


Melatonin Improves Vasogenic Edema via Inhibition to Water Channel Aquaporin-4 (AQP4) and Metalloproteinase-9 (MMP-9) Following Permanent Focal Cerebral Ischemia

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Distal Middle Cerebral Artery Occlusion
2.3. Euthanasia and Quantification of Brain Infarct
2.4. Immunofluorescence Staining
2.5. Evans Blue (EB) Leakage Assessment
2.6. Immunoblotting
2.7. Electromicroscopic Examinations
2.8. Neurobehavioral Scoring
2.9. Statistical Analysis
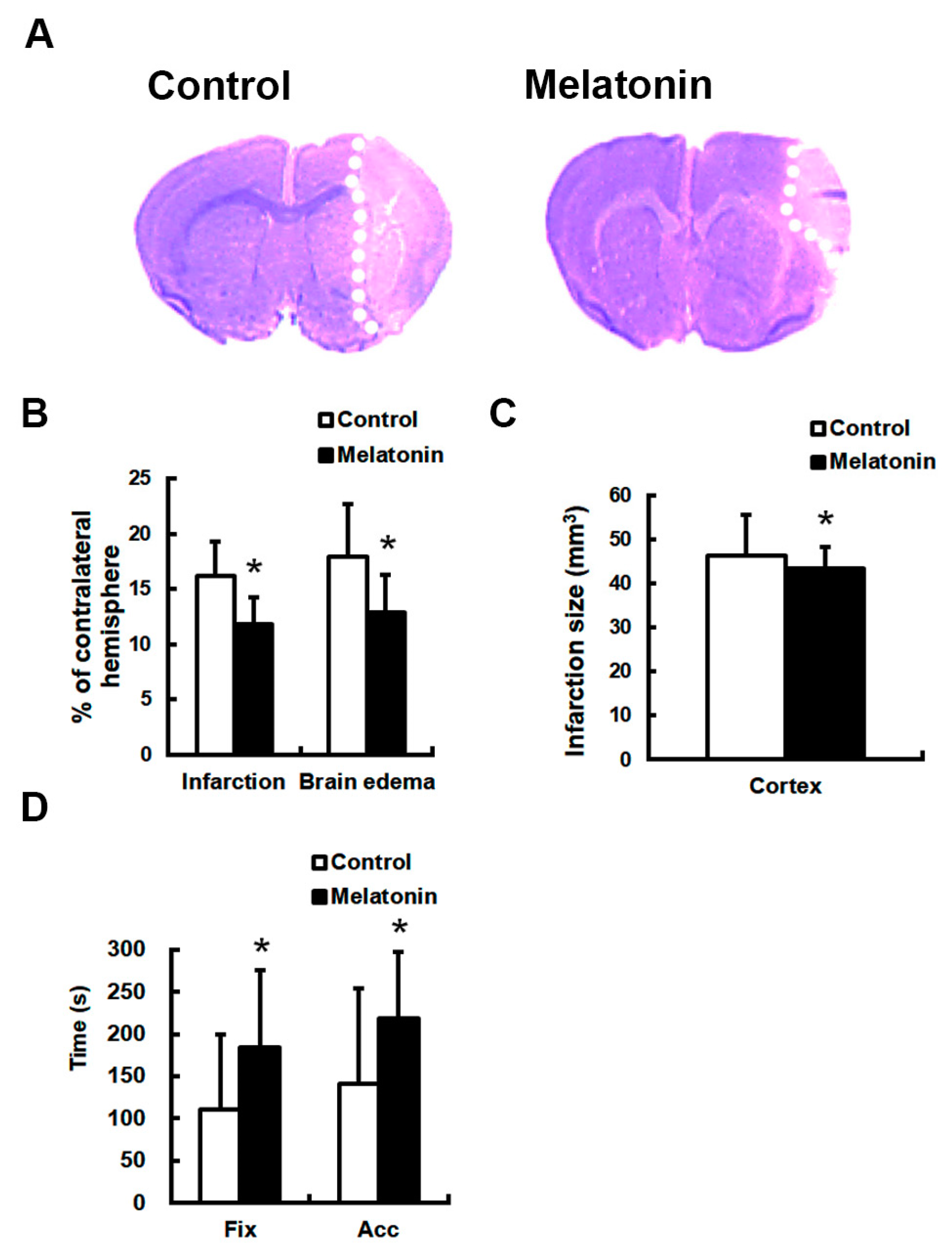
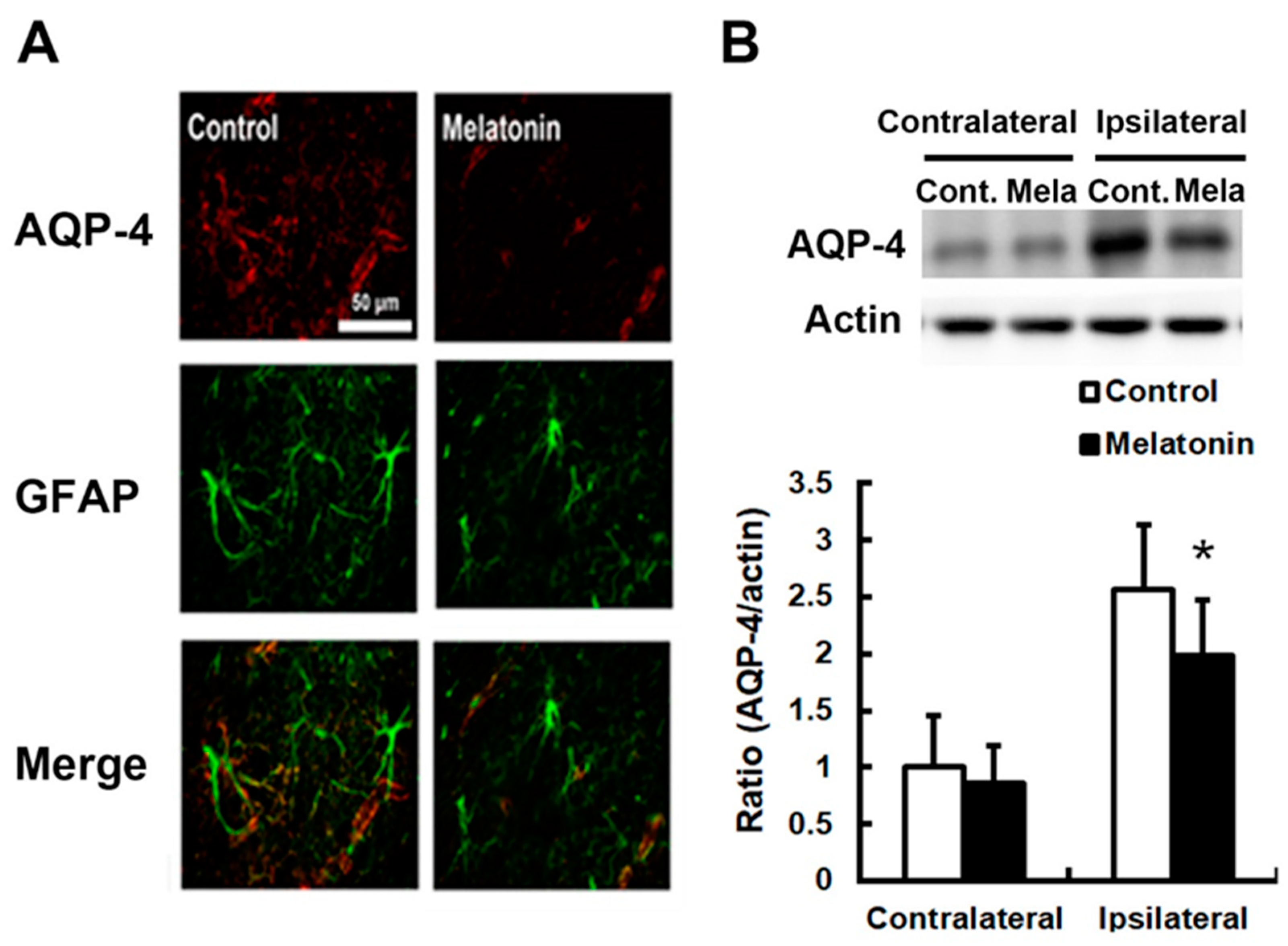
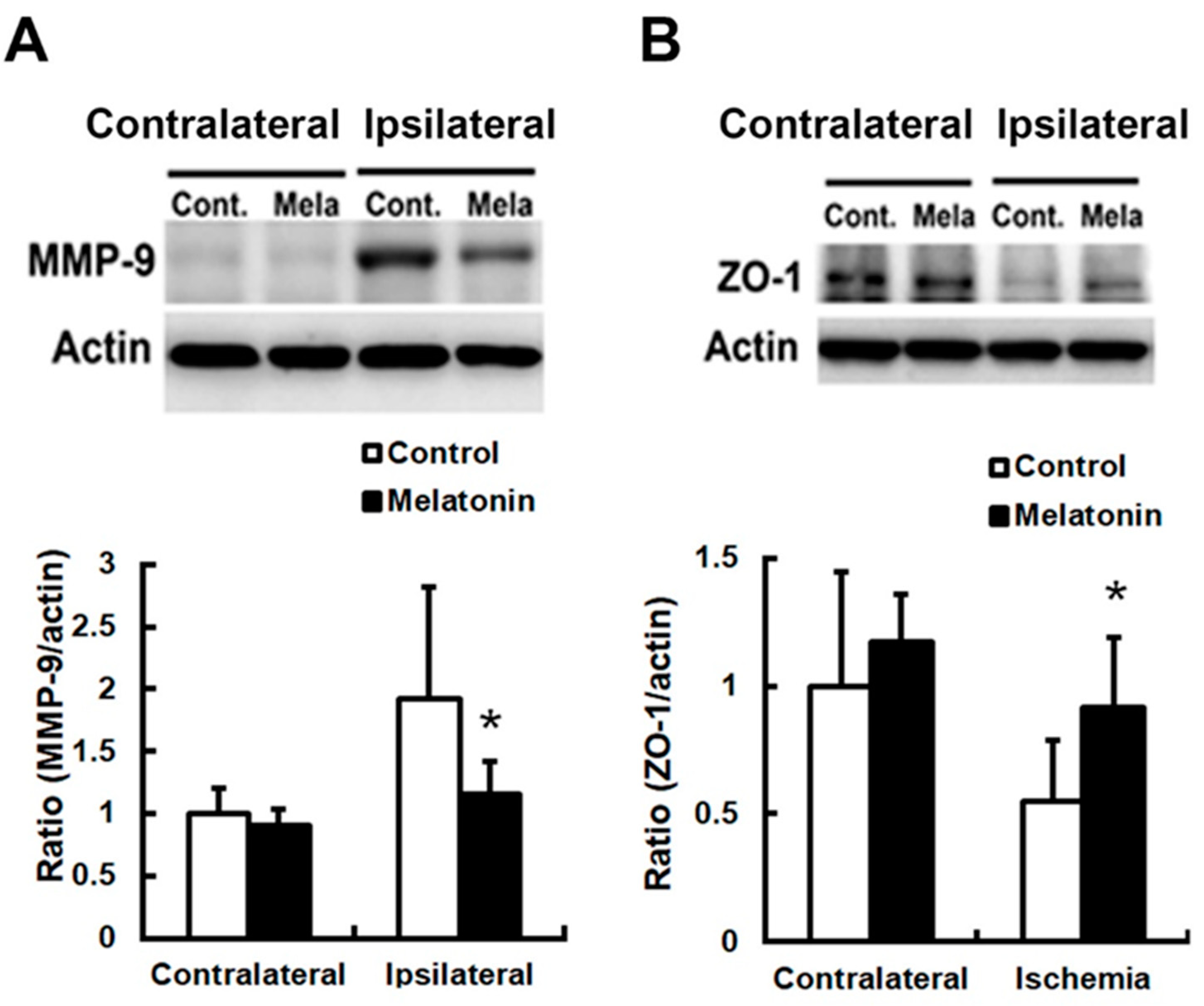
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Choi, D.W. Glutamate neurotoxicity and diseases of the nervous system. Neuron 1988, 1, 623–634. [Google Scholar] [CrossRef]
- Hazell, A.S. Excitotoxic mechanisms in stroke: An update of concepts and treatment strategies. Neurochem. Int. 2007, 50, 941–953. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Dou, J.; Luo, J.; Li, W.; Chan, H.H.; Cui, W.; Zhang, H.; Han, R.; Carlier, P.R.; Zhang, X.; et al. Neuroprotection against excitotoxic and ischemic insults by bis(12)-hupyridone, a novel anti-acetylcholinesterase dimer, possibly via acting on multiple targets. Brain Res. 2011, 1421, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Gasche, Y.; Copin, J.C.; Sugawara, T.; Fujimura, M.; Chan, P.H. Matrix metalloproteinase inhibition prevents oxidative stress-associated blood-brain barrier disruption after transient focal cerebral ischemia. J. Cereb. Blood Flow. Metab. 2001, 21, 1393–1400. [Google Scholar] [CrossRef] [PubMed]
- Hung, Y.C.; Chen, T.Y.; Lee, E.J.; Chen, W.L.; Huang, S.Y.; Lee, W.T.; Lee, M.Y.; Chen, H.Y.; Wu, T.S. Melatonin decreases matrix metalloproteinase-9 activation and expression and attenuates reperfusion-induced hemorrhage following transient focal cerebral ischemia in rats. J. Pineal Res. 2008, 45, 459–467. [Google Scholar] [CrossRef]
- Tai, S.H.; Chen, H.Y.; Lee, E.J.; Chen, T.Y.; Lin, H.W.; Hung, Y.C.; Huang, S.Y.; Chen, Y.H.; Lee, W.T.; Wu, T.S. Melatonin inhibits postischemic matrix metalloproteinase-9 (MMP-9) activation via dual modulation of plasminogen/plasmin system and endogenous MMP inhibitor in mice subjected to transient focal cerebral ischemia. J. Pineal Res. 2010, 49, 332–341. [Google Scholar] [CrossRef]
- Yang, B.; Zador, Z.; Verkman, A.S. Glial cell aquaporin-4 overexpression in transgenic mice accelerates cytotoxic brain swelling. J. Biol. Chem. 2008, 283, 15280–15286. [Google Scholar] [CrossRef]
- Manley, G.T.; Fujimura, M.; Ma, T.; Noshita, N.; Filiz, F.; Bollen, A.W.; Chan, P.; Verkman, A.S. Aquaporin-4 deletion in mice reduces brain edema after acute water intoxication and ischemic stroke. Nat. Med. 2000, 6, 159–163. [Google Scholar] [CrossRef]
- Papadopoulos, M.C.; Manley, G.T.; Krishna, S.; Verkman, A.S. Aquaporin-4 facilitates reabsorption of excess fluid in vasogenic brain edema. Faseb J. 2004, 18, 1291–1293. [Google Scholar] [CrossRef]
- Saadoun, S.; Papadopoulos, M.C.; Watanabe, H.; Yan, D.; Manley, G.T.; Verkman, A.S. Involvement of aquaporin-4 in astroglial cell migration and glial scar formation. J. Cell Sci. 2005, 118 Pt 24, 5691–5698. [Google Scholar] [CrossRef]
- Verkman, A.S.; Binder, D.K.; Bloch, O.; Auguste, K.; Papadopoulos, M.C. Three distinct roles of aquaporin-4 in brain function revealed by knockout mice. Biochim. Biophys. Acta 2006, 1758, 1085–1093. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.Q.; Ikeda, Y.; Ihara, H.; Urano, T.; Fan, W.; Mikawa, S.; Suzuki, Y.; Kondo, K.; Sato, K.; Nagai, N.; et al. Essential role of endogenous tissue plasminogen activator through matrix metalloproteinase 9 induction and expression on heparin-produced cerebral hemorrhage after cerebral ischemia in mice. Blood 2004, 103, 2610–2616. [Google Scholar] [CrossRef] [PubMed]
- Romanic, A.M.; White, R.F.; Arleth, A.J.; Ohlstein, E.H.; Barone, F.C. Matrix metalloproteinase expression increases after cerebral focal ischemia in rats: Inhibition of matrix metalloproteinase-9 reduces infarct size. Stroke 1998, 29, 1020–1030. [Google Scholar] [CrossRef] [PubMed]
- Torii, K.; Uneyama, H.; Nishino, H.; Kondoh, T. Melatonin suppresses cerebral edema caused by middle cerebral artery occlusion/reperfusion in rats assessed by magnetic resonance imaging. J. Pineal Res. 2004, 36, 18–24. [Google Scholar] [CrossRef]
- Chen, T.Y.; Lee, M.Y.; Chen, H.Y.; Kuo, Y.L.; Lin, S.C.; Wu, T.S.; Lee, E.J. Melatonin attenuates the postischemic increase in blood-brain barrier permeability and decreases hemorrhagic transformation of tissue-plasminogen activator therapy following ischemic stroke in mice. J. Pineal Res. 2006, 40, 242–250. [Google Scholar] [CrossRef]
- Chen, H.Y.; Chen, T.Y.; Lee, M.Y.; Chen, S.T.; Hsu, Y.S.; Kuo, Y.L.; Chang, G.L.; Wu, T.S.; Lee, E.J. Melatonin decreases neurovascular oxidative/nitrosative damage and protects against early increases in the blood-brain barrier permeability after transient focal cerebral ischemia in mice. J. Pineal Res. 2006, 41, 175–182. [Google Scholar] [CrossRef]
- Tan, D.X.; Manchester, L.C.; Terron, M.P.; Flores, L.J.; Reiter, R.J. One molecule, many derivatives: A never-ending interaction of melatonin with reactive oxygen and nitrogen species? J. Pineal Res. 2007, 42, 28–42. [Google Scholar] [CrossRef]
- Hardeland, R.; Tan, D.X.; Reiter, R.J. Kynuramines, metabolites of melatonin and other indoles: The resurrection of an almost forgotten class of biogenic amines. J. Pineal Res. 2009, 47, 109–126. [Google Scholar] [CrossRef]
- Esposito, E.; Genovese, T.; Caminiti, R.; Bramanti, P.; Meli, R.; Cuzzocrea, S. Melatonin regulates matrix metalloproteinases after traumatic experimental spinal cord injury. J. Pineal Res. 2008, 45, 149–156. [Google Scholar] [CrossRef]
- Paul, S.; Sharma, A.V.; Mahapatra, P.D.; Bhattacharya, P.; Reiter, R.J.; Swarnakar, S. Role of melatonin in regulating matrix metalloproteinase-9 via tissue inhibitors of metalloproteinase-1 during protection against endometriosis. J. Pineal Res. 2008, 44, 439–449. [Google Scholar] [CrossRef]
- Esposito, E.; Mazzon, E.; Riccardi, L.; Caminiti, R.; Meli, R.; Cuzzocrea, S. Matrix metalloproteinase-9 and metalloproteinase-2 activity and expression is reduced by melatonin during experimental colitis. J. Pineal Res. 2008, 45, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, K.; Swarnakar, S. Induction of matrix metalloproteinase-9 and -3 in nonsteroidal anti-inflammatory drug-induced acute gastric ulcers in mice: Regulation by melatonin. J. Pineal Res. 2009, 47, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Paul, S.; Swarnakar, S. Downregulation of matrix metalloproteinase-9 by melatonin during prevention of alcohol-induced liver injury in mice. Biochimie 2011, 93, 854–866. [Google Scholar] [CrossRef]
- Lee, M.Y.; Kuan, Y.H.; Chen, H.Y.; Chen, T.Y.; Chen, S.T.; Huang, C.C.; Yang, I.P.; Hsu, Y.S.; Wu, T.S.; Lee, E.J. Intravenous administration of melatonin reduces the intracerebral cellular inflammatory response following transient focal cerebral ischemia in rats. J. Pineal Res. 2007, 42, 297–309. [Google Scholar] [CrossRef]
- Chang, C.C.; Tien, C.H.; Lee, E.J.; Juan, W.S.; Chen, Y.H.; Hung, Y.C.; Chen, T.Y.; Chen, H.Y.; Wu, T.S. Melatonin inhibits matrix metalloproteinase-9 (MMP-9) activation in the lipopolysaccharide (LPS)-stimulated RAW 264.7 and BV2 cells and a mouse model of meningitis. J. Pineal Res. 2012, 53, 188–197. [Google Scholar] [CrossRef]
- Tamura, A.; Graham, D.I.; McCulloch, J.; Teasdale, G.M. Focal cerebral ischaemia in the rat: 1. Description of technique and early neuropathological consequences following middle cerebral artery occlusion. J. Cereb. Blood Flow. Metab. 1981, 1, 53–60. [Google Scholar] [CrossRef]
- Hung, Y.C.; Chou, Y.S.; Chang, C.H.; Lin, H.W.; Chen, H.Y.; Chen, T.Y.; Tai, S.H.; Lee, E.J. Early reperfusion improves the recovery of contralateral electrophysiological diaschisis following focal cerebral ischemia in rats. Neurol. Res. 2010, 32, 828–834. [Google Scholar] [CrossRef]
- Lee, E.J.; Lee, M.Y.; Chen, H.Y.; Hsu, Y.S.; Wu, T.S.; Chen, S.T.; Chang, G.L. Melatonin attenuates gray and white matter damage in a mouse model of transient focal cerebral ischemia. J. Pineal Res. 2005, 38, 42–52. [Google Scholar] [CrossRef]
- Lee, E.J.; Chen, H.Y.; Lee, M.Y.; Chen, T.Y.; Hsu, Y.S.; Hu, Y.L.; Chang, G.L.; Wu, T.S. Cinnamophilin reduces oxidative damage and protects against transient focal cerebral ischemia in mice. Free Radic. Biol. Med. 2005, 39, 495–510. [Google Scholar] [CrossRef]
- Swanson, R.A.; Morton, M.T.; Tsao-Wu, G.; Savalos, R.A.; Davidson, C.; Sharp, F.R. A semiautomated method for measuring brain infarct volume. J. Cereb. Blood Flow. Metab. 1990, 10, 290–293. [Google Scholar] [CrossRef]
- Chen, T.Y.; Tai, S.H.; Lee, E.J.; Huang, C.C.; Lee, A.C.; Huang, S.Y.; Wu, T.S. Cinnamophilin offers prolonged neuroprotection against gray and white matter damage and improves functional and electrophysiological outcomes after transient focal cerebral ischemia. Crit. Care Med. 2011, 39, 1130–1137. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.Y.; Lin, M.H.; Lee, W.T.; Huang, S.Y.; Chen, Y.H.; Lee, A.C.; Lin, H.W.; Lee, E.J. Nicotinamide inhibits nuclear factor-kappa B translocation after transient focal cerebral ischemia. Crit. Care Med. 2012, 40, 532–537. [Google Scholar] [CrossRef] [PubMed]
- Belayev, L.; Alonso, O.F.; Busto, R.; Zhao, W.; Ginsberg, M.D. Middle cerebral artery occlusion in the rat by intraluminal suture. Neurological and pathological evaluation of an improved model. Stroke 1996, 27, 1616–1622; discussion 23. [Google Scholar] [CrossRef] [PubMed]
- Clark, W.M.; Rinker, L.G.; Lessov, N.S.; Hazel, K.; Hill, J.K.; Stenzel-Poore, M.; Eckenstein, F. Lack of interleukin-6 expression is not protective against focal central nervous system ischemia. Stroke 2000, 31, 1715–1720. [Google Scholar] [CrossRef]
- Slivka, A.; Murphy, E.; Horrocks, L. Cerebral edema after temporary and permanent middle cerebral artery occlusion in the rat. Stroke 1995, 26, 1061–1065; discussion 5–6. [Google Scholar] [CrossRef]
- Shah, F.A.; Liu, G.; Al Kury, L.T.; Zeb, A.; Abbas, M.; Li, T.; Yang, X.; Liu, F.; Jiang, Y.; Li, S.; et al. Melatonin Protects MCAO-Induced Neuronal Loss via NR2A Mediated Prosurvival Pathways. Front. Pharmacol. 2019, 10, 297. [Google Scholar] [CrossRef]
- Lucke-Wold, B.P.; Smith, K.E.; Nguyen, L.; Turner, R.C.; Logsdon, A.F.; Jackson, G.J.; Huber, J.D.; Rosen, C.L.; Miller, D.B. Sleep disruption and the sequelae associated with traumatic brain injury. Neurosci. Biobehav. Rev. 2015, 55, 68–77. [Google Scholar] [CrossRef]
- Motwani, K.; Dodd, W.S.; Laurent, D.; Lucke-Wold, B.; Chalouhi, N. Delayed cerebral ischemia: A look at the role of endothelial dysfunction, emerging endovascular management, and glymphatic clearance. Clin. Neurol. Neurosurg. 2022, 218, 107273. [Google Scholar] [CrossRef]
- Sánchez-Barceló, E.J.; Mediavilla, M.D.; Tan, D.X.; Reiter, R.J. Clinical uses of melatonin: Evaluation of human trials. Curr. Med. Chem. 2010, 17, 2070–2095. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Neurologic Behavioral Score | |||
|---|---|---|---|
| Sensory | Motor | 28-Point Clinical Scale | |
| Vehicle-treated (n = 13) | 2.0 (1.60–2.40) | 2.5 (1.99–3.01) | 12.5 (9.95–15.05) |
| Melatonin-treated (n = 12) | 1.0 (0.55–1.45) | 1.0 (0.9–1.1) * | 7.5 (6.64–8.36) * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, A.-H.; Tai, S.-H.; Huang, S.-Y.; Chang, L.-D.; Chen, L.-Y.; Chen, Y.-N.; Hsu, H.-H.; Lee, E.-J. Melatonin Improves Vasogenic Edema via Inhibition to Water Channel Aquaporin-4 (AQP4) and Metalloproteinase-9 (MMP-9) Following Permanent Focal Cerebral Ischemia. Biomedicines 2024, 12, 2184. https://doi.org/10.3390/biomedicines12102184
Lee A-H, Tai S-H, Huang S-Y, Chang L-D, Chen L-Y, Chen Y-N, Hsu H-H, Lee E-J. Melatonin Improves Vasogenic Edema via Inhibition to Water Channel Aquaporin-4 (AQP4) and Metalloproteinase-9 (MMP-9) Following Permanent Focal Cerebral Ischemia. Biomedicines. 2024; 12(10):2184. https://doi.org/10.3390/biomedicines12102184
Chicago/Turabian StyleLee, Ai-Hua, Shih-Huang Tai, Sheng-Yang Huang, Li-Der Chang, Liang-Yi Chen, Yu-Ning Chen, Hao-Hsiang Hsu, and E-Jian Lee. 2024. "Melatonin Improves Vasogenic Edema via Inhibition to Water Channel Aquaporin-4 (AQP4) and Metalloproteinase-9 (MMP-9) Following Permanent Focal Cerebral Ischemia" Biomedicines 12, no. 10: 2184. https://doi.org/10.3390/biomedicines12102184