
Diabetes Mellitus and Pregnancy: An Insight into the Effects on the Epigenome

Abstract
:1. Introduction
2. Methods
3. Results
3.1. Epigenetics as the Possible Origin of Diabetes during Gestation: Experimental Models and Studies in Humans
3.1.1. Experimental Models
3.1.2. Studies in Humans
3.2. DNA Methylation in Diabetic Pregnancies: Cellular and Animal Models, and Studies in Humans
3.2.1. Cell Models
3.2.2. Animal Models
3.2.3. Studies in Humans
3.3. Acetylation, Methylation, and Other Epigenetic Modifications in Histones: Cell and Animal Models and Studies in Humans
3.3.1. Cell Models
3.3.2. Animal Models
3.3.3. Studies in Humans
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- ElSayed, N.; Aleppo, G.; Aroda, V.; Bannuru, R.; Brown, F.; Bruemmer, D.; Collins, B.; Hilliard, M.; Isaacs, D.; Johnson, E.; et al. 15. Management of Diabetes in Pregnancy: Standards of Care in Diabetes—2023. Diabetes Care 2023, 46, S254–S266. [Google Scholar] [CrossRef] [PubMed]
- International Diabetes Federation. IDF Diabetes Atlas, 10th ed.; International Diabetes Federation: Brussels, Belgium, 2021. [Google Scholar]
- Luo, P.; Fan, Y.; Zhang, C.; Yang, Z.; Sun, F.; Mei, B. Metabolic Characteristics of Gestational Diabetes Mellitus and the Effects on Pregnancy Outcomes. Diabetes Metab. Syndr. Obes. 2023, 16, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Rassie, K.; Giri, R.; Joham, A.; Teede, H.; Mousa, A. Human Placental Lactogen in Relation to Maternal Metabolic Health and Fetal Outcomes: A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2022, 23, 15621. [Google Scholar] [CrossRef]
- Holt, R.; DeVries, J.; Hess-Fischl, A.; Hirsch, I.; Kirkman, M.; Klupa, T.; Ludwig, B.; Nørgaard, K.; Pettus, J.; Renard, E.; et al. The management of type 1 diabetes in adults. A consensus report by the American Diabetes Association (ADA) and the European Association for the Study of Diabetes (EASD). Diabetologia 2021, 64, 2609–2652. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Cao, L.; Reece, E.; Zhao, Z. Impact of protein O-GlcNAcylation on neural tube malformation in diabetic embryopathy. Sci. Rep. 2017, 7, 11107. [Google Scholar] [CrossRef]
- Pearce, E. Introduction to Endocrine Disorders in Pregnancy. In A Case-Based Guide to Clinical Endocrinology, 3rd ed.; Davies, T., Ed.; Springer: New York, NY, USA, 2022; pp. 303–305. [Google Scholar]
- Brown, H.; Green, E.; Tan, T.; Gonzalez, M.; Rumbold, A.; Hull, M.; Norman, R.; Packer, N.; Robertson, S.; Thompson, J. Periconception onset diabetes is associated with embryopathy and fetal growth retardation, reproductive tract hyperglycosylation and impaired immune adaptation to pregnancy. Sci. Rep. 2018, 8, 2114. [Google Scholar] [CrossRef]
- Franzago, M.; Fraticelli, F.; Stuppia, L.; Vitacolonna, E. Nutrigenetics, epigenetics and gestational diabetes: Consequences in mother and child. Epigenetics 2019, 14, 215–235. [Google Scholar] [CrossRef]
- Ding, M.; Chavarro, J.; Olsen, S.; Lin, Y.; Ley, S.; Bao, W.; Rawal, S.; Grunnet, L.; Thuesen, A.; Mills, J.; et al. Genetic variants of gestational diabetes mellitus: A study of 112 SNPs among 8722 women in two independent populations. Diabetologia 2018, 61, 1758–1768. [Google Scholar] [CrossRef]
- Elliot, H.; Sharp, G.; Relton, C.; Lawlor, D. Epigenetics and gestational diabetes: A review of epigenetic epidemiology studies and their use to explore epigenetic mediation and improve prediction. Diabetologia 2019, 62, 2171–2178. [Google Scholar] [CrossRef]
- Zuccarello, D.; Sorrentino, U.; Brasson, V.; Marin, L.; Piccolo, C.; Capalbo, A.; Andrisani, A.; Cassina, M. Epigenetics of pregnancy: Looking beyond the DNA code. J. Assist. Reprod. Genet. 2022, 39, 801–816. [Google Scholar] [CrossRef]
- Agarwal, P.; Morriseau, T.; Kereliuk, S.; Doucette, C.; Wicklow, B.; Dolinsky, V. Maternal obesity, diabetes during pregnancy and epigenetic mechanisms that influence the developmental origins of cardiometabolic disease in the offspring. Crit. Rev. Clin. Lab. Sci. 2018, 55, 71–101. [Google Scholar] [CrossRef]
- Ma, R.; Tutino, G.; Lillycrop, K.; Hanson, M.; Tam, W. Maternal diabetes, gestational diabetes and the role of epigenetics in their long-term effects on offspring. Prog. Biophys. Mol. Biol. 2015, 118, 55–68. [Google Scholar] [CrossRef]
- Pinney, S.; Simmons, R. Epigenetic mechanisms in the development of type 2 diabetes. Trends Endocrinol. Metab. 2010, 21, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Pirola, L.; Balcerczyk, A.; Okabe, J.; El-Osta, A. Epigenetic phenomena linked to diabetic complications. Nat. Rev. Endocrinol. 2010, 6, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Smallwood, S.; Kelsey, G. De novo DNA methylation: A germ cell perspective. Trends Genet. 2012, 28, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Salbaum, J.; Jones, S.; Burk, D.; Kappen, C. Aberrant lipid accumulation in the mouse visceral yolk sac resulting from maternal diabetes and obesity. Front. Cell Dev. Biol. 2023, 11, 1073807. [Google Scholar] [CrossRef]
- Bequer, L.; Gómez, T.; Molina, J.; Álvarez, A.; Chaviano, C.; Clapés, S. Experimental diabetes impairs maternal reproductive performance in pregnant Wistar rats and their offspring. Syst. Biol. Reprod. Med. 2018, 64, 60–70. [Google Scholar] [CrossRef]
- Dong, M.; Li, Q.; Fan, L.; Li, L.; Shen, W.; Wang, Z.; Sun, Q. Diabetic Uterine Environment Leads to Disorders in Metabolism of Offspring. Front. Cell Dev. Biol. 2021, 9, 706879. [Google Scholar] [CrossRef]
- Bueno, A.; Sinzato, Y.; Volpato, G.; Gallego, F.; Perecin, F.; Rodrigues, T.; Damasceno, D. Severity of pre-pregnancy diabetes on the fetal malformations and viability associated with early embryos in rats. Biol. Reprod. 2020, 103, 938–950. [Google Scholar] [CrossRef]
- Shao, W.; Tao, L.; Gao, C.; Xie, J.; Zhao, R. Alterations in methylation and expression levels of imprinted genes H19 and Igf2 in the fetuses of diabetic mice. Comp. Med. 2008, 58, 341–346. [Google Scholar]
- Ding, G.; Wang, F.; Shu, J.; Tian, S.; Jiang, Y.; Zhang, D.; Wang, N.; Luo, Q.; Zhang, Y.; Jin, F.; et al. Transgenerational glucose intolerance with Igf2/H19 epigenetic alterations in mouse islet induced by intrauterine hyperglycemia. Diabetes 2012, 61, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Stoffers, D.; Nicholls, R.; Simmons, R. Development of type 2 diabetes following intrauterine growth. J. Clin. Investig. 2008, 118, 2316–2324. [Google Scholar] [PubMed]
- Gonçalves, V.; Sinzato, Y.; Queiroz de Moraes-Souza, R.; Sousa, T.; Gallego, F.; Karki, B.; de Andrade, A.; Corrente, J.; Damasceno, D.; Volpato, G. Metabolic changes in female rats exposed to intrauterine hyperglycemia and postweaning consumption of high-fat diet. Biol. Reprod. 2022, 106, 200–212. [Google Scholar]
- Lecoutre, S.; Oger, F.; Pourpe, C.; Butruille, L.; Marousez, L.; Dickes-Coopman, A.; Laborie, C.; Guinez, C.; Lesage, J.; Vieau, D.; et al. Maternal obesity programs increased leptin gene expression in rat male offspring via epigenetic modifications in a depot-specific manner. Mol. Metab. 2017, 6, 922–930. [Google Scholar] [CrossRef] [PubMed]
- Vucetic, Z.; Kimmel, J.; Totoki, K.; Hollenbeck, E.; Reyes, T. Maternal High-Fat Diet Alters Methylation and Gene Expression of Dopamine and Opioid-Related Genes. Endocrinology 2010, 151, 4754–4764. [Google Scholar] [CrossRef]
- Ramamoorthy, T.; Allen, T.; Davies, A.; Harno, E.; Sefton, C.; Murgatroyd, C.; White, W. Maternal overnutrition programs epigenetic changes in the regulatory regions of hypothalamic Pomc in the offspring of rats. Int. J. Obes. 2018, 42, 1431–1444. [Google Scholar] [CrossRef]
- Taqui, B.; Asadi, F.; Capobianco, E.; Hardy, D.; Jawerbaum, A.; Arany, E. Addition of olive oil to diet of rats with mild pre-gestational diabetes impacts offspring β-cell development. J. Endocrinol. 2020, 246, 175–187. [Google Scholar] [CrossRef]
- Finer, S.; Saravanan, P.; Hitman, G.; Yajnik, C. The role of the one-carbon cycle in the developmental origins of Type 2 diabetes and obesity. Diabet. Med. 2014, 31, 263–272. [Google Scholar] [CrossRef]
- Sideratou, T.; Atkinson, F.; Campbell, G.; Petocz, P.; Bell-Anderson, K.; Brand-Miller, J. Glycaemic Index of Maternal Dietary Carbohydrate Differentially Alters Fto and Lep Expression in Offspring in C57BL/6 Mice. Nutrients 2018, 10, 1342. [Google Scholar] [CrossRef]
- Blasetti, A.; Quarta, A.; Guarino, M.; Cicolini, I.; Iannucci, D.; Giannini, C.; Chiarelli, F. Role of Prenatal Nutrition in the Development of Insulin Resistance in Children. Nutrients 2023, 15, 87. [Google Scholar] [CrossRef]
- Gómez, T.; Bequer, L.; Molineda, A.; Molina, J.; Álvarez, A.; Lavastida, M.; Cruz, G.; Freire, C.; Clapés, S. Benefits of zinc supplementation on the metabolic, redox and trace elements status in mild diabetic rats. J. Pharm. Pharmacogn. Res. 2019, 7, 144–155. [Google Scholar]
- Wilson, R.; Leemaqz, S.; Goh, Z.; McAninch, D.; Jankovic-Karasoulos, T.; Leghi, G.; Phillips, J.; Colafella, K.; Tran, C.; O’Leary, S.; et al. Zinc is a critical regulator of placental morphogenesis and maternal hemodynamics during pregnancy in mice. Sci. Rep. 2017, 7, 15137. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, K.; Allegrucci, C.; Singh, R.; Gardner, D.; Sebastian, S.; Bispham, J.; Thurston, A.; Huntley, J.; Rees, W.; Maloney, C.; et al. DNA methylation, insulin resistance, and blood pressure in offspring determined by maternal periconceptional B vitamin and methionine status. Proc. Natl. Acad. Sci. USA 2007, 104, 19351–19356. [Google Scholar] [CrossRef] [PubMed]
- Hatchwell, E.; Greally, J. The potential role of epigenomic dysregulation in complex human disease. Trends Genet. 2007, 23, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Feil, R.; Fraga, M. Epigenetics and the environment: Emerging patterns and implications. Nat. Rev. Genet. 2012, 13, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Geurtsen, M.; van Soest, E.; Voerman, E.; Steegers, E.; Jaddoe, V.; Gaillard, R. High maternal early-pregnancy blood glucose levels are associated with altered fetal growth and increased risk of adverse birth outcomes. Diabetologia 2019, 62, 1880–1890. [Google Scholar] [CrossRef]
- Wahab, R.; Voerman, E.; Jansen, P.; Oei, E.; Steegers, E.; Jaddoe, V.; Gaillard, R. Maternal glucose concentrations in early pregnancy and cardiometabolic risk factors in childhood. Obesity 2020, 28, 985–993. [Google Scholar] [CrossRef]
- Cafiero, P.; Krochik, G. Maternal diabetes and neurodevelopmental disorders in offspring. Medicina 2020, 80, 685–695. [Google Scholar]
- Bouchard, L.; Hivert, M.; Guay, S.; St-Pierre, J.; Perron, P.; Brisson, D. Placental Adiponectin Gene DNA Methylation Levels Are Associated with Mothers’ Blood Glucose Concentration. Diabetes 2012, 61, 1272–1280. [Google Scholar] [CrossRef]
- Bouchard, L.; Thibault, S.; Guay, S.; Santure, M.; Monpetit, A.; St-Pierre, J.; Perron, P.; Brisson, D. Leptin gene epigenetic adaptation to impaired glucose metabolism during pregnancy. Diabetes Care 2010, 33, 2436–2441. [Google Scholar] [CrossRef]
- Anand, S.; Gupta, M.; Teo, K.; Schulze, K.; Desai, D.; Abdalla, N.; Zulyniak, M.; de Souza, R.; Wahi, G.; Shaikh, M.; et al. Causes and consequences of gestational diabetes in South Asians living in Canada: Results from a prospective cohort study. Can. Med. Assoc. J. 2017, 5, E604–E611. [Google Scholar] [CrossRef] [PubMed]
- Yajnik, C.; Deshpande, S.; Jackson, A.; Refsum, H.; Rao, S.; Fisher, D.; Bhat, D.; Naik, S.; Coyaji, K.; Joglekar, C.; et al. Vitamin B12 and folate concentrations during pregnancy and insulin resistance in the offspring: The Pune Maternal Nutrition Study. Diabetologia 2008, 51, 29–38. [Google Scholar] [CrossRef]
- Heijmans, B.; Tobi, E.; Stein, A.; Putter, H.; Blauw, G.; Susser, E.; Slagboom, P.; Lume, L. Persistent epigenetic differences associated with prenatal exposure to famine in humans. Proc. Natl. Acad. Sci. USA 2008, 105, 17046–17049. [Google Scholar] [CrossRef]
- Tobi, E.; Lumey, L.; Talens, R.; Kremer, D.; Putter, H.; Stein, A.; Slagboom, P.; Heijmans, B. DNA methylation differences after exposure to prenatal famine are common and timing- and sex-specific. Hum. Mol. Genet. 2009, 18, 4046–4053. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, K.; Sheppard, A.; Gluckman, P.; Lillycrop, K.; Burdge, G.; McLean, C.; Rodford, J.; Slater-Jefferies, J.; Garratt, E.; Crozier, S.; et al. Epigenetic gene promoter methylation at birth is associated with child’s later adiposity. Diabetes 2011, 60, 1528–1534. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, M. Genomics, type 2 diabetes, and obesity. N. Engl. J. Med. 2010, 363, 2339–2350. [Google Scholar] [CrossRef] [PubMed]
- Cvitic, S.; Novakovic, B.; Gordon, L.; Ulz, C.; Mühlberger, M.; Diaz-Perez, F.; Joo, J.; Svendova, V.; Schimek, M.; Trajanoski, S.; et al. Human fetoplacental arterial and venous endothelial cells are differentially programmed by gestational diabetes mellitus, resulting in cell-specific barrier function changes. Diabetologia 2018, 61, 2398–2411. [Google Scholar] [CrossRef] [PubMed]
- Howe, C.; Cox, B.; Fore, R.; Jungius, J.; Kvist, T.; Lent, S.; Miles, H.; Salas, L.; Rifas-Shiman, S.; Starling, A.; et al. Maternal Gestational Diabetes Mellitus and Newborn DNA Methylation: Findings From the Pregnancy and Childhood Epigenetics Consortium. Diabetes Care 2020, 43, 98–105. [Google Scholar] [CrossRef]
- Haertle, L.; El Hajj, N.; Dittrich, M.; Müller, T.; Nanda, I.; Lehnen, H.; Haaf, T. Epigenetic signatures of gestational diabetes mellitus on cord blood methylation. Clin. Epigenet. 2017, 9, 28. [Google Scholar] [CrossRef]
- Tam, W.; Ma, R.; Yang, X.; Ko, G.; Tong, P.; Cockram, C.; Sahota, D.; Rogers, M.; Chan, J. Glucose intolerance and cardiometabolic risk in children exposed to maternal gestational diabetes mellitus in utero. Pediatrics 2008, 122, 1229–1234. [Google Scholar] [CrossRef]
- Hromadnikova, I.; Kotlabova, K.; Dvorakova, L.; Krofta, L.; Sirc, J. Substantially Altered Expression Profile of Diabetes/Cardiovascular/Cerebrovascular Disease Associated microRNAs in Children Descending from Pregnancy Complicated by Gestational Diabetes Mellitus-One of Several Possible Reasons for an Increased Cardiovascular Risk. Cells 2020, 9, 1557. [Google Scholar] [PubMed]
- Ruchat, S.; Houde, A.; Voisin, G.; St-Pierre, J.; Perron, P.; Baillargeon, J.; Gaudet, D.; Hivert, M.; Brisson, D.; Bouchar, L. Gestational diabetes mellitus epigenetically affects genes predominantly involved in metabolic diseases. Epigenetics 2013, 8, 935–943. [Google Scholar] [CrossRef]
- Yang, I.; Zhang, W.; Davidson, J.; Fingerlin, T.; Kechris, K.; Dabelea, D. Epigenetic marks of in utero exposure to gestational diabetes and childhood adiposity outcomes: The EPOCH study. Diabet. Med. 2018, 35, 612–620. [Google Scholar] [CrossRef] [PubMed]
- Hillier, T.; Pedula, K.; Schmidt, M.; Mullen, J.; Charles, M.; Pettitt, D. Childhood obesity and metabolic imprinting: The ongoing effects of maternal hyperglycemia. Diabetes Care 2007, 30, 2287–2292. [Google Scholar] [CrossRef] [PubMed]
- Clausen, T.; Mathiesen, E.; Hansen, T.; Pedersen, O.; Jensen, D.; Lauenborg, J.; Schmidt, L.; Damm, P. Overweight and the metabolic syndrome in adult offspring of women with diet-treated gestational diabetes mellitus or type 1 diabetes. J. Clin. Endocrinol. Metab. 2009, 94, 2464–2470. [Google Scholar] [CrossRef] [PubMed]
- Gagné-Ouellet, V.; Houde, A.; Guay, S.; Perron, P.; Gaudet, D.; Guérin, R.; Baillargeon, J.; Hivert, M.; Brisson, D.; Bouchard, L. Placental lipoprotein lipase DNA methylation alterations are associated with gestational diabetes and body composition at 5 years of age. Epigenetics 2017, 12, 616–662. [Google Scholar] [CrossRef] [PubMed]
- Rancourt, R.; Ott, R.; Schellong, K.; Melchior, K.; Ziska, T.; Henrich, W.; Plagemann, A. Visceral adipose tissue alteration of PI3KR1 expression is associated with gestational diabetes but not promoter DNA methylation. Adipocyte 2019, 8, 339–346. [Google Scholar] [CrossRef]
- Guillemette, L.; Wicklow, B.; Sellers, E.; Dart, A.; Shen, G.; Dolinsky, V.; Gordon, J.; Jassal, D.; Nickel, N.; Duhamel, T.; et al. Intrauterine exposure to diabetes and risk of cardiovascular disease in adolescence and early adulthood: A population-based birth cohort study. Can. Med. Assoc. J. 2020, 192, E1104–E1113. [Google Scholar] [CrossRef]
- Hoodbhoy, Z.; Mohammed, N.; Nathani, K.; Sattar, S.; Chowdhury, D.; Maskatia, S.; Tierney, S.; Hasan, B.; Das, J. The Impact of Maternal Preeclampsia and Hyperglycemia on the Cardiovascular Health of the Offspring: A Systematic Review and Meta-Analysis. Am. J. Perinatol. 2023, 40, 363–374. [Google Scholar] [CrossRef]
- del Rosario, M.; Ossowski, V.; Knowler, W.; Bogardus, C.; Baier, L.; Hanson, R. Potential epigenetic dysregulation of genes associated with MODY and type 2 diabetes in humans exposed to a diabetic intrauterine environment: An analysis of genome-wide DNA methylation. Metabolism 2014, 63, 654–660. [Google Scholar] [CrossRef]
- Alekseenkova, E.; Babakov, V.; Selkov, S.; Di Renzo, G.; Kogan, I.; Kapustin, R. Maternal insulin-like growth factors and insulin-like growth factor–binding proteins for macrosomia prediction in diabetic and nondiabetic pregnancy: A prospective study. Int. J. Gynecol. Obstet. 2023, 162, 605–613. [Google Scholar] [CrossRef]
- Zhang, T.; Huang, X.; Zhao, X.; Wang, W.; Wen, R.; Gao, S. Risks of specific congenital anomalies in offspring of women with diabetes: A systematic review and meta-analysis of population-based studies including over 80 million births. PLoS Med. 2022, 19, e1003900. [Google Scholar] [CrossRef] [PubMed]
- Fujikawa, K.; Waguri, M.; Takahara, M.; Katakami, N.; Shimomura, I. Trends in maternal characteristics and perinatal outcomes among Japanese pregnant women with type 1 and type 2 diabetes from 1982 to 2020. J. Diabetes Investig. 2022, 13, 1761–1770. [Google Scholar] [CrossRef] [PubMed]
- Roberti, A.; Fernández, A.; Fraga, M. Nicotinamide N-methyltransferase: At the crossroads between cellular metabolism and epigenetic regulation. Mol. Metab. 2021, 45, 101165. [Google Scholar] [CrossRef]
- Liu, J.; Deng, Z.; Zhu, X.; Zeng, Y.; Guan, X.; Li, J. Roles of Nicotinamide N-Methyltransferase in Obesity and Type 2 Diabetes. Biomed. Res. Int. 2021, 2021, 9924314. [Google Scholar] [CrossRef] [PubMed]
- Crujeiras, A.; Pissios, P.; Moreno-Navarrete, J.; Diaz-Lagares, A.; Sandoval, J.; Gomez, A.; Ricart, W.; Esteller, M.; Casanueva, F.; Fernandez-Real, J. An Epigenetic Signature in Adipose Tissue Is Linked to Nicotinamide N-Methyltransferase Gene Expression. Mol. Nutr. Food Res. 2018, 62, e1700933. [Google Scholar] [CrossRef] [PubMed]
- Ruf, S.; Rajagopal, S.; Kadnur, S.; Hallur, M.; Rani, S.; Kristam, R.; Swaminathan, S.; Zope, B.; Gondrala, P.; Swamy, I.; et al. Novel tricyclic small molecule inhibitors of Nicotinamide N-methyltransferase for the treatment of metabolic disorders. Sci. Rep. 2022, 12, 15440. [Google Scholar] [CrossRef] [PubMed]
- Kraus, D.; Yang, Q.; Kong, D.; Banks, A.; Zhang, L.; Rodgers, J.; Pirinen, E.; Pulinilkunnil, T.; Gong, F.; Wang, Y.; et al. Nicotinamide N-methyltransferase knockdown protects against diet-induced obesity. Nature 2014, 508, 258–262. [Google Scholar] [CrossRef]
- Dias, S.; Pheiffer, C.; Abrahams, Y.; Rheeder, P.; Adam, S. Molecular Biomarkers for Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2018, 19, 2926. [Google Scholar] [CrossRef]
- Kandilya, D.; Shyamasundar, S.; Singh, D.; Banik, A.; Hande, M.; Stünkel, W.; Chong, Y.; Dheen, S. High glucose alters the DNA methylation pattern of neurodevelopment associated genes in human neural progenitor cells in vitro. Sci. Rep. 2020, 10, 15676. [Google Scholar] [CrossRef]
- Ladd-Acosta, C.; Vang, E.; Barrett, E.; Bulka, C.; Bush, N.; Cardenas, A.; Dabelea, D.; Dunlop, A.; Fry, R.; Gao, X.; et al. Analysis of Pregnancy Complications and Epigenetic Gestational Age of Newborns. JAMA Netw. Open 2023, 6, e230672. [Google Scholar] [CrossRef]
- Hall, E.; Dekker, N.; Volkov, P.; Malmgren, S.; Mulder, H.; Bacos, K.; Ling, C. The effects of high glucose exposure on global gene expression and DNA methylation in human pancreatic islets. Mol. Cell Endocrinol. 2018, 472, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Cheng, Y.; Ming, Z.; Dong, X.; Zhou, Y.; Ding, G.; Pang, H.; Rahman, T.; Akbar, R.; Huang, H.; et al. Intrauterine hyperglycemia exposure results in intergenerational inheritance via DNA methylation reprogramming on F1 PGCs. Epigenet. Chromatin 2018, 11, 20. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Gong, L.; Zhang, P.; Li, Y.; Liu, B.; Zhang, L.; Zhuang, J.; Xiao, D. Epigenetic Down-Regulation of Sirt 1 via DNA Methylation and Oxidative Stress Signaling Contributes to the Gestational Diabetes Mellitus-Induced Fetal Programming of Heart Ischemia-Sensitive Phenotype in Late Life. Int. J. Biol. Sci. 2019, 15, 1240–1251. [Google Scholar] [CrossRef]
- Lister, R.; Chamberlain, A.; Einstein, F.; Wu, B.; Zheng, D.; Zhou, B. Intrauterine Programming of Diabetes Induced Cardiac Embryopathy. Diabetes Obes. Int. J. 2019, 4, 202. [Google Scholar] [CrossRef] [PubMed]
- Meng, R.; Song, J.; Guan, L.; Li, Q.; Shi, C.; Su, D.; Ma, X. Genome-wide analysis of methylation in rat fetal heart under hyperglycemia by methylation-dependent restriction site–associated DNA sequencing. PLoS ONE 2022, 17, e0268117. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Chen, X.; Xiao, Y.; Wen, J.; Chen, J.; Wang, K.; Chen, G. Gestational diabetes mellitus alters DNA methylation profiles in pancreas of the offspring mice. J. Diabetes Complicat. 2019, 33, 15–22. [Google Scholar] [CrossRef]
- Nazari, Z.; Shahryari, A.; Ghafari, S.; Nabiuni, M.; Golalipour, M. In Utero Exposure to Gestational Diabetes Alters DNA Methylation and Gene Expression of CDKN2A/B in Langerhans Islets of Rat Offspring. Cell J. 2020, 22, 203–211. [Google Scholar]
- Keleher, M.; Zaidi, R.; Shah, S.; Oakley, E.; Pavlatos, C.; Idrissi, S.; Xing, X.; Li, D.; Wang, T.; Cheverud, J. Maternal high-fat diet associated with altered gene expression, DNA methylation, and obesity risk in mouse offspring. PLoS ONE 2018, 13, e0192606. [Google Scholar] [CrossRef]
- Sinha, N.; Biswas, A.; Nave, O.; Seger, C.; Sen, A. Gestational Diabetes Epigenetically Reprograms the Cart Promoter in Fetal Ovary, Causing Subfertility in Adult Life. Endocrinology 2019, 160, 1684–1700. [Google Scholar] [CrossRef]
- Tabebordbar, M.; Sarabi, M.; Vakili, S.; Zare, R.; Zal, F. Effect of folic acid and vitamin E on promoter DNA methylation and expression of TGF-β1, ESR-1 and CDH-1 in the uterus of STZ-induced diabetic rats. Arch. Physiol. Biochem. 2022, 128, 1339–1345. [Google Scholar] [CrossRef] [PubMed]
- Houshmand-Oeregaard, A.; Hansen, N.; Hjort, L.; Kelstrup, L.; Broholm, C.; Mathiesen, E.; Clausen, T.; Damm, P.; Vaag, A. Differential adipokine DNA methylation and gene expression in subcutaneous adipose tissue from adult offspring of women with diabetes in pregnancy. Clin. Epigenet. 2017, 9, 37. [Google Scholar] [CrossRef] [PubMed]
- Hjort, L.; Martino, D.; Grunnet, L.; Naeem, H.; Maksimovic, J.; Olsson, A.; Zhang, C.; Ling, C.; Olsen, S.; Saffery, R.; et al. Gestational diabetes and maternal obesity are associated with epigenome-wide methylation changes in children. JCI Insight 2018, 3, e122572. [Google Scholar] [CrossRef] [PubMed]
- Ott, R.; Stupin, J.; Melchior, K.; Schellong, K.; Ziska, T.; Dudenhausen, J.; Henrich, W.; Rancourt, R.; Plagemann, A. Alterations of adiponectin gene expression and DNA methylation in adipose tissues and blood cells are associated with gestational diabetes and neonatal outcome. Clin. Epigenet. 2018, 10, 131. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Lee, K.; Byeon, E.; Choi, J.; Kim, S.; Shin, J. Maternal Gestational Diabetes Influences DNA Methylation in the Serotonin System in the Human Placenta. Life 2022, 12, 1869. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.; Teague, A.; Chen, J.; Aston, C.; Leung, Y.; Chernausek, S.; Simmons, R.; Pinney, S. Offspring sex impacts DNA methylation and gene expression in placentae from women with diabetes during pregnancy. PLoS ONE 2018, 13, e0190698. [Google Scholar] [CrossRef] [PubMed]
- Schulze, K.; Bhatt, A.; Azamian, M.; Sundgren, N.; Zapata, G.; Hernandez, P.; Fox, K.; Kaiser, J.; Belmont, J.; Hanchard, N. Aberrant DNA methylation as a diagnostic biomarker of diabetic embryopathy. Genet. Med. 2019, 21, 2453–2461. [Google Scholar] [CrossRef]
- Shiau, S.; Wang, L.; Liu, H.; Zheng, Y.; Drong, A.; Joyce, B.T.; Wang, J.; Li, W.; Leng, J.; Shen, Y.; et al. Prenatal gestational diabetes mellitus exposure and accelerated offspring DNA methylation age in early childhood. Epigenetics 2021, 16, 186–195. [Google Scholar] [CrossRef]
- Taschereau, A.; Thibeault, K.; Allard, C.; Juvinao-Quintero, D.; Perron, P.; Lutz, S.; Bouchard, L.; Hivert, M. Maternal glycemia in pregnancy is longitudinally associated with blood DNAm variation at the FSD1L gene from birth to 5 years of age. Clin. Epigenet. 2023, 15, 107. [Google Scholar] [CrossRef]
- Juvinao-Quintero, D.; Starling, A.; Cardenas, A.; Powe, C.; Perron, P.; Bouchard, L.; Dabalea, D.; Hivert, M. Epigenome-wide association study of maternal hemoglobin A1c in pregnancy and cord blood DNA methylation. Epigenomics 2021, 13, 3. [Google Scholar] [CrossRef]
- Manitta, E.; Fontes, I.; Stokholm, S.; Kelstrup, L.; Houshmand-Oeregaard, A.; Dalsgaard, T.; Groth, L.; Reinhardt, E.; Torp, L.; Barrès, R.; et al. DNA Methylation and Gene Expression in Blood and Adipose Tissue of Adult Offspring of Women with Diabetes in Pregnancy—A Validation Study of DNA Methylation Changes Identified in Adolescent Offspring. Biomedicines 2022, 10, 1244. [Google Scholar] [CrossRef]
- Dluski, D.; Wolinska, E.; Skrzypczak, M. Epigenetic Changes in Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2021, 22, 7649. [Google Scholar] [CrossRef]
- Di Pietrantonio, N.; Shumliakivska, M.; Suades, R.; Di Tomo, P.; Bonfini, T.; Pandolfi, A.; Consentino, F. Epigenetic regulation of oxidative and inflammatory phenotypes in women with gestational diabetes and offspring. Eur. Heart J. 2021, 42, 3346. [Google Scholar] [CrossRef]
- Hepp, P.; Hutter, S.; Knabl, J.; Hofmann, S.; Kuhn, C.; Mahner, S.; Jeschke, U. Histone H3 Lysine 9 Acetylation is Downregulated in GDM Placentas and Calcitriol Supplementation Enhanced This Effect. Int. J. Mol. Sci. 2018, 19, 4061. [Google Scholar] [CrossRef] [PubMed]
- Kuo, T.; Damle, M.; González, B.; Egli, D.; Lazar, M.; Accili, D. Induction of α cell–restricted Gc in dedifferentiating β cells contributes to stress-induced β cell dysfunction. JCI Insight 2019, 4, e128351. [Google Scholar] [CrossRef] [PubMed]
- Vanderkruk, B.; Maeshima, N.; Pasula, D.; An, M.; McDonald, C.; Suresh, P.; Luciani, D.; Lynn, F.; Hoffman, B. Methylation of histone H3 lysine 4 is required for maintenance of beta cell function in adult mice. Diabetologia 2023, 66, 1097–1115. [Google Scholar] [PubMed]
- Zhang, Q.; Cai, T.; Xiao, Z.; Li, D.; Wan, C.; Cui, X.; Bai, B. Identification of histone malonylation in the human fetal brain and implications for diabetes-induced neural tube defects. Mol. Genet. Genom. Med. 2020, 8, e1404. [Google Scholar] [CrossRef]
- Kusuyama, J.; Makarewicz, N.; Albertson, B.; Alves-Wagner, A.; Conlin, R.; Prince, N.; Alves, C.; Ramachandran, K.; Kozuka, C.; Xiudong, Y.; et al. Maternal Exercise-Induced SOD3 Reverses the Deleterious Effects of Maternal High Fat Diet on Offspring Metabolism Through Stabilization of H3K4me3 and Protection Against WDR82 Carbonylation. Diabetes 2022, 71, 1170–1181. [Google Scholar]
- Upadhyaya, B.; Larsen, T.; Barwari, S.; Louwagie, E.; Baack, M.; Dey, M. Prenatal Exposure to a Maternal High-Fat Diet Affects Histone Modification of Cardiometabolic Genes in Newborn Rats. Nutrients 2017, 9, 407. [Google Scholar] [CrossRef]
- Glendining, K.; Jasoni, C. Maternal High Fat Diet-Induced Obesity Modifies Histone Binding and Expression of Oxtr in Offspring Hippocampus in a Sex-Specific Manner. Int. J. Mol. Sci. 2019, 20, 329. [Google Scholar] [CrossRef]
- Ji, S.; Zhou, W.; Li, X.; Liu, S.; Wang, F.; Li, X.; Zhao, T.; Ji, G.; Du, J.; Hao, A. Maternal hyperglycemia disturbs neocortical neurogenesis via epigenetic regulation in C57BL/6J mice. Cell Death Dis. 2019, 10, 211. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Shen, W.; Reece, A.; Hasuwa, H.; Harman, C.; Kaushal, S.; Yang, P. Maternal diabetes induces senescence and neural tube defects sensitive to the senomorphic rapamycin. Sci. Adv. 2021, 7, eabf5089. [Google Scholar] [CrossRef]
- Hivert, M.; Cardenas, A.; Allard, C.; Doyon, M.; Powe, C.; Catalano, P.; Perron, P.; Bouchard, L. Interplay of Placental DNA Methylation and Maternal Insulin Sensitivity in Pregnancy. Diabetes 2020, 69, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Teague, A.; Tryggestad, J.; Jensen, M.; Chernausek, S. Role of metformin in epigenetic regulation of placental mitochondrial biogenesis in maternal diabetes. Sci. Rep. 2020, 10, 8314. [Google Scholar] [CrossRef] [PubMed]
- Kopylov, A.; Papysheva, O.; Gribova, I.; Kaysheva, A.; Kotaysch, G.; Kharitonova, L.; Mayatskaya, T.; Nurbekov, M.; Schipkova, E.; Terekhina, O.; et al. Severe types of fetopathy are associated with changes in the serological proteome of diabetic mothers. Medicine 2021, 100, e27829. [Google Scholar] [CrossRef] [PubMed]
- Bhushan, R.; Rani, A.; Ali, A.; Singh, V.; Dubey, P. Bioinformatics enrichment analysis of genes and pathways related to maternal type 1 diabetes associated with adverse fetal outcomes. J. Diabetes Complicat. 2020, 34, 107556. [Google Scholar]
- Qu, X.; Yu, H.; Jia, B.; Yu, X.; Cui, Q.; Liu, Z.; Sun, C.; Chu, Y. Association of downregulated HDAC 2 with the impaired mitochondrial function and cytokine secretion in the monocytes/macrophages from gestational diabetes mellitus patients. Cell Biol. Int. 2016, 40, 642–651. [Google Scholar] [CrossRef]
- Antoun, E.; Kitaba, N.; Titcombe, P.; Dalrymple, K.; Garratt, E.; Barton, S.; Murray, R.; Seed, P.; Holbrook, J.; Kobor, M.; et al. Maternal dysglycaemia, changes in the infant’s epigenome modified with a diet and physical activity intervention in pregnancy: Secondary analysis of a randomised control trial. PLoS Med. 2020, 17, e1003229. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Studies in Animals | |||||
| Experimental Model | Epigenetic Modification | Gene | Expression Effect | Tissue or Cell | Reference |
| High-fat diet (HFD) | Hypomethylation and increased hydroxymethylation | LEP | Increased | Mouse plasma and fat pads | [26] |
| Hypomethylation | Slc6a3 | Increased | Mouse brains | [27] | |
| Oprm1 | |||||
| Penk | |||||
| Hypermethylation | Pomc | Unaltered | Rats exposed to HFD before and after birth | [28] | |
| Reduced overall variation in DNA methylation | N/S | Changes in the expression of 3908 genes | Mouse liver tissue | [81] | |
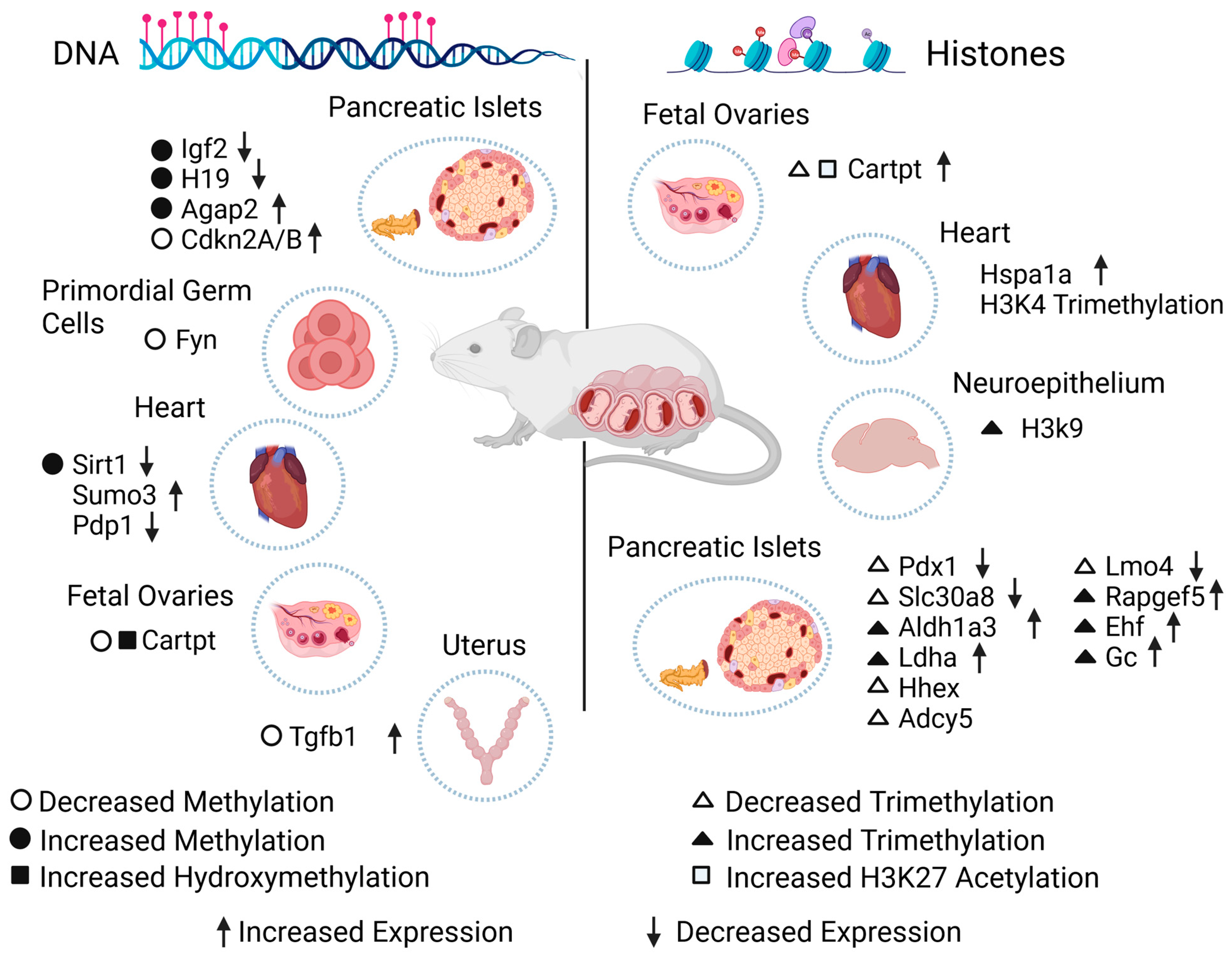
| GDM | Hypermethylation | Igf2 | Downregulated | Mouse pancreatic islets and sperm | [23] |
| H19 | |||||
| Decreased methylation | Fyn | N/S | Primordial germ cells from male mice | [75] | |
| Global DNA hypermethylation | Sirt1 | Downregulated | Rat hearts | [76] | |
| Abnormal methylation | Sumo3 | Increased | Genomic DNA of fetal heart tissue | [78] | |
| Pdp1 | Decreased | ||||
| Hypermethylation | Agap2 | Upregulated | Mouse pancreas | [79] | |
| Hypomethylation | Cdkn2A/B promoter | Upregulated | Rat pancreatic islets | [80] | |
| Hypomethylation and increased hydroxymethylation | Cartpt promoter | No changes compared to controls | Mouse fetal ovaries | [82] | |
| Undefined diabetes | Hypomethylation | Tgfb1 | Increased | Rat uterus | [83] |
| PGDM | Hypermethylation | 21 genes, see reference [73] | Decreased | Embryonic mouse hearts | [77] |
| Studies in Humans | |||||
| Epigenetic Modification | Gene | Expression Effect | Tissue or Cell | Reference | |
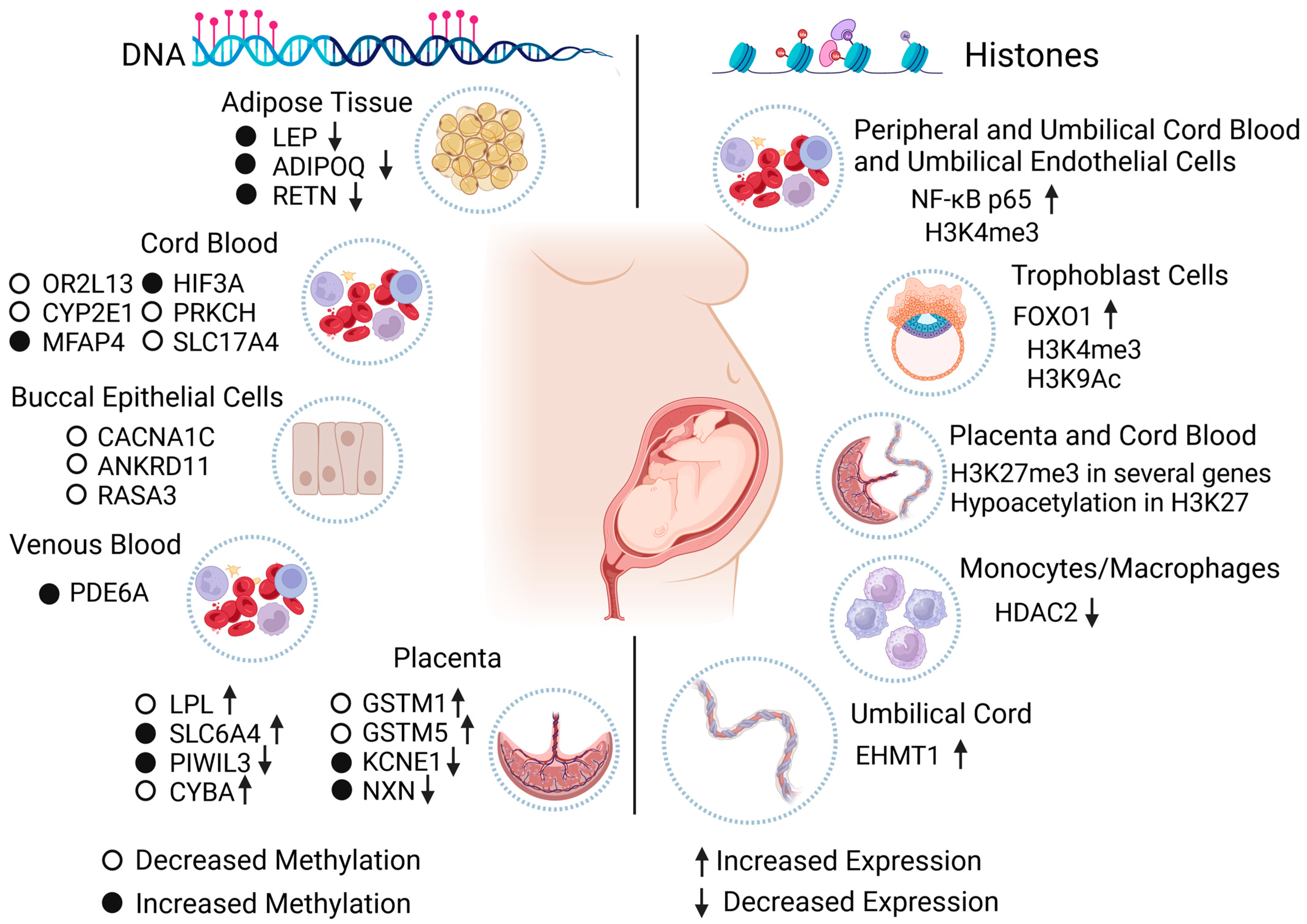
| Impaired glucose tolerance (IGT) | Hypermethylation | ADIPOQ | Decreased | Maternal side of placenta samples | [41] |
| Hypermethylation in 20 fetal and 26 maternal CpG sites | LEP | Decreased | Placental tissue from fetal and maternal sides | [42] | |
| Hyperglycemia | Hypomethylation and hypermethylation in YAP | SLIT1 | Downregulated | Human fetal neural progenitor cells | [72] |
| ROBO2 | |||||
| TAZ | |||||
| YAP | |||||
| Altered methylation in one or more CpG sites | RASD1 | Increased | Human pancreatic islets | [74] | |
| GLRA1 | Decreased | ||||
| VAC14 | Decreased | ||||
| SLCO5A1 | Increased | ||||
| CHRNA5 | Increased | ||||
| GDM | Hypomethylation | OR2L13 (Promoter) | N/S | Cord blood | [50] |
| CYP2E1 (Gene body) | |||||
| Hypermethylation | MFAP4 | N/S | Fetal cord blood | [51] | |
| Hypermethylation | HIF3A | ||||
| Hypomethylation | PRKCH | ||||
| Hypomethylation | SLC17A4 | ||||
| Hypomethylation | LPL | Increased | Placenta samples | [58] | |
| Increased methylation | LEP and ADIPOQ | Decreased | Abdominal subcutaneous adipose tissue of adult offspring | [84] | |
| Hypermethylation | PDE6A (cg09109411) | N/S | Offspring venous blood | [85] | |
| Moderate to higher methylation | ADIPOQ | Decreased | Maternal subcutaneous and visceral adipose tissue and maternal and fetal blood | [86] | |
| Hypermethylation | SLC6A4 | Increased | Fetal placenta side | [87] | |
| Diabetes in pregnancy | Hypermethylation | PIWIL3 | Decreased | Fetal placenta side | [88] |
| Hypomethylation | CYBA | Increased | |||
| Hypomethylation | GSTM1 | Increased | |||
| Hypomethylation | GSTM5 | Increased | |||
| Hypermethylation | KCNE1 | Decreased | |||
| Hypermethylation | NXN | Decreased | |||
| Global DNA hypomethylation | CACNA1C ANKRD11 RASA3 | N/S | Offspring buccal epithelial cells | [89] | |
| PGDM | Hypermethylation | RETN | Decreased | Abdominal subcutaneous adipose tissue of adult offspring | [84] |
| Studies in Animals | |||||
| Experimental Model | Epigenetic Modification | Gene | Expression Effect | Tissue or Cell | Reference |
| Intrauterine growth retardation (IUGR) | Fetal stage: | Pdx1 | Rats with induced IUGR through the reduction of uteroplacental blood flow | [24] | |
| Deacetylation | |||||
| Histone H3 | Decreased | ||||
| Histone H4 | Decreased | ||||
| After birth: | |||||
| Demethylated H3K4 | N/S | ||||
| Methylated H3K9 | N/S | ||||
| High-fat diet (HFD) | Decreased H3K4me3 | Pfkl | Increased | Hepatoblasts and hepatocytes from mouse fetuses | [100] |
| Pdha1 | |||||
| Ogdh | |||||
| Acox1 | |||||
| Cpt1a | |||||
| Increased H3K9 binding Oxtr at transcriptional start site | Oxtr | Increased | Mouse hippocampus | [102] | |
| Hyperglycemia | H3K14ac | P300 | Increased | Mouse embryos | [103] |
| Deacetylation in H2K9 | Sirt1 | Decreased | |||
| GDM | Decreased H3K27me3 | Cartpt promoter | Increased | Mouse fetal ovaries | [82] |
| Increased H3K27ac | |||||
| H3K4me3 | Hspa1a | Increased | Offspring rat hearts | [101] | |
| PGDM | Decreased H3K4me3 | Pdx1 | Downregulated | Adult mouse pancreatic islets | [98] |
| Decreased H3K4me3 | Slc30a8 | Downregulated | |||
| Increased H3K4me3 | Aldh1a3 | Upregulated | |||
| Increased H3K4me3 | Ldha | Upregulated | |||
| Increased histone lysine malonylation in 21 histone lysine malonylation sites | N/S | N/S | Mouse brain cells | [99] | |
| Increased H3K9me3 | N/S | N/S | Neuroepithelium from mice embryos | [104] | |
| Multiparity-induced diabetes | Decreased H3K4me3 | Hhex | N/S | Mice pancreatic islets with FOXO1 deficient gene | [97] |
| Decreased H3K4me3 | Adcy5 | N/S | |||
| Decreased H3K4me3 | Lmo4 | Decreased | |||
| Increased H3K4me3 | Rapgef5 | Increased | |||
| Increased H3K4me3 | Ehf | Increased | |||
| Increased H3K4me3 | Gc | Increased | |||
| Studies in Humans | |||||
| Maternal Condition | Epigenetic Modification | Gene | Expression Effect | Tissue or Cell | Reference |
| GDM | H3K4me3 | NF-kB p65 | Upregulated | Maternal peripheral blood mononuclear cells and umbilical vein endothelial and cord blood mononuclear cells from newborns | [95] |
| H3K9ac | FOXO1 | Downregulated | Trophoblast cells | [96] | |
| H3K4me3 | No difference | ||||
| H3K27me3 | Several genes, see reference [97] | N/S | Cord blood and placenta from the fetal side | [105] | |
| N/S | HDAC2 | Downregulated | Maternal peripheral blood monocytes/macrophages | [109] | |
| Diabetes in pregnancy | Hypoacetylation H3K27 in males | N/S | N/S | Placental explants from the maternal side | [106] |
| PGDM | N/S | EHMT1 | Upregulated | Umbilical cord | [108] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meza-León, A.; Montoya-Estrada, A.; Reyes-Muñoz, E.; Romo-Yáñez, J. Diabetes Mellitus and Pregnancy: An Insight into the Effects on the Epigenome. Biomedicines 2024, 12, 351. https://doi.org/10.3390/biomedicines12020351
Meza-León A, Montoya-Estrada A, Reyes-Muñoz E, Romo-Yáñez J. Diabetes Mellitus and Pregnancy: An Insight into the Effects on the Epigenome. Biomedicines. 2024; 12(2):351. https://doi.org/10.3390/biomedicines12020351
Chicago/Turabian StyleMeza-León, Andrea, Araceli Montoya-Estrada, Enrique Reyes-Muñoz, and José Romo-Yáñez. 2024. "Diabetes Mellitus and Pregnancy: An Insight into the Effects on the Epigenome" Biomedicines 12, no. 2: 351. https://doi.org/10.3390/biomedicines12020351