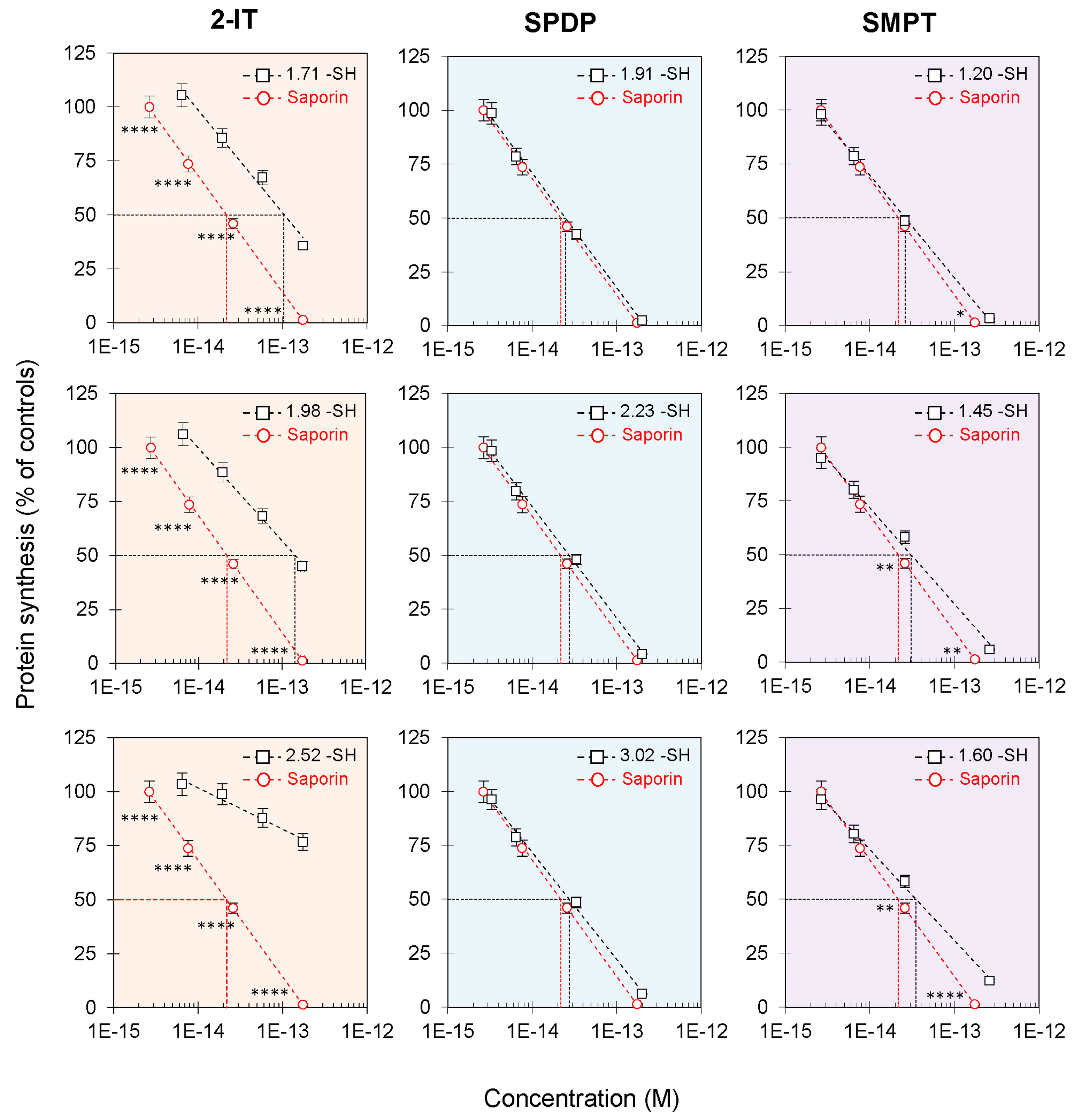
3.2. Effect of Derivatized Saporin on Cell-Free Protein Synthesis Inhibition
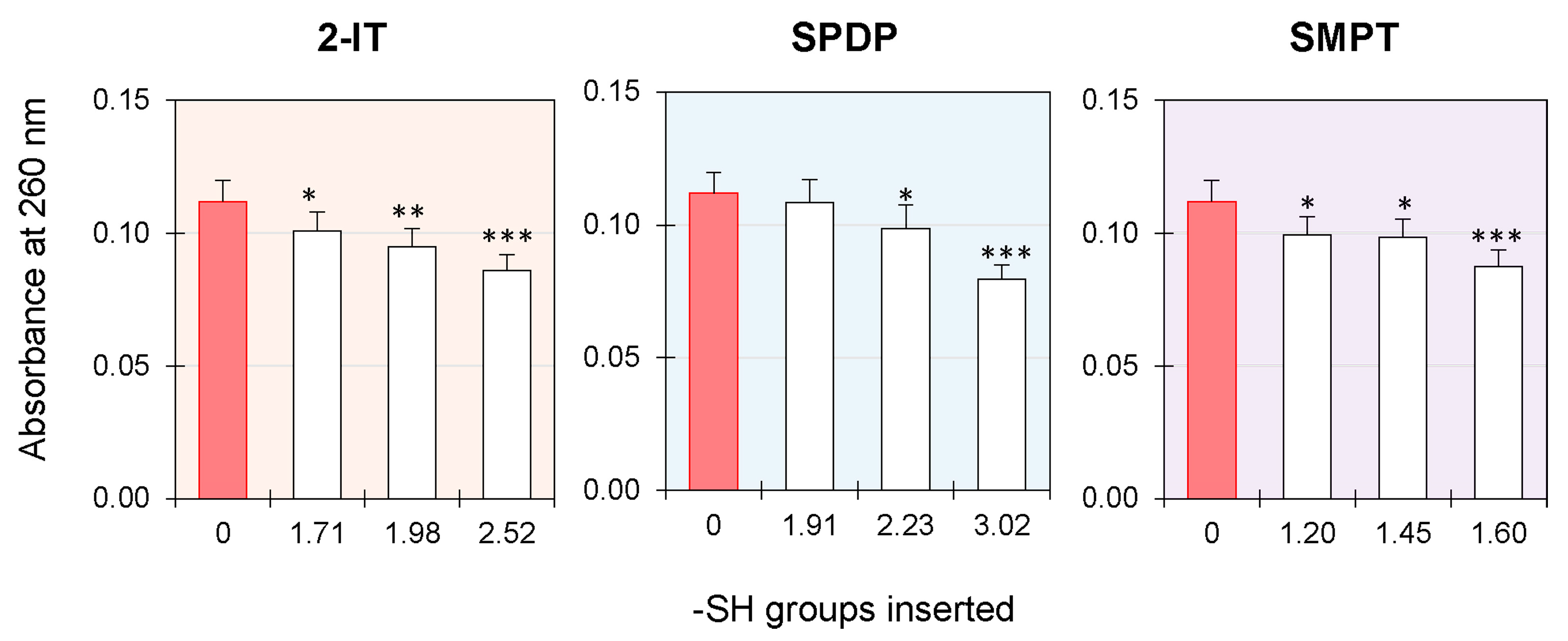
3.3. Effect of Derivatized Saporin on Polynucleotide:Adenosine Glycosylase (PNAG) Activity
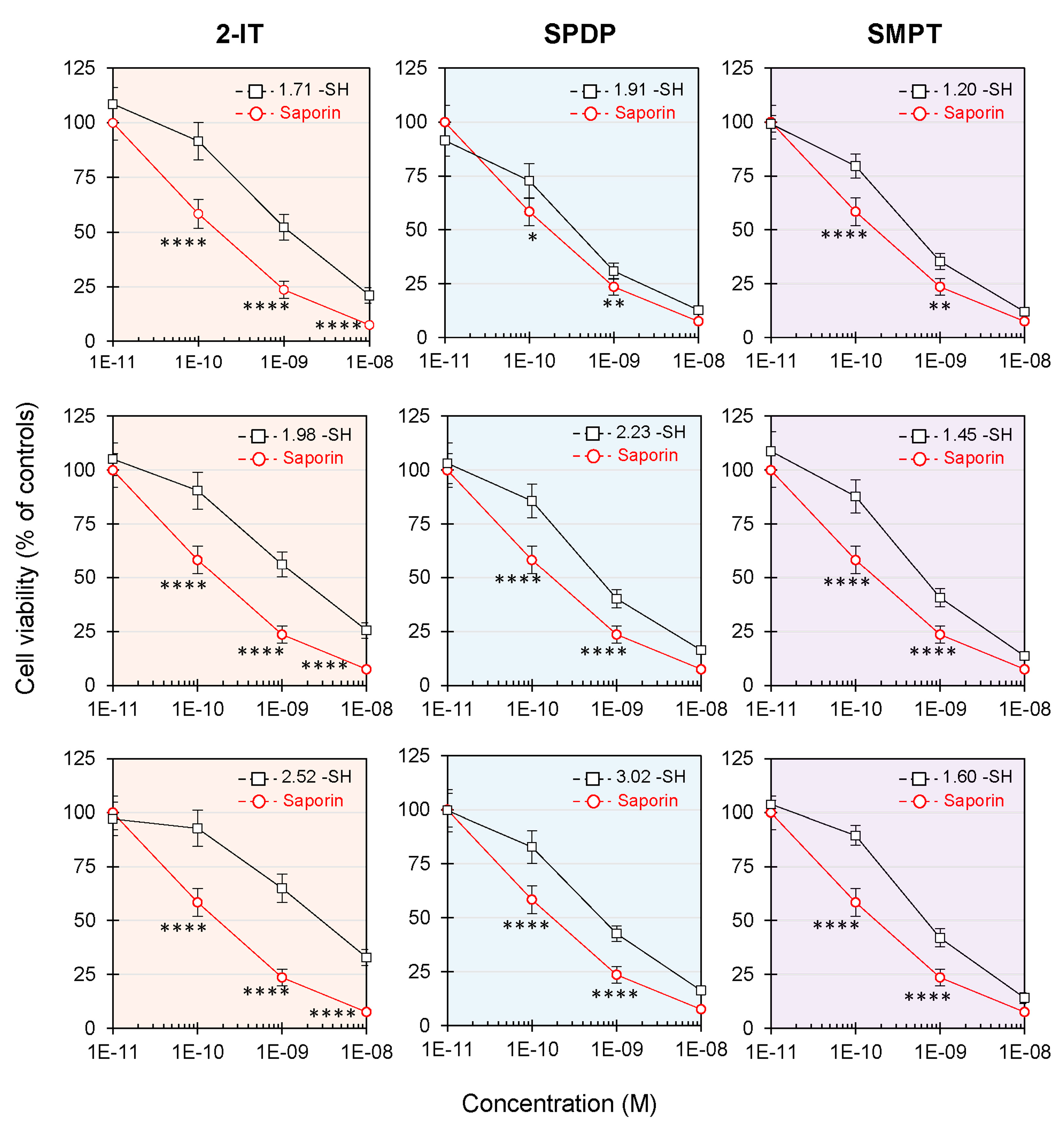
3.4. Effect of Derivatized Saporin on NB100 Cell Viability
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Walsh, M.J.; Dodd, J.E.; Hautbergue, G.M. Ribosome-inactivating proteins: Potent poisons and molecular tools. Virulence 2013, 4, 774–784. [Google Scholar] [CrossRef]
- Barbieri, L.; Battelli, M.G.; Stirpe, F. Ribosome-inactivating proteins from plants. Biochim. Biophys. Acta 1993, 1154, 237–282. [Google Scholar] [CrossRef]
- Dougherty, K.; Hudak, K.A. Phylogeny and domain architecture of plant ribosome inactivating proteins. Phytochemistry 2022, 202, 113337. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Gupta, S.; Sharma, N.R.; Paul, K. Expanding role of ribosome-inactivating proteins: From toxins to therapeutics. IUBMB Life 2023, 75, 82–96. [Google Scholar] [CrossRef]
- Bolognesi, A.; Bortolotti, M.; Maiello, S.; Battelli, M.G.; Polito, L. Ribosome-Inactivating Proteins from Plants: A Historical Overview. Molecules 2016, 21, 1627. [Google Scholar] [CrossRef] [PubMed]
- Bolognesi, A.; Polito, L.; Lubelli, C.; Barbieri, L.; Parente, A.; Stirpe, F. Ribosome-inactivating and adenine polynucleotide glycosylase activities in Mirabilis jalapa L. tissues. J. Biol. Chem. 2002, 277, 13709–13716. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, L.; Valbonesi, P.; Bonora, E.; Gorini, P.; Bolognesi, A.; Stirpe, F. Polynucleotide:adenosine glycosidase activity of ribosome-inactivating proteins: Effect on DNA, RNA and poly(A). Nucleic Acids Res. 1997, 25, 518–522. [Google Scholar] [CrossRef]
- Polito, L.; Bortolotti, M.; Farini, V.; Battelli, M.G.; Barbieri, L.; Bolognesi, A. Saporin induces multiple death pathways in lymphoma cells with different intensity and timing as compared to ricin. Int. J. Biochem. Cell Biol. 2009, 41, 1055–1061. [Google Scholar] [CrossRef]
- Polito, L.; Mercatelli, D.; Bortolotti, M.; Maiello, S.; Djemil, A.; Battelli, M.G.; Bolognesi, A. Two Saporin-Containing Immunotoxins Specific for CD20 and CD22 Show Different Behavior in Killing Lymphoma Cells. Toxins 2017, 9, 182. [Google Scholar] [CrossRef] [PubMed]
- Polito, L.; Djemil, A.; Bortolotti, M. Plant Toxin-Based Immunotoxins for Cancer Therapy: A Short Overview. Biomedicines 2016, 4, 12. [Google Scholar] [CrossRef]
- Polito, L.; Calafato, G.; Bortolotti, M.; Chiarelli Olivari, C.; Maiello, S.; Bolognesi, A. Antibody Conjugates for Sarcoma Therapy: How Far along Are We? Biomedicines 2021, 9, 978. [Google Scholar] [CrossRef] [PubMed]
- Stoessel, A.; Groysbeck, N.; Guyot, L.; Barret, L.; Nominé, Y.; Nguekeu-Zebaze, L.; Bender, A.; Voilquin, L.; Lutz, T.; Pallaoro, N.; et al. Modular Conjugation of a Potent Anti-HER2 Immunotoxin Using Coassociating Peptides. Bioconjug. Chem. 2020, 31, 2421–2430. [Google Scholar] [CrossRef] [PubMed]
- Flavell, D.J.; Flavell, S.U. Plant-Derived Type I Ribosome Inactivating Protein-Based Targeted Toxins: A Review of the Clinical Experience. Toxins 2022, 14, 563. [Google Scholar] [CrossRef]
- Polito, L.; Bortolotti, M.; Mercatelli, D.; Battelli, M.G.; Bolognesi, A. Saporin-S6: A useful tool in cancer therapy. Toxins 2013, 5, 1698–1722. [Google Scholar] [CrossRef] [PubMed]
- Giansanti, F.; Flavell, D.J.; Angelucci, F.; Fabbrini, M.S.; Ippoliti, R. Strategies to Improve the Clinical Utility of Saporin-Based Targeted Toxins. Toxins 2018, 10, 82. [Google Scholar] [CrossRef]
- Wiley, R.G.; Lappi, D.A. Targeted toxins in pain. Adv. Drug Deliv. Rev. 2003, 55, 1043–1054. [Google Scholar] [CrossRef]
- Ancheta, L.R.; Shramm, P.A.; Bouajram, R.; Higgins, D.; Lappi, D.A. Saporin as a Commercial Reagent: Its Uses and Unexpected Impacts in the Biological Sciences-Tools from the Plant Kingdom. Toxins 2022, 14, 184. [Google Scholar] [CrossRef]
- Stirpe, F.; Gasperi-Campani, A.; Barbieri, L.; Falasca, A.; Abbondanza, A.; Stevens, W.A. Ribosome-inactivating proteins from the seeds of Saponaria officinalis L. (soapwort), of Agrostemma githago L. (corn cockle) and of Asparagus officinalis L. (asparagus), and from the latex of Hura crepitans L. (sandbox tree). Biochem. J. 1983, 216, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Lord, J.M.; Spooner, R.A. Ricin trafficking in plant and mammalian cells. Toxins 2011, 3, 787–801. [Google Scholar] [CrossRef]
- Su, Z.; Xiao, D.; Xie, F.; Liu, L.; Wang, Y.; Fan, S.; Zhou, X.; Li, S. Antibody-drug conjugates: Recent advances in linker chemistry. Acta Pharm. Sin. B 2021, 11, 3889–3907. [Google Scholar] [CrossRef] [PubMed]
- King, T.P.; Li, Y.; Kochoumian, L. Preparation of protein conjugates via intermolecular disulfide bond formation. Biochemistry 1978, 17, 1499–1506. [Google Scholar] [CrossRef] [PubMed]
- Jue, R.; Lambert, J.M.; Pierce, L.R.; Traut, R.R. Addition of sulfhydryl groups to Escherichia coli ribosomes by protein modification with 2-iminothiolane (methyl 4-mercaptobutyrimidate). Biochemistry 1978, 17, 5399–5406. [Google Scholar] [CrossRef]
- Bolognesi, A.; Barbieri, L.; Abbondanza, A.; Falasca, A.I.; Carnicelli, D.; Battelli, M.G.; Stirpe, F. Purification and properties of new ribosome-inactivating proteins with RNA N-glycosidase activity. Biochim. Biophys. Acta 1990, 1087, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, P.E.; Wallace, P.M.; Knowles, P.P.; Relf, M.G.; Brown, A.N.; Watson, G.J.; Knyba, R.E.; Wawrzynczak, E.J.; Blakey, D.C. New coupling agents for the synthesis of immunotoxins containing a hindered disulfide bond with improved stability in vivo. Cancer Res. 1987, 47, 5924–5931. [Google Scholar]
- Flavell, D.J.; Boehm, D.A.; Okayama, K.; Kohler, J.A.; Flavell, S.U. Therapy of human T-cell acute lymphoblastic leukaemia in severe combined immunodeficient mice with two different anti-CD7-saporin immunotoxins containing hindered or non-hindered disulphide cross-linkers. Int. J. Cancer 1994, 58, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Manjappa, A.S.; Chaudhari, K.R.; Venkataraju, M.P.; Dantuluri, P.; Nanda, B.; Sidda, C.; Sawant, K.K.; Murthy, R.S. Antibody derivatization and conjugation strategies: Application in preparation of stealth immunoliposome to target chemotherapeutics to tumor. J. Control Release 2011, 150, 2–22. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, L.; Stoppa, C.; Bolognesi, A. Large scale chromatographic purification of ribosome-inactivating proteins. J. Chromatogr. 1987, 408, 235–243. [Google Scholar] [CrossRef]
- Ferreras, J.M.; Barbieri, L.; Girbés, T.; Battelli, M.G.; Rojo, M.A.; Arias, F.J.; Rocher, M.A.; Soriano, F.; Mendéz, E.; Stirpe, F. Distribution and properties of major ribosome-inactivating proteins (28 S rRNA N-glycosidases) of the plant Saponaria officinalis L. (Caryophyllaceae). Biochim. Biophys. Acta 1993, 1216, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Polito, L.; Bortolotti, M.; Pedrazzi, M.; Mercatelli, D.; Battelli, M.G.; Bolognesi, A. Apoptosis and necroptosis induced by stenodactylin in neuroblastoma cells can be completely prevented through caspase inhibition plus catalase or necrostatin-1. Phytomedicine 2016, 23, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Eyer, P.; Worek, F.; Kiderlen, D.; Sinko, G.; Stuglin, A.; Simeon-Rudolf, V.; Reiner, E. Molar absorption coefficients for the reduced Ellman reagent: Reassessment. Anal. Biochem. 2003, 312, 224–227. [Google Scholar] [CrossRef]
- Carlsson, J.; Drevin, H.; Axén, R. Protein thiolation and reversible protein-protein conjugation: N-succynimidyl 3-(2-pyridyldithio)propionate, a new heterobifunctional reagent. Biochem. J. 1978, 173, 723–737. [Google Scholar] [CrossRef] [PubMed]
- Polito, L.; Bolognesi, A.; Tazzari, P.L.; Farini, V.; Lubelli, C.; Zinzani, P.L.; Ricci, F.; Stirpe, F. The conjugate Rituximab/saporin-S6 completely inhibits clonogenic growth of CD20-expressing cells and produces a synergistic toxic effect with Fludarabine. Leukemia 2004, 18, 1215–1222. [Google Scholar] [CrossRef] [PubMed]
- Bortolotti, M.; Maiello, S.; Ferreras, J.M.; Iglesias, R.; Polito, L.; Bolognesi, A. Kirkiin: A New Toxic Type 2 Ribosome-Inactivating Protein from the Caudex of Adenia kirkii. Toxins 2021, 13, 81. [Google Scholar] [CrossRef]
- Stirpe, F.; Bolognesi, A.; Bortolotti, M.; Farini, V.; Lubelli, C.; Pelosi, E.; Polito, L.; Dozza, B.; Strocchi, P.; Chambery, A.; et al. Characterization of highly toxic type 2 ribosome-inactivating proteins from Adenia lanceolata and Adenia stenodactyla (Passifloraceae). Toxicon 2007, 50, 94–105. [Google Scholar] [CrossRef]
- Polito, L.; Bortolotti, M.; Pedrazzi, M.; Bolognesi, A. Immunotoxins and other conjugates containing saporin-S6 for cancer therapy. Toxins 2011, 3, 697–720. [Google Scholar] [CrossRef] [PubMed]
- Holmes, S.E.; Bachran, C.; Fuchs, H.; Weng, A.; Melzig, M.F.; Flavell, S.U.; Flavell, D.J. Triterpenoid saponin augmention of saporin-based immunotoxin cytotoxicity for human leukaemia and lymphoma cells is partially immunospecific and target molecule dependent. Immunopharmacol. Immunotoxicol. 2015, 37, 42–55. [Google Scholar] [CrossRef]
- Savino, C.; Federici, L.; Brancaccio, A.; Ippoliti, R.; Lendaro, E.; Tsernoglou, D. Crystallization and preliminary X-ray study of saporin, a ribosome-inactivating protein from Saponaria officinalis. Acta Crystallogr. D Biol. Crystallogr. 1998, 54, 636–638. [Google Scholar] [CrossRef]
- Savino, C.; Federici, L.; Ippoliti, R.; Lendaro, E.; Tsernoglou, D. The crystal structure of saporin SO6 from Saponaria officinalis and its interaction with the ribosome. FEBS Lett. 2000, 470, 239–243. [Google Scholar] [CrossRef]
- Santanché, S.; Bellelli, A.; Brunori, M. The unusual stability of saporin, a candidate for the synthesis of immunotoxins. Biochem. Biophys. Res. Commun. 1997, 234, 129–132. [Google Scholar] [CrossRef]
- Bortolotti, M.; Bolognesi, A.; Battelli, M.G.; Polito, L. High in vitro anti-tumor efficacy of dimeric Rituximab/Saporin-S6 immunotoxin. Toxins 2016, 8, 192. [Google Scholar] [CrossRef]
- Pasqualucci, L.; Wasik, M.; Teicher, B.A.; Flenghi, L.; Bolognesi, A.; Stirpe, F.; Polito, L.; Falini, B.; Kadin, M.E. Antitumor activity of anti-CD30 immunotoxin (Ber-H2/saporin) in vitro and in severe combined immunodeficiency disease mice xenografted with human CD30+ anaplastic large-cell lymphoma. Blood 1995, 85, 2139–2146. [Google Scholar] [CrossRef] [PubMed]
- Bolognesi, A.; Tazzari, P.L.; Tassi, C.; Gromo, G.; Gobbi, M.; Stirpe, F. A comparison of anti-lymphocyte immunotoxins containing different ribosome-inactivating proteins and antibodies. Clin. Exp. Immunol. 1992, 89, 341–346. [Google Scholar] [CrossRef]
- Ricci, C.; Polito, L.; Nanni, P.; Landuzzi, L.; Astolfi, A.; Nicoletti, G.; Rossi, I.; De Giovanni, C.; Bolognesi, A.; Lollini, P.L. HER/erbB receptors as therapeutic targets of immunotoxins in human rhabdomyosarcoma cells. J. Immunother. 2002, 25, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Polito, L.; Bortolotti, M.; Farini, V.; Pedrazzi, M.; Tazzari, P.L.; Bolognesi, A. ATG-saporin-S6 immunotoxin: A new potent and selective drug to eliminate activated lymphocytes and lymphoma cells. Br. J. Haematol. 2009, 147, 710–718. [Google Scholar] [CrossRef]
- Schrot, J.; Weng, A.; Melzig, M.F. Ribosome-inactivating and related proteins. Toxins 2015, 7, 1556–1615. [Google Scholar] [CrossRef] [PubMed]
- Fermani, S.; Tosi, G.; Farini, V.; Polito, L.; Falini, G.; Ripamonti, A.; Barbieri, L.; Chambery, A.; Bolognesi, A. Structure/function studies on two type 1 ribosome inactivating proteins: Bouganin and lychnin. J. Struct. Biol. 2009, 168, 278–287. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Calculation Scheme | Formula |
|---|---|
| 1—Concentration of inserted -SH groups ([-SH]) | (A412 (+βME) − A412 (−βME))/ε412 (TNB) |
| 2—Contribution of A280 of TNB (A280 (TNB)) | [-SH] × ε280 (TNB) |
| 3—Concentration of saporin ([saporin]) | (A280 (saporin) − A280 (TNB))/ε280 (saporin) |
| 4—Derivatization ratio | [-SH]/[saporin] |
| Calculation Scheme | Formula |
|---|---|
| 1—Concentration of inserted -SH groups ([-SH]) | (A343 (+βME) − A343 (−βME))/ε343 (pyridine-2-thione) |
| 2—Contribution of A280 of pyridine-2-thione (A280 (pyridine-2-thione)) | [-SH] × ε280 (pyridine-2-thione) |
| 3—Concentration of saporin ([saporin]) | (A280 (saporin) − A280 (pyridine-2-thione))/ε280 (saporin) |
| 4—Derivatization ratio | [-SH]/[saporin] |
| Concentration (mM) | Reagent/Saporin (mol/mol) | -SH/Saporin Inserted 1 | |
|---|---|---|---|
| 2-IT | 1.75 | 8 | 1.71 |
| 2 | 9 | 1.98 | |
| 2.5 | 11 | 2.52 | |
| SPDP | 0.48 | 2 | 1.91 |
| 0.57 | 2.5 | 2.23 | |
| 0.71 | 3 | 3.02 | |
| SMPT | 0.48 | 2 | 1.20 |
| 0.57 | 2.5 | 1.45 | |
| 0.71 | 3 | 1.60 |
| -SH/Saporin Inserted 1 | IC50 2 (pM) | PNAG Activity Ratio 3 (%) | EC50 4 (pM) | |
|---|---|---|---|---|
| Saporin | --- | 22 | 100 | 259 |
| 2-IT | 1.71 | 103 | 90.2 | 1280 |
| 1.98 | 143 | 84.8 | 1620 | |
| 2.52 | >173 | 76.8 | 2840 | |
| SPDP | 1.91 | 25 | 97.3 | 375 |
| 2.23 | 27 | 88.4 | 718 | |
| 3.02 | 28 | 71.4 | 740 | |
| SMPT | 1.20 | 26 | 88.4 | 511 |
| 1.45 | 31 | 88.4 | 763 | |
| 1.60 | 35 | 76.8 | 774 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bortolotti, M.; Biscotti, F.; Zanello, A.; Bolognesi, A.; Polito, L. New Insights on Saporin Resistance to Chemical Derivatization with Heterobifunctional Reagents. Biomedicines 2023, 11, 1214. https://doi.org/10.3390/biomedicines11041214
Bortolotti M, Biscotti F, Zanello A, Bolognesi A, Polito L. New Insights on Saporin Resistance to Chemical Derivatization with Heterobifunctional Reagents. Biomedicines. 2023; 11(4):1214. https://doi.org/10.3390/biomedicines11041214
Chicago/Turabian StyleBortolotti, Massimo, Francesco Biscotti, Andrea Zanello, Andrea Bolognesi, and Letizia Polito. 2023. "New Insights on Saporin Resistance to Chemical Derivatization with Heterobifunctional Reagents" Biomedicines 11, no. 4: 1214. https://doi.org/10.3390/biomedicines11041214
APA StyleBortolotti, M., Biscotti, F., Zanello, A., Bolognesi, A., & Polito, L. (2023). New Insights on Saporin Resistance to Chemical Derivatization with Heterobifunctional Reagents. Biomedicines, 11(4), 1214. https://doi.org/10.3390/biomedicines11041214