

Age-Related Effects of Exogenous Melatonin on Anxiety-like Behavior in C57/B6J Mice

 and
and 
Abstract
:
1. Introduction
2. Materials and Methods
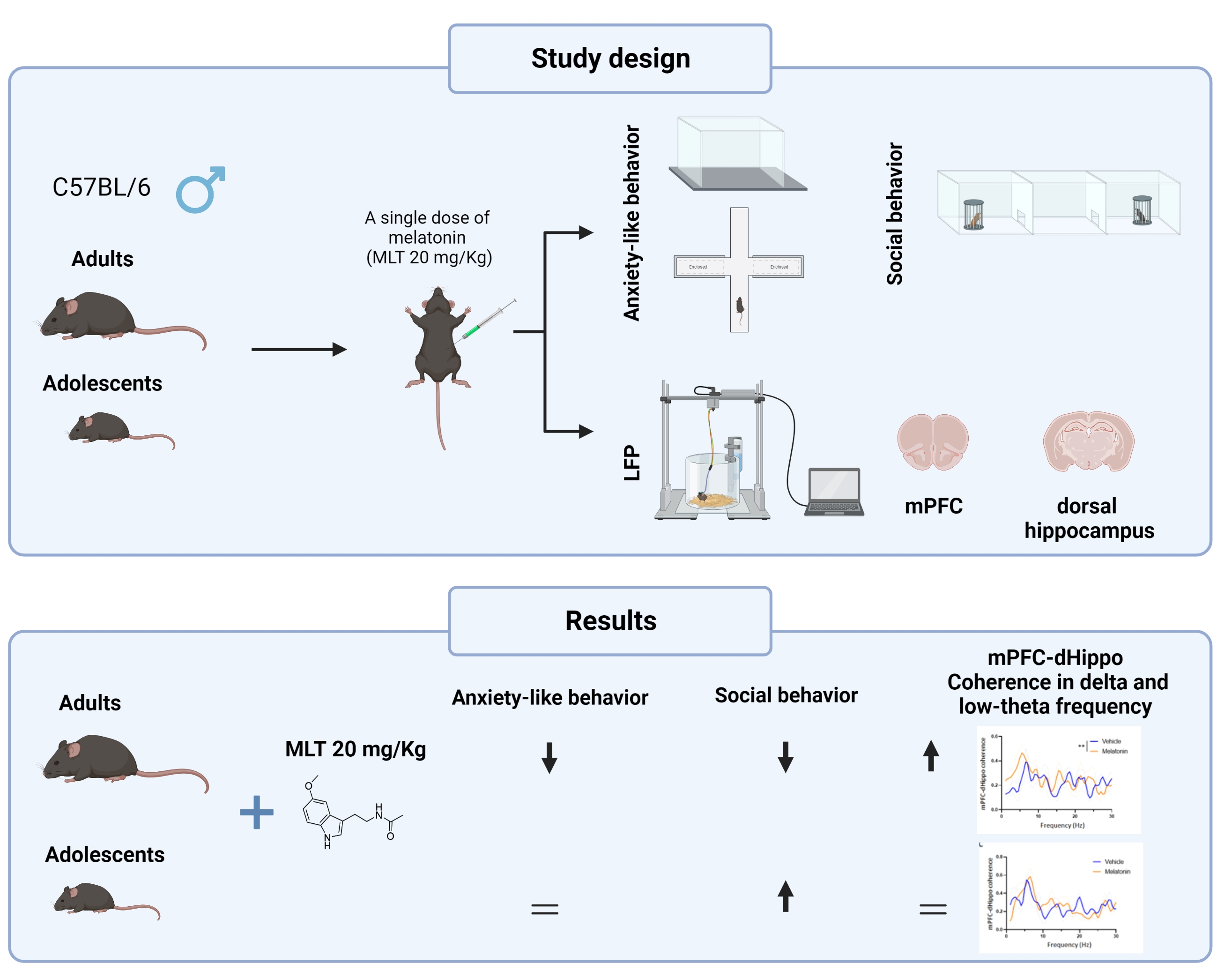
2.1. Experimental Design
2.2. Animals
2.3. Treatment
2.4. Behavioral Testing
2.4.1. Open-Field Test (OFT)
2.4.2. Elevated Plus Maze Test (EPMT)
2.4.3. Three-Chamber Sociability Test
2.4.4. Marble Burying Test
2.5. In Vivo LFP Recordings and Analysis
2.6. Statistical Analysis
3. Results
3.1. Adolescent Mice
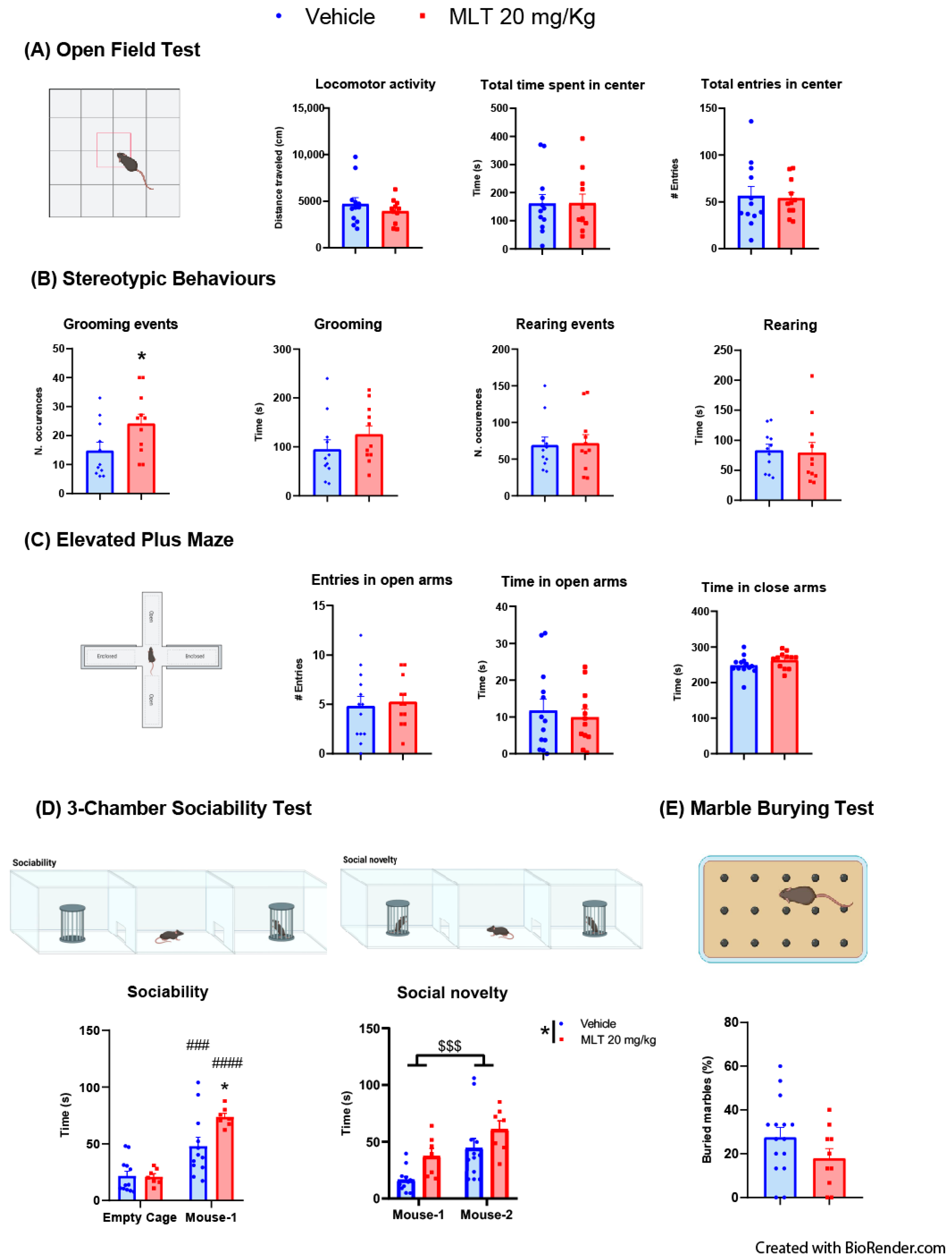
3.1.1. Evaluation of the Effects of MLT on Anxiety-like Behaviors: OFT, Stereotypic Behaviors and EPMT
3.1.2. Evaluation of the Effects of MLT on Sociability: Three-Chambers Sociability Test
3.1.3. Evaluation of the Effects of MLT on Obsessive–Compulsive Disorder (OCD) Behavior: Marble Burying Test
3.1.4. In Vivo Electrophysiology
3.2. Adult Mice
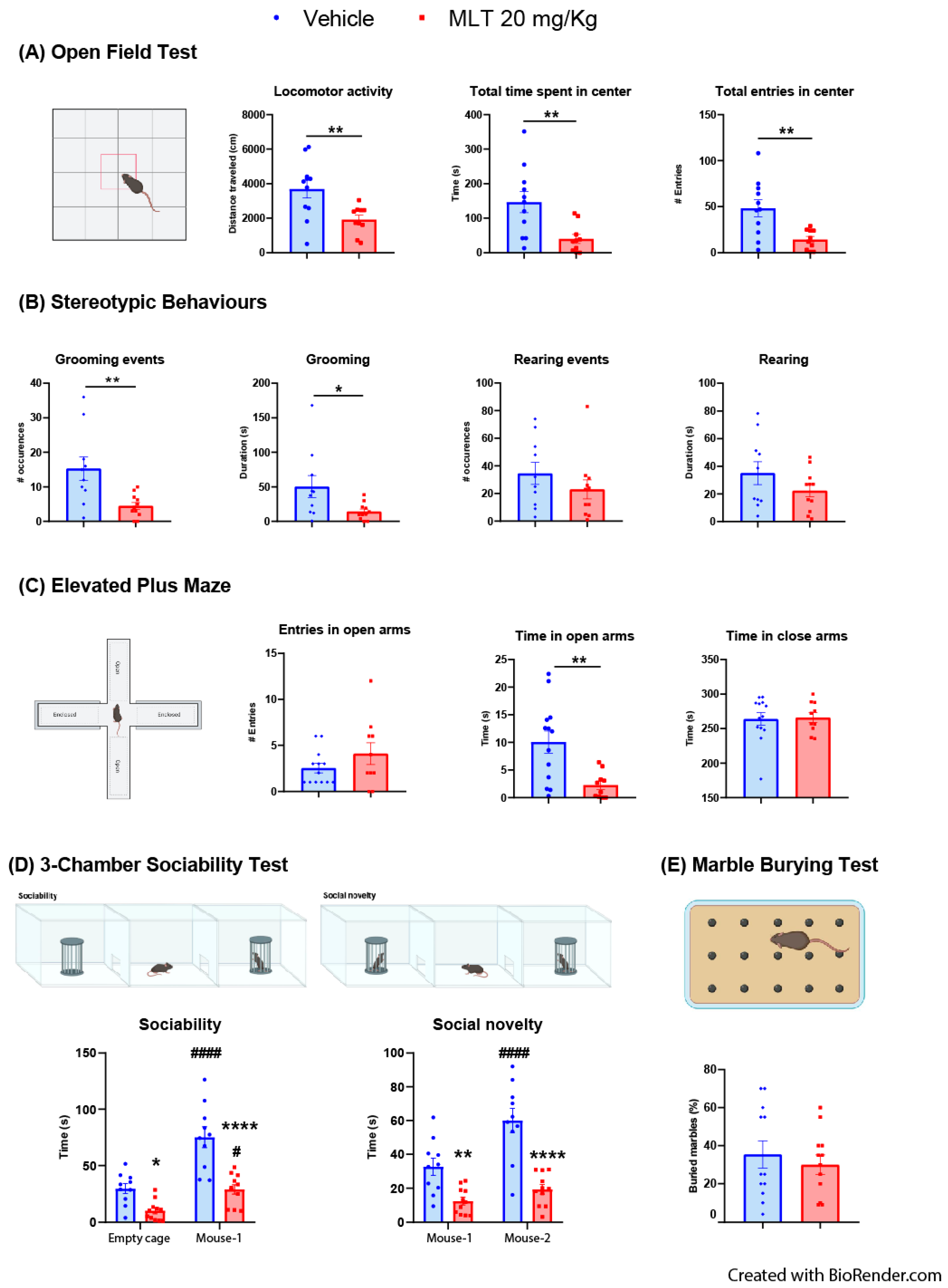
3.2.1. Evaluation of the Effects of MLT on Anxiety-like Behaviors: OFT, Stereotypic Behaviors and Epmt
3.2.2. Evaluation of the Effects of MLT on Sociability: Three-Chambers Sociability Test
3.2.3. Evaluation of the Effects of MLT on Obsessive–Compulsive Disorder (OCD) Behavior: Marble Burying Test
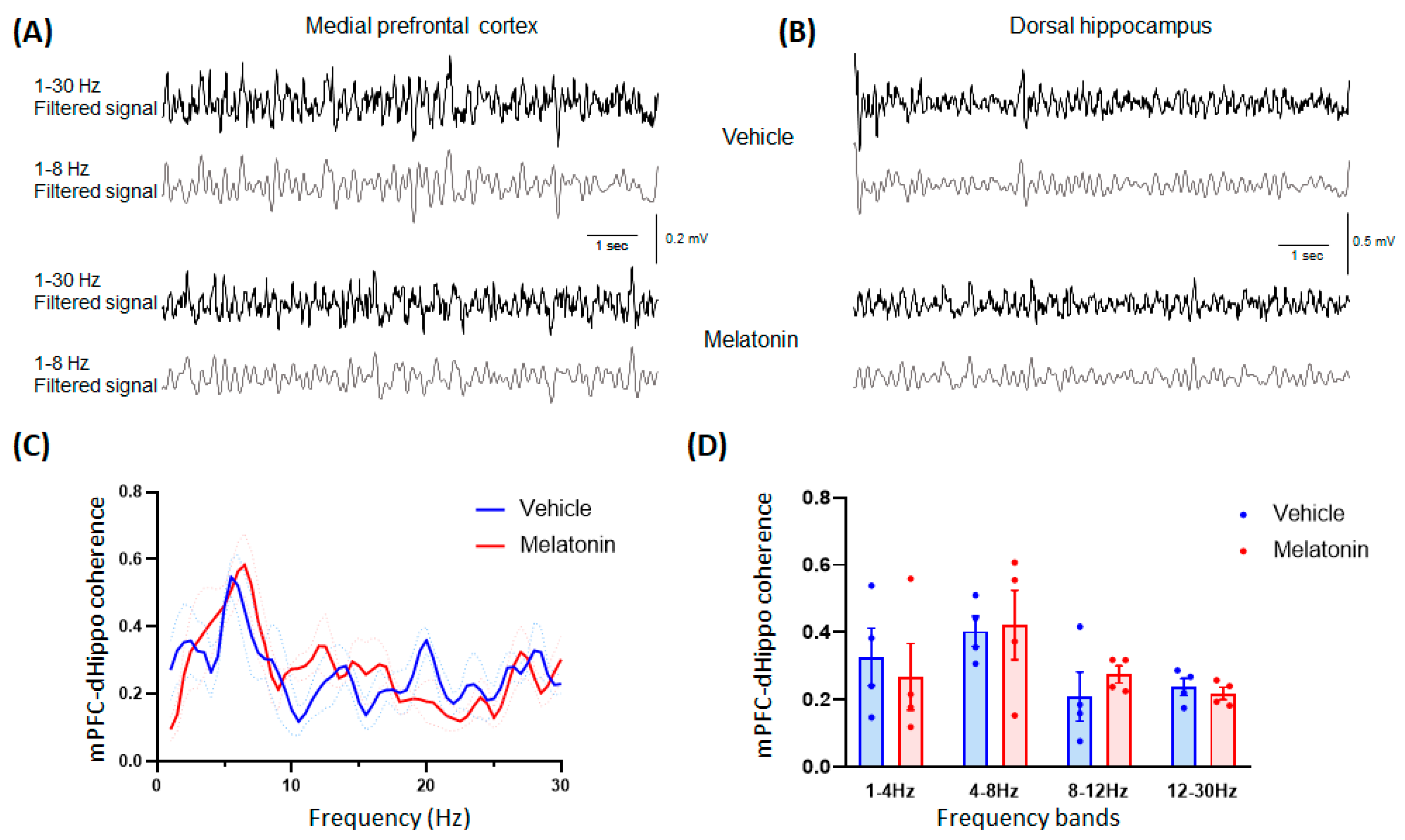
3.2.4. In Vivo Electrophysiology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Mental Health and COVID-19: Early Evidence of the Pandemic’s Impact: Scientific Brief, 2 March 2022 (No. WHO/2019-nCoV/Sci_Brief/Mental_health/2022.1); World Health Organization: Geneva, Switzerland, 2022. [Google Scholar]
- Bandelow, B.; Michaelis, S. Epidemiology of Anxiety Disorders in the 21st Century. Dialogues Clin. Neurosci. 2015, 17, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Xiong, P.; Liu, M.; Liu, B.; Hall, B.J. Trends in the Incidence and DALYs of Anxiety Disorders at the Global, Regional, and National Levels: Estimates from the Global Burden of Disease Study 2019. J. Affect. Disord. 2022, 297, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Koskinen, M.K.; Hovatta, I. Genetic Insights into the Neurobiology of Anxiety. Trends Neurosci. 2023, 46, 318–331. [Google Scholar] [CrossRef]
- The Psychobiology and Pathophysiology of Anxiety and Fear. In Anxiety and the Anxiety Disorders; Routledge: Londomn, UK, 2019; pp. 333–354. [CrossRef]
- Moon, E.; Kim, K.; Partonen, T.; Linnaranta, O. Role of Melatonin in the Management of Sleep and Circadian Disorders in the Context of Psychiatric Illness. Curr. Psychiatry Rep. 2022, 24, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Marseglia, L.; D’Angelo, G.; Manti, S.; Aversa, S.; Arrigo, T.; Reiter, R.J.; Gitto, E. Analgesic, Anxiolytic and Anaesthetic Effects of Melatonin: New Potential Uses in Pediatrics. Int. J. Mol. Sci. 2015, 16, 1209–1220. [Google Scholar] [CrossRef] [Green Version]
- Papp, M.; Litwa, E.; Gruca, P.; Mocaë, E. Anxiolytic-like Activity of Agomelatine and Melatonin in Three Animal Models of Anxiety. Behav. Pharmacol. 2006, 17, 9–18. [Google Scholar]
- Golus, P.; King, M.G. The Effects of Melatonin on Open Field Behavior. Pharmacol. Biochem. Behav. 1981, 15, 883–885. [Google Scholar] [CrossRef]
- Ochoa-Sanchez, R.; Rainer, Q.; Comai, S.; Spadoni, G.; Bedini, A.; Rivara, S.; Fraschini, F.; Mor, M.; Tarzia, G.; Gobbi, G. Anxiolytic Effects of the Melatonin MT2 Receptor Partial Agonist UCM765: Comparison with Melatonin and Diazepam. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2012, 39, 318–325. [Google Scholar] [CrossRef]
- Golombek, D.A.; Martini, M.; Cardinali, D.P. Melatonin as an Anxiolytic in Rats: Time Dependence and Interaction with the Central GABAergic System. Eur. J. Pharmacol. 1993, 237, 231–236. [Google Scholar] [CrossRef]
- Repova, K.; Baka, T.; Krajcirovicova, K.; Stanko, P.; Aziriova, S.; Reiter, R.J.; Simko, F. Melatonin as a Potential Approach to Anxiety Treatment. Int. J. Mol. Sci. 2022, 23, 16187. [Google Scholar] [CrossRef]
- Crupi, R.; Mazzon, E.; Marino, A.; La Spada, G.; Bramanti, P.; Cuzzocrea, S.; Spina, E. Melatonin Treatment Mimics the Antidepressant Action in Chronic Corticosterone-Treated Mice. J. Pineal Res. 2010, 49, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Samarkandi, A.; Naguib, M.; Riad, W.; Thalaj, A.; Alotibi, W.; Aldammas, F.; Albassam, A. Melatonin vs. Midazolam Premedication in Children: A Double-Blind, Placebo-Controlled Study. Eur. J. Anaesthesiol. 2005, 22, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Madsen, B.K.; Zetner, D.; Møller, A.M.; Rosenberg, J. Melatonin for Preoperative and Postoperative Anxiety in Adults. Cochrane Database Syst. Rev. 2020, 12, 12. [Google Scholar] [CrossRef]
- Karasek, M. Does Melatonin Play a Role in Aging Processes? J. Physiol. Pharmacol. 2007, 58 (Suppl. S6), 105–113. [Google Scholar] [PubMed]
- Jockers, R.; Delagrange, P.; Dubocovich, M.L.; Markus, R.P.; Renault, N.; Tosini, G.; Cecon, E.; Zlotos, D.P. Update on Melatonin Receptors: IUPHAR Review 20. Br. J. Pharmacol. 2016, 173, 2702–2725. [Google Scholar] [CrossRef] [Green Version]
- Gobbi, G.; Comai, S. Sleep Well. Untangling the Role of Melatonin MT1 and MT2 Receptors in Sleep. J. Pineal Res. 2019, 66, e12544. [Google Scholar] [CrossRef]
- López-Canul, M.; Hyun Min, S.; Posa, L.; De Gregorio, D.; Bedini, A.; Spadoni, G.; Gobbi, G.; Comai, S. Melatonin MT1 and MT2 Receptors Exhibit Distinct Effects in the Modulation of Body Temperature across the Light/Dark Cycle. Int. J. Mol. Sci. 2019, 20, 2452. [Google Scholar] [CrossRef] [Green Version]
- Slominski, R.M.; Reiter, R.J.; Schlabritz-Loutsevitch, N.; Ostrom, R.S.; Slominski, A.T. Melatonin Membrane Receptors in Peripheral Tissues: Distribution and Functions. Mol. Cell. Endocrinol. 2012, 351, 152–166. [Google Scholar] [CrossRef] [Green Version]
- Comai, S.; De Gregorio, D.; Posa, L.; Ochoa-Sanchez, R.; Bedini, A.; Gobbi, G. Dysfunction of Serotonergic Activity and Emotional Responses across the Light-Dark Cycle in Mice Lacking Melatonin MT2 Receptors. J. Pineal Res. 2020, 69, e12653. [Google Scholar] [CrossRef]
- Thomson, D.M.; Mitchell, E.J.; Openshaw, R.L.; Pratt, J.A.; Morris, B.J. Mice Lacking Melatonin MT2 Receptors Exhibit Attentional Deficits, Anxiety and Enhanced Social Interaction. J. Psychopharmacol. 2021, 35, 1265–1276. [Google Scholar] [CrossRef]
- Noseda, A.C.D.; Targa, A.D.S.; Rodrigues, L.S.; Aurich, M.F.; Lima, M.M.S. REM Sleep Deprivation Promotes a Dopaminergic Influence in the Striatal MT2 Anxiolytic-like Effects. Sleep Sci. 2016, 9, 47–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Clough, S.J.; Dubocovich, M.L. Role of the MT1 and MT2 Melatonin Receptors in Mediating Depressive- and Anxiety-like Behaviors in C3H/HeN Mice. Genes Brain Behav. 2017, 16, 546–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacoste, B.; Angeloni, D.; Dominguez-Lopez, S.; Calderoni, S.; Mauro, A.; Fraschini, F.; Descarries, L.; Gobbi, G. Anatomical and Cellular Localization of Melatonin MT1 and MT2 Receptors in the Adult Rat Brain. J. Pineal Res. 2015, 58, 397–417. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.Y.; Leong, M.K.; Liang, H.; Paxinos, G. Melatonin Receptors: Distribution in Mammalian Brain and Their Respective Putative Functions. Brain Struct. Funct. 2017, 222, 2921–2939. [Google Scholar] [CrossRef]
- Sugden, D. Psychopharmacological Effects of Melatonin in Mouse and Rat. J. Pharmacol. Exp. Ther. 1983, 227, 587–591. [Google Scholar]
- Tassan Mazzocco, M.; Guarnieri, F.C.; Monzani, E.; Benfenati, F.; Valtorta, F.; Comai, S. Dysfunction of the Serotonergic System in the Brain of Synapsin Triple Knockout Mice Is Associated with Behavioral Abnormalities Resembling Synapsin-Related Human Pathologies. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 105, 110135. [Google Scholar] [CrossRef]
- Onaolapo, O.J.; Onaolapo, A.Y.; Akanmu, M.A.; Olayiwola, G.; Onaolapo, O.J.; Onaolapo, A.Y.; Akanmu, M.A.; Olayiwola, G. Changes in Spontaneous Working-Memory, Memory-Recall and Approach-Avoidance Following “Low Dose” Monosodium Glutamate in Mice. AIMS Neurosci. 2016, 3, 317–337. [Google Scholar] [CrossRef]
- Comai, S.; Ochoa-Sanchez, R.; Dominguez-Lopez, S.; Bambico, F.R.; Gobbi, G. Melancholic-Like Behaviors and Circadian Neurobiological Abnormalities in Melatonin MT1 Receptor Knockout Mice. Int. J. Neuropsychopharmacol. 2015, 18, pyu075. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Silverman, J.L.; Crawley, J.N. Automated Three-Chambered Social Approach Task for Mice. Curr. Protoc. Neurosci. 2011, 56, 8–26. [Google Scholar] [CrossRef] [Green Version]
- Deacon, R.M.J. Digging and Marble Burying in Mice: Simple Methods for in Vivo Identification of Biological Impacts. Nat. Protoc. 2006, 1, 122–124. [Google Scholar] [CrossRef]
- Njung’e, K.; Handley, S.L. Evaluation of Marble-Burying Behavior as a Model of Anxiety. Pharmacol. Biochem. Behav. 1991, 38, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Cambiaghi, M.; Grosso, A.; Likhtik, E.; Mazziotti, X.R.; Concina, G.; Renna, A.; Sacco, T.; Gordon, J.A.; Benedetto Sacchetti, X.; Levi-Montalcini, R. Higher-Order Sensory Cortex Drives Basolateral Amygdala Activity during the Recall of Remote, but Not Recently Learned Fearful Memories. J. Neurosci. 2016, 36, 1647–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Concina, G.; Cambiaghi, X.M.; Renna, A.; Sacchetti, X.B. Behavioral/Cognitive Coherent Activity between the Prelimbic and Auditory Cortex in the Slow-Gamma Band Underlies Fear Discrimination. J. Neurosci. 2018, 38, 8313–8328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharman, E.H.; Bondy, S.C.; Sharman, K.G.; Lahiri, D.; Cotman, C.W.; Perreau, V.M. Effects of Melatonin and Age on Gene Expression in Mouse CNS Using Microarray Analysis. Neurochem. Int. 2007, 50, 336–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perreau, V.M.; Bondy, S.C.; Cotman, C.W.; Sharman, K.G.; Sharman, E.H. Melatonin Treatment in Old Mice Enables a More Youthful Response to LPS in the Brain. J. Neuroimmunol. 2007, 182, 22–31. [Google Scholar] [CrossRef] [Green Version]
- Roseboom, P.H.; Namboodiri, M.A.A.; Zimonjic, D.B.; Popescu, N.C.; Rodriguez, I.R.; Gastel, J.A.; Klein, D.C. Natural Melatonin ‘knockdown’ in C57BL/6J Mice: Rare Mechanism Truncates Serotonin N-Acetyltransferase. Mol. Brain Res. 1998, 63, 189–197. [Google Scholar] [CrossRef]
- Slominski, A.; Pisarchik, A.; Semak, I.; Sweatman, T.; Wortsman, J. Characterization of the Serotoninergic System in the C57BL/6 Mouse Skin. Eur. J. Biochem. 2003, 270, 3335–3344. [Google Scholar] [CrossRef] [Green Version]
- Slominski, A.; Wortsman, J.; Tobin, D.J. The Cutaneous Serotoninergic/Melatoninergic System: Securing a Place under the Sun. FASEB J. 2005, 19, 176–194. [Google Scholar] [CrossRef]
- Slominski, A.; Pisarchik, A.; Semak, I.; Sweatman, T.; Szczesniewski, A.; Wortsman, J. Serotoninergic System in Hamster Skin. J. Investig. Dermatol. 2002, 119, 934–942. [Google Scholar] [CrossRef] [Green Version]
- Semak, I.; Korik, E.; Naumova, M.; Wortsman, J.; Slominski, A. Serotonin Metabolism in Rat Skin: Characterization by Liquid Chromatography-Mass Spectrometry. Arch. Biochem. Biophys. 2004, 421, 61–66. [Google Scholar] [CrossRef]
- Slominski, A.; Pisarchik, A.; Semak, I.; Sweatman, T.; Wortsman, J.; Szczesniewski, A.; Slugocki, G.; McNulty, J.; Kauser, S.; Tobin, D.J.; et al. Serotoninergic and Melatoninergic Systems Are Fully Expressed in Human Skin. FASEB J. 2002, 16, 896–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slominski, A.T.; Kim, T.K.; Kleszczyński, K.; Semak, I.; Janjetovic, Z.; Sweatman, T.; Skobowiat, C.; Steketee, J.D.; Lin, Z.; Postlethwaite, A.; et al. Characterization of Serotonin and N-Acetylserotonin Systems in the Human Epidermis and Skin Cells. J. Pineal Res. 2020, 68, e12626. [Google Scholar] [CrossRef]
- Ochoa-Sanchez, R.; Comai, S.; Spadoni, G.; Bedini, A.; Tarzia, G.; Gobbi, G. Melatonin, Selective and Non-Selective MT1/MT2 Receptors Agonists: Differential Effects on the 24-h Vigilance States. Neurosci. Lett. 2014, 561, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Clough, S.J.; Adamah-Biassi, E.B.; Sveinsson, M.H.; Hutchinson, A.J.; Miura, I.; Furuse, T.; Wakana, S.; Matsumoto, Y.K.; Okanoya, K.; et al. Impact of Endogenous Melatonin on Rhythmic Behaviors, Reproduction, and Survival Revealed in Melatonin-Proficient C57BL/6J Congenic Mice. J. Pineal Res. 2021, 71, e12748. [Google Scholar] [CrossRef] [PubMed]
- Paulose, J.K.; Wang, C.; O’Hara, B.F.; Cassone, V.M. The Effects of Aging on Sleep Parameters in a Healthy, Melatonin-Competent Mouse Model. Nat. Sci. Sleep 2019, 11, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Witt-Enderby, P.A.; Bennett, J.; Jarzynka, M.J.; Firestine, S.; Melan, M.A. Melatonin Receptors and Their Regulation: Biochemical and Structural Mechanisms. Life Sci. 2003, 72, 2183–2198. [Google Scholar] [CrossRef]
- Gobbi, G.; Comai, S. Differential Function of Melatonin MT1 and MT2 Receptors in REM and NREM Sleep. Front. Endocrinol. 2019, 10, 87. [Google Scholar] [CrossRef] [Green Version]
- Pandi-Perumal, S.R.; Trakht, I.; Srinivasan, V.; Spence, D.W.; Maestroni, G.J.M.; Zisapel, N.; Cardinali, D.P. Physiological Effects of Melatonin: Role of Melatonin Receptors and Signal Transduction Pathways. Prog. Neurobiol. 2008, 85, 335–353. [Google Scholar] [CrossRef]
- Ochoa-Sanchez, R.; Comai, S.; Lacoste, B.; Bambico, F.R.; Dominguez-Lopez, S.; Spadoni, G.; Rivara, S.; Bedini, A.; Angeloni, D.; Fraschini, F.; et al. Promotion of Non-Rapid Eye Movement Sleep and Activation of Reticular Thalamic Neurons by a Novel MT2 Melatonin Receptor Ligand. J. Neurosci. 2011, 31, 18439–18452. [Google Scholar] [CrossRef] [Green Version]
- Harris, A.S.; Burgess, H.J.; Dawson, D. The Effects of Day-Time Exogenous Melatonin Administration on Cardiac Autonomic Activity. J. Pineal Res. 2001, 31, 199–205. [Google Scholar] [CrossRef]
- Tzischinsky, O.; Lavie, P. Melatonin Possesses Time-Dependent Hypnotic Effects. Sleep 1994, 17, 638–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchison, L.C.; O’brien, C.E. Changes in Pharmacokinetics and Pharmacodynamics in the Elderly Patient. J. Pharm. Pract. 2007, 20, 4–12. [Google Scholar] [CrossRef]
- Leary, M.R.; Kowalski, R.M. Social Anxiety; Guilford Press: New York, NY, USA, 1997. [Google Scholar]
- Maddox, B.B.; White, S.W. Comorbid Social Anxiety Disorder in Adults with Autism Spectrum Disorder. J. Autism Dev. Disord. 2015, 45, 3949–3960. [Google Scholar] [CrossRef] [PubMed]
- van Steensel, F.J.A.; Bögels, S.M.; Perrin, S. Anxiety Disorders in Children and Adolescents with Autistic Spectrum Disorders: A Meta-Analysis. Clin. Child Fam. Psychol. Rev. 2011, 14, 302–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossignol, D.; Frye, R. Melatonin in Autism Spectrum Disorders. Curr. Clin. Pharmacol. 2014, 9, 326–334. [Google Scholar] [CrossRef]
- Liu, X.; Cui, Y.; Zhang, Y.; Xiang, G.; Yu, M.; Wang, X.; Qiu, B.; Li, X.; Liu, W.; Zhang, D. Rescue of Social Deficits by Early-Life Melatonin Supplementation through Modulation of Gut Microbiota in a Murine Model of Autism. Biomed. Pharmacother. 2022, 156, 113949. [Google Scholar] [CrossRef]
- Kumar, H.; Sharma, B.M.; Sharma, B. Benefits of Agomelatine in Behavioral, Neurochemical and Blood Brain Barrier Alterations in Prenatal Valproic Acid Induced Autism Spectrum Disorder. Neurochem. Int. 2015, 91, 34–45. [Google Scholar] [CrossRef]
- Adhikari, A.; Topiwala, M.A.; Gordon, J.A. Synchronized Activity between the Ventral Hippocampus and the Medial Prefrontal Cortex during Anxiety. Neuron 2010, 65, 257–269. [Google Scholar] [CrossRef] [Green Version]
- Arushanian, E.B.; Beier, E.V. The Participation of the Dorsal Hippocampus in the Antianxiety Action of Melatonin and Diazepam. Eksperimental’naia Klin. Farmakol. 1998, 61, 13–16. [Google Scholar]
- Lee, J.L.C. Memory Reconsolidation Mediates the Strengthening of Memories by Additional Learning. Nat. Neurosci. 2008, 11, 1264–1266. [Google Scholar] [CrossRef] [Green Version]
- Kindt, M.; Elsey, J.W.B. A Paradigm Shift in the Treatment of Emotional Memory Disorders: Lessons from Basic Science. Brain Res. Bull. 2023, 192, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, H.; Zhang, Y.; Archbold, G.; Ishikawa, R.; Nader, K.; Kida, S. Enhancement of Fear Memory by Retrieval through Reconsolidation. eLife 2014, 3, e02736. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Kapeller-Libermann, D.; Travaglia, A.; Inda, M.C.; Alberini, C.M. Direct Dorsal Hippocampal-Prelimbic Cortex Connections Strengthen Fear Memories. Nat. Neurosci. 2017, 20, 52–61. [Google Scholar] [CrossRef] [Green Version]
- Colgin, L.L. Oscillations and Hippocampal-Prefrontal Synchrony. Curr. Opin. Neurobiol. 2011, 21, 467–474. [Google Scholar] [CrossRef] [Green Version]
- Sigurdsson, T.; Stark, K.L.; Karayiorgou, M.; Gogos, J.A.; Gordon, J.A. Impaired Hippocampal-Prefrontal Synchrony in a Genetic Mouse Model of Schizophrenia. Nature 2010, 464, 763–767. [Google Scholar] [CrossRef] [Green Version]
- Kuga, N.; Abe, R.; Takano, K.; Ikegaya, Y.; Sasaki, T. Prefrontal-amygdalar oscillations related to social behavior in mice. eLife 2022, 11, e78428. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, N.S.; Cavanagh, J.F.; Frank, M.J.; Laubach, M. Common medial frontal mechanisms of adaptive control in humans and rodents. Nat. Neurosci. 2013, 16, 1888–1895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britton, J.C.; Grillon, C.; Lissek, S.; Norcross, M.A.; Szuhany, K.L.; Chen, G.; Ernst, M.; Nelson, E.E.; Leibenluft, E.; Shechner, T.; et al. Response to Learned Threat: An FMRI Study in Adolescent and Adult Anxiety. Am. J. Psychiatry 2013, 170, 1195–1204. [Google Scholar] [CrossRef]
- Jarcho, J.M.; Romer, A.L.; Shechner, T.; Galvan, A.; Guyer, A.E.; Leibenluft, E.; Pine, D.S.; Nelson, E.E. Forgetting the Best When Predicting the Worst: Preliminary Observations on Neural Circuit Function in Adolescent Social Anxiety. Dev. Cogn. Neurosci. 2015, 13, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Gunning-Dixon, F.M.; Gur, R.C.; Perkins, A.C.; Schroeder, L.; Turner, T.; Turetsky, B.I.; Chan, R.M.; Loughead, J.W.; Alsop, D.C.; Maldjian, J.; et al. Age-Related Differences in Brain Activation during Emotional Face Processing. Neurobiol. Aging 2003, 24, 285–295. [Google Scholar] [CrossRef]
- Ganella, D.E.; Drummond, K.D.; Ganella, E.P.; Whittle, S.; Kim, J.H. Extinction of Conditioned Fear in Adolescents and Adults: A Human FMRI Study. Front. Hum. Neurosci. 2018, 11, 647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Somers, V.K.; Xu, H.; Lopez-Jimenez, F.; Covassin, N. Trends in Use of Melatonin Supplements Among US Adults, 1999–2018. JAMA 2022, 327, 483–485. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Figure | Panel | Test | Group-Size | Statistic | p Value | Pair-Wise Comparison | Statistic 2 | |||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | A | Open field test | Student’s t-test | Vehicle = 11 mice | locomotor activity t = 0.9675; df = 21 | p = 0.3443 | N/A | N/A | ||
| time spent in centre t = 0.03147; df = 21 | p = 0.9752 | |||||||||
| MLT 20 mg/kg = 10 mice | entries in centre t = 0.193; df = 21 | p = 0.8488 | ||||||||
| B | Stereotypic behaviours | Student’s t-test | Vehicle = 11 mice | grooming events t = 2.18; df = 20 | p = 0.0414 | N/A | N/A | |||
| time of grooming t = 1.167; df = 20 | p = 0.257 | |||||||||
| MLT 20 mg/kg = 11 mice | rearing events t = 0.1577; df = 20 | p = 0.8763 | ||||||||
| time of rearing t = 0.1773; df = 20 | p = 0.8610 | |||||||||
| C | Elevated Plus Maze | Student’s t-test | Vehicle = 13 mice | entries in open arms t = 0.3292; df = 23 | p = 0.745 | N/A | N/A | |||
| MLT 20 mg/kg = 12 mice | time in open arms t = 0.4702; df = 23 | p = 0.6427 | ||||||||
| time in close arms t = 1.469; df = 23 | p = 0.1553 | |||||||||
| D | 3-chamber sociability test | Two-way ANOVA | Vehicle = 12 mice | Sociability: | Test Details | t | p Value | |||
| interaction F (1,17) = 8.171 | p = 0.0109 | Bonferroni post hoc comparison | empty cage vehicle vs. empty cage MLT 20 mg/kg | 0.0695 | p > 0.9999 | |||||
| treatment F(1,17) = 2.886 | p = 0.1076 | mouse-1 vehicle vs. mouse-1 MLT 20 mg/kg | 2.953 | p = 0.0113 | ||||||
| sociability F (1,17) = 73.49 | p < 0.0001 | vehicle empty cage vs. vehicle mouse-1 | 4.707 | p = 0.0004 | ||||||
| MLT 20 mg/kg empty cage vs. MLT 20 m/kg mouse-1 | 7.192 | p < 0.0001 | ||||||||
| MLT 20 mg/kg = 7 mice | Social novelty: | N/A | N/A | |||||||
| interaction F (1,17) = 0.1712 | p = 0.6842 | |||||||||
| treatment F (1,17) = 4.570 | p = 0.0474 | |||||||||
| social novelty F (1,17) = 22.93 | p < 0.0001 | |||||||||
| E | Marble burying test | Student’s t-test | Vehicle = 15 mice | t = 1.454; df = 23 | p = 0.1594 | N/A | N/A | |||
| MLT 20 mg/kg = 10 mice | ||||||||||
| 2 | C | mPFC-dHippo coherence | Two-way ANOVA | Vehicle = 4 mice | interaction F (58,354) = 0.8933 | p = 0.6937 | N/A | N/A | ||
| frequency F (58,354) = 3.410 | p < 0.0001 | |||||||||
| MLT 20 mg/kg = 4 mice | treatment F (58,354) = 0.1881 | p = 0.6648 | ||||||||
| D | mPFC-dHippo coherence | Student’s t-test | Vehicle = 4 mice | 1–4 Hz t = 0.4544; df = 6 | p = 0.6655 | N/A | N/A | |||
| 4–8 Hz t = 0.1669; df = 6 | p = 0.8729 | |||||||||
| MLT 20 mg/kg = 4 mice | 8–12 Hz t = 0.8479; df = 6 | p = 0.4290 | ||||||||
| 12–30 Hz t = 0.6128; df = 6 | p = 0.5625 | |||||||||
| Figure | Panel | Test | Group-Size | Statistic | p Value | Pair-Wise Comparison | Statistic 2 | |||
|---|---|---|---|---|---|---|---|---|---|---|
| 3 | A | Open field test | Student’s t-test | Vehicle = 11 mice | locomotor activity t = 3.011; df = 19 | p = 0.0072 | N/A | N/A | ||
| time spent in centre t = 3.096; df = 19 | p = 0.006 | |||||||||
| MLT 20 mg/kg = 10 mice | entries in centre t = 3.319; df = 19 | p = 0.0036 | ||||||||
| B | Stereotypic behaviours | Student’s t-test | Vehicle = 10 mice | grooming events t = 3.138; df = 19 | p = 0.0054 | N/A | N/A | |||
| time of grooming t = 2.336; df = 19 | p = 0.0306 | |||||||||
| MLT 20 mg/kg = 11 mice | rearing events t = 1.102; df = 19 | p = 0.2843 | ||||||||
| time of rearing t = 1.377; df = 19 | p = 0.1844 | |||||||||
| C | Elevated Plus Maze | Student’s t-test | Vehicle = 13 mice; | entries in open arms t = 1.32; df = 21 | p = 0.2011 | N/A | N/A | |||
| MLT 20 mg/kg = 10 mice | time in open arms t = 3.260; df = 21 | p = 0.0037 | ||||||||
| time in close arms t = 0.177; df = 21 | p = 0.8612 | |||||||||
| D | 3-chamber sociability test | Two-way ANOVA | Vehicle = 11 mice | Sociability: | Test Details | t | p Value | |||
| interaction F(1,19) = 9.176 | p = 0.0069 | Bonferroni post hoc comparison | vehicle empty cage vs. vehicle mouse-1 | 7.172 | p < 0.0001 | |||||
| treatment F(1,19) = 26.29 | p < 0.0001 | MLT 20 mg/kg empty cage vs. MLT 20 mg/kg mouse-1 | 3.132 | p = 0.0110 | ||||||
| sociability F(1,19) = 54.06 | p < 0.0001 | empty cage vehicle vs. empty cage MLT 20 mg/kg | 2.53 | p = 0.0314 | ||||||
| mouse-1 vehicle vs mouse-1 MLT 20 mg/kg | 5.943 | p < 0.0001 | ||||||||
| MLT 20 mg/kg = 11 mice | Social novelty: | Test Details | t | p Value | ||||||
| interaction F(1,19) = 9.559 | p = 0.006 | Bonferroni post hoc comparison | vehicle mouse-1 vs. vehicle mouse-2 | 5.729 | p < 0.0001 | |||||
| treatment F(1,19) = 26.46 | p < 0.0001 | MLT 20 mg/kg mouse-1 vs. MLT 20 mg/kg mouse-2 | 1.529 | p = 0.2857 | ||||||
| social novelty F(1,19) = 27.06 | p < 0.0001 | mouse-1 vehicle vs. mouse-1 MLT 20 mg/kg | 3.11 | p = 0.0071 | ||||||
| mouse-2 vehicle vs. mouse-2 MLT 20 mg/kg | 6.246 | p < 0.0001 | ||||||||
| E | Marble burying test | Student’s t-test | Vehicle = 12 mice | t = 0.6125; df = 22 | p = 0.5465 | N/A | N/A | |||
| MLT 20 mg/kg = 12 mice | ||||||||||
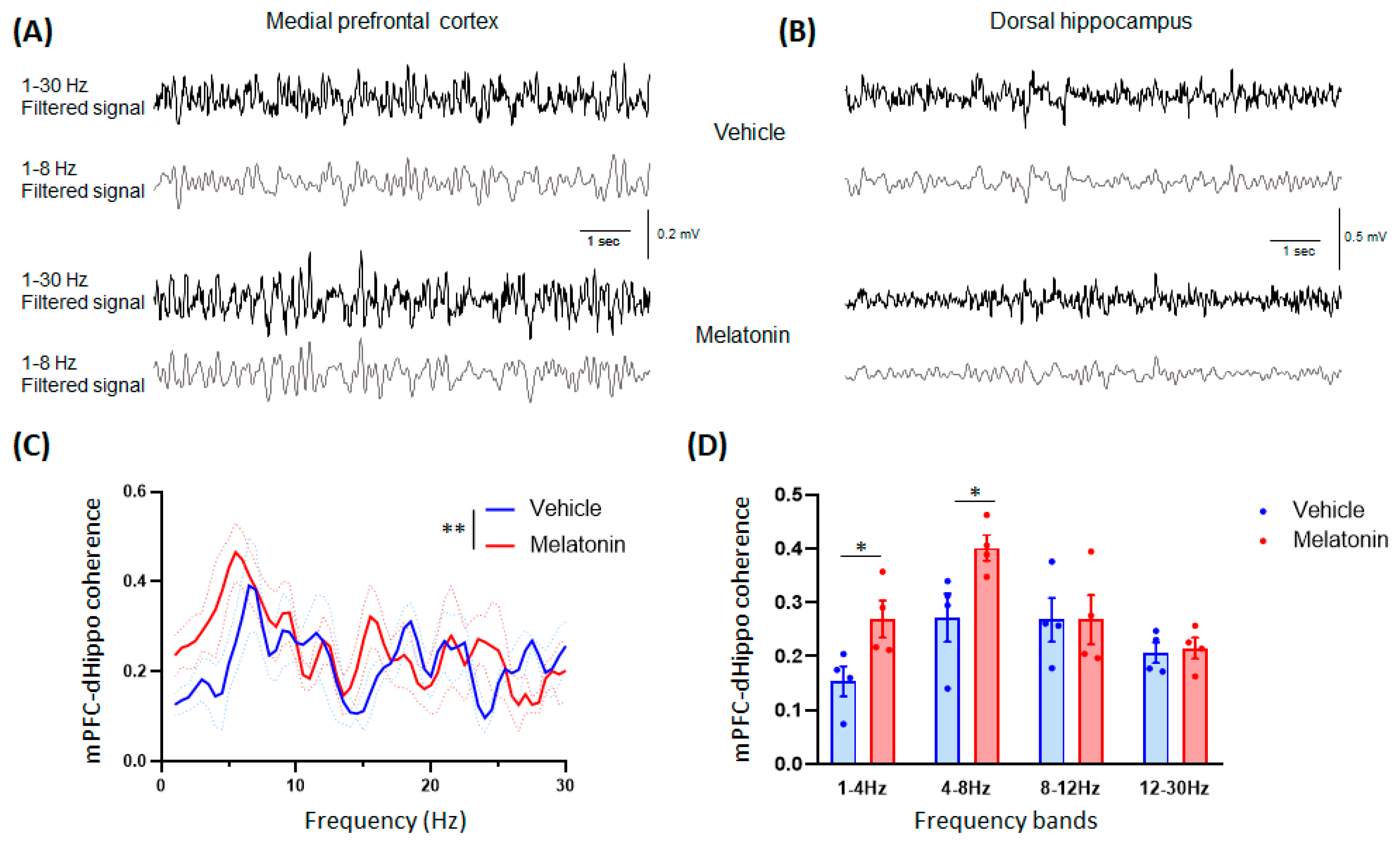
| 4 | C | mPFC-dHippo coherence | Two-way ANOVA | Vehicle = 4 mice | interaction F (1,354) | p = 0.3028 | N/A | N/A | ||
| frequency F (1,354) | p = 0.0008 | |||||||||
| MLT 20 mg/kg = 4 mice | treatment F (1,354) | p = 0.0025 | ||||||||
| D | mPFC-dHippo coherence | Student’s t-test | Vehicle = 4 mice | 1–4 Hz t = 2.613; df = 6 | p = 0.04 | N/A | N/A | |||
| 4–8 Hz t = 2.553; df = 6 | p = 0.0433 | |||||||||
| MLT 20 mg/kg = 4 mice | 8–12 Hz t = 0.001256; df = 6 | p = 0.9990 | ||||||||
| 12–30 Hz t = 0.3003; df = 6 | p = 0.7741 | |||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nasini, S.; Tidei, S.; Shkodra, A.; De Gregorio, D.; Cambiaghi, M.; Comai, S. Age-Related Effects of Exogenous Melatonin on Anxiety-like Behavior in C57/B6J Mice. Biomedicines 2023, 11, 1705. https://doi.org/10.3390/biomedicines11061705
Nasini S, Tidei S, Shkodra A, De Gregorio D, Cambiaghi M, Comai S. Age-Related Effects of Exogenous Melatonin on Anxiety-like Behavior in C57/B6J Mice. Biomedicines. 2023; 11(6):1705. https://doi.org/10.3390/biomedicines11061705
Chicago/Turabian StyleNasini, Sofia, Sara Tidei, Atea Shkodra, Danilo De Gregorio, Marco Cambiaghi, and Stefano Comai. 2023. "Age-Related Effects of Exogenous Melatonin on Anxiety-like Behavior in C57/B6J Mice" Biomedicines 11, no. 6: 1705. https://doi.org/10.3390/biomedicines11061705
APA StyleNasini, S., Tidei, S., Shkodra, A., De Gregorio, D., Cambiaghi, M., & Comai, S. (2023). Age-Related Effects of Exogenous Melatonin on Anxiety-like Behavior in C57/B6J Mice. Biomedicines, 11(6), 1705. https://doi.org/10.3390/biomedicines11061705