
APOE Gene Variation’s Impact on Cardiovascular Health: A Case-Control Study




Abstract
1. Introduction
2. Materials and Methods
2.1. Vascular Material Collection and Listing
2.2. Primer Optimization, Gradient PCR and Gel Electrophoresis
2.3. RNA Isolation, cDNA Synthesis, and Gene Expression Analysis
2.4. Statistical Analysis
3. Results
3.1. Population Study Characteristics
3.2. Gradient PCR and Gel Electrophoresis Findings
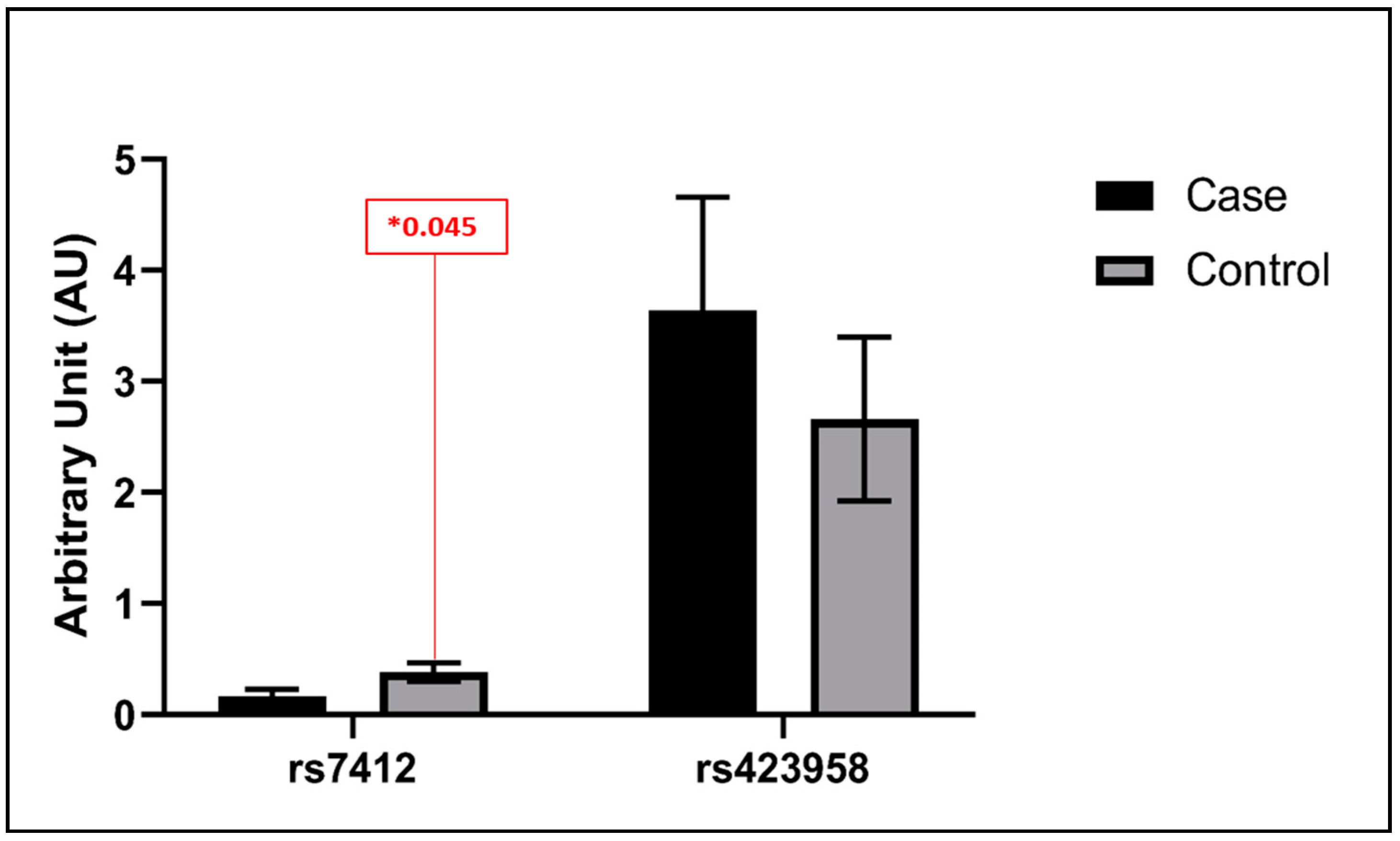
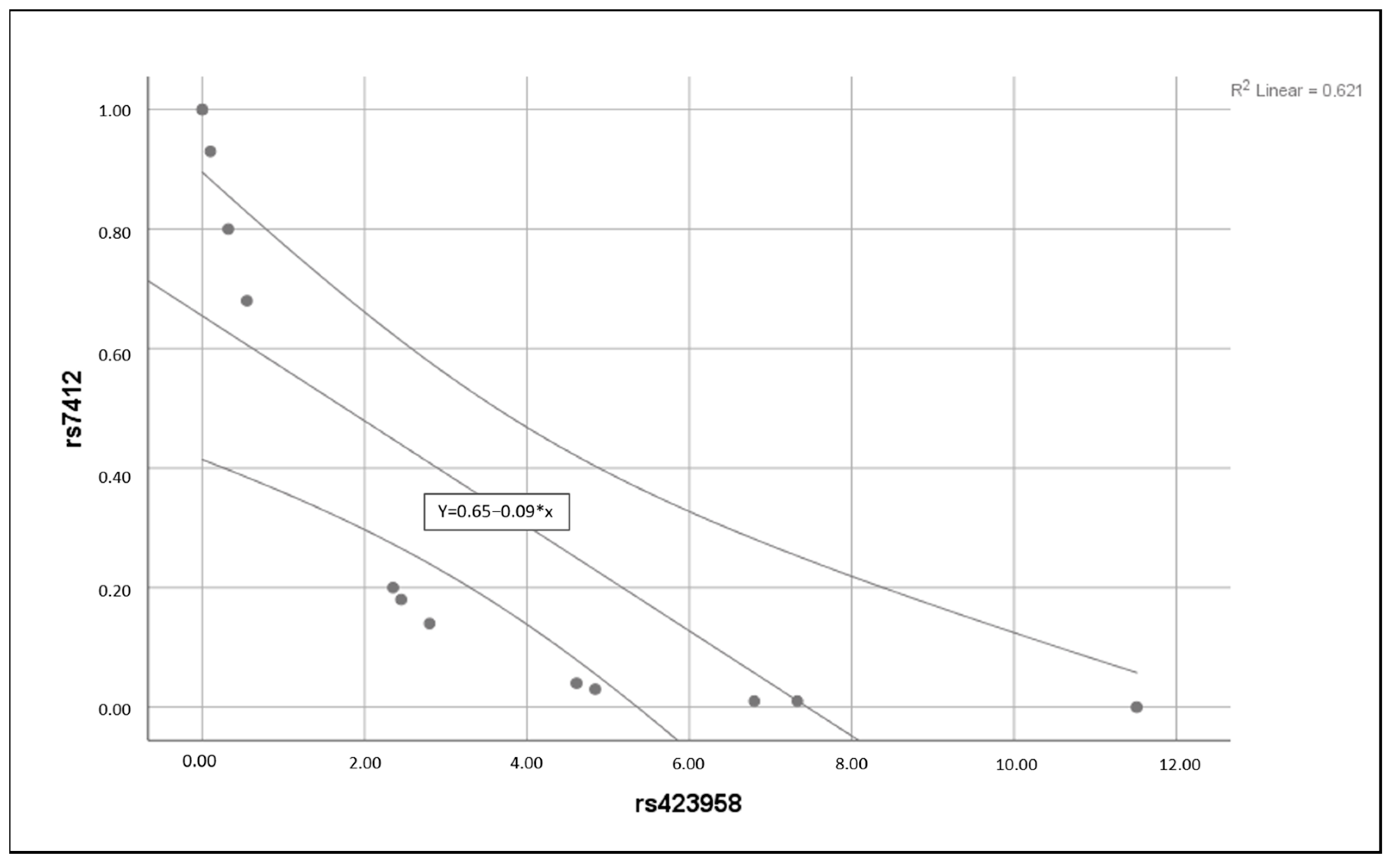
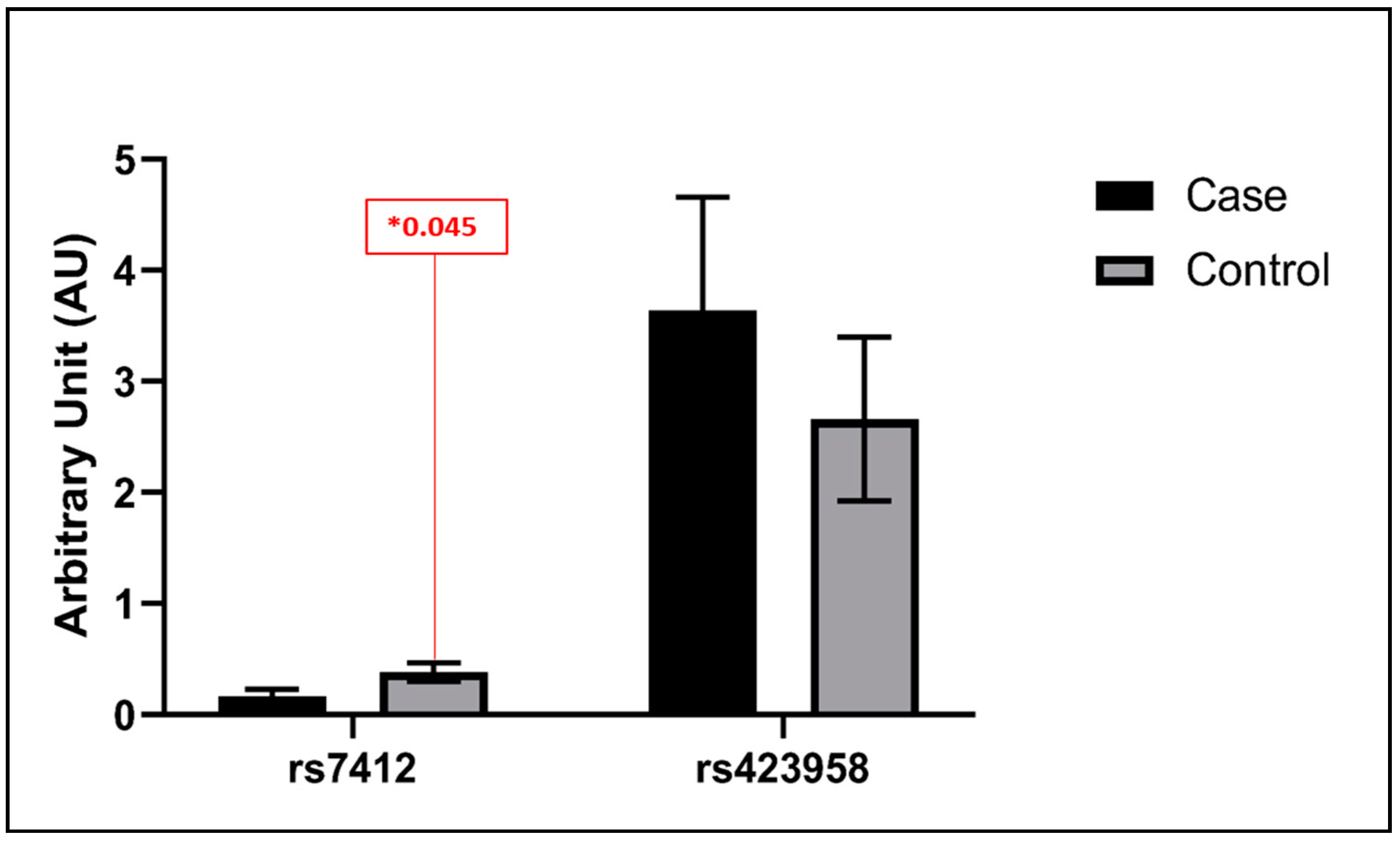
3.3. Statistical Analysis Findings
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CVI | Chronic venous Insufficiency |
| CVDs | Cardiovascular diseases |
| LEDVT | Lower extremity deep venous thrombosis |
| APOE | Apolipoprotein E |
| ε2 | Epsilon 2 |
| ε3 | Epsilon 3 |
| ε4 | Epsilon4 |
| SNPs | Single Nucleotide Polymorphisms |
| NCDs | Non-communicable diseases |
| IL-6 | Interleukin-6 |
| APOB | Apolipoprotein B |
| AD | Alzheimer’s disease |
| NEUH | Near East University Hospital |
| DNA | Deoxyribonucleic acid |
| PCR | Polymerase chain reaction |
| RTQ-PCR | Quantitative reverse transcription polymerase chain reaction |
| CT | Cycle Threshold |
| TM | Melting temperature |
References
- Bergan, J.J.; Schmid-Schönbein, G.W.; Smith, P.D.; Nicolaides, A.N.; Boisseau, M.R.; Eklof, B. Chronic venous disease. N. Engl. J. Med. 2006, 355, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Prochaska, J.H.; Arnold, N.; Falcke, A.; Kopp, S.; Schulz, A.; Buch, G.; Moll, S.; Panova-Noeva, M.; Jünger, C.; Eggebrecht, L. Chronic venous insufficiency, cardiovascular disease, and mortality: A population study. Eur. Heart J. 2021, 42, 4157–4165. [Google Scholar] [CrossRef] [PubMed]
- Beebe-Dimmer, J.L.; Pfeifer, J.R.; Engle, J.S.; Schottenfeld, D. The epidemiology of chronic venous insufficiency and varicose veins. Ann. Epidemiol. 2005, 15, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.J.; Fowkes, F.G.; Ruckley, C.V.; Lee, A.J. Prevalence of varicose veins and chronic venous insufficiency in men and women in the general population: Edinburgh Vein Study. J. Epidemiol. Community Health 1999, 53, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Vuylsteke, M.E.; Colman, R.; Thomis, S.; Guillaume, G.; Van Quickenborne, D.; Staelens, I. An Epidemiological Survey of Venous Disease Among General Practitioner Attendees in Different Geographical Regions on the Globe: The Final Results of the Vein Consult Program. Angiology 2018, 69, 779–785. [Google Scholar] [CrossRef]
- Curry, S.J.; Krist, A.H.; Owens, D.K.; Barry, M.J.; Caughey, A.B.; Davidson, K.W.; Doubeni, C.A.; Epling, J.W., Jr.; Kemper, A.R.; Kubik, M.; et al. Risk Assessment for Cardiovascular Disease With Nontraditional Risk Factors: US Preventive Services Task Force Recommendation Statement. JAMA 2018, 320, 272–280. [Google Scholar] [CrossRef]
- Chang, S.-L.; Huang, Y.-L.; Lee, M.-C.; Hu, S.; Hsiao, Y.-C.; Chang, S.-W.; Chang, C.J.; Chen, P.-C. Association of varicose veins with incident venous thromboembolism and peripheral artery disease. JAMA 2018, 319, 807–817. [Google Scholar] [CrossRef]
- Prandoni, P. Links between arterial and venous disease. J. Intern. Med. 2007, 262, 341–350. [Google Scholar] [CrossRef]
- Piazza, G. Varicose veins. Circulation 2014, 130, 582–587. [Google Scholar] [CrossRef]
- Ridker, P.M.; Rane, M. Interleukin-6 signaling and anti-interleukin-6 therapeutics in cardiovascular disease. Circ. Res. 2021, 128, 1728–1746. [Google Scholar] [CrossRef]
- Walsh, R.; Jurgens, S.J.; Erdmann, J.; Bezzina, C.R. Genome-wide association studies of cardiovascular disease. Physiol. Rev. 2023, 103, 2039–2055. [Google Scholar] [CrossRef] [PubMed]
- Bos, M.M.; de Vries, L.; Rensen, P.C.; van Dijk, K.W.; Blauw, G.J.; van Heemst, D.; Noordam, R. Apolipoprotein E genotype, lifestyle and coronary artery disease: Gene-environment interaction analyses in the UK Biobank population. Atherosclerosis 2021, 328, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wu, H.; Yu, Z.; Huang, Q.; Zhong, Z. APOE gene ε4 allele (388C-526C) effects on serum lipids and risk of coronary artery disease in southern Chinese Hakka population. J. Clin. Lab. Anal. 2021, 35, e23925. [Google Scholar] [CrossRef] [PubMed]
- Karczewski, K.J.; Francioli, L.C.; Tiao, G.; Cummings, B.B.; Alföldi, J.; Wang, Q.; Collins, R.L.; Laricchia, K.M.; Ganna, A.; Birnbaum, D.P. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature 2020, 581, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Painter, M.M.; Bu, G.; Kanekiyo, T. Apolipoprotein E as a Therapeutic Target in Alzheimer’s Disease: A Review of Basic Research and Clinical Evidence. CNS Drugs 2016, 30, 773–789. [Google Scholar] [CrossRef]
- Moghadasian, M.H.; McManus, B.M.; Nguyen, L.B.; Shefer, S.; Nadji, M.; Godin, D.V.; Green, T.J.; Hill, J.; Yang, Y.; Scudamore, C.H. Pathophysiology of apolipoprotein E deficiency in mice: Relevance to apo E-related disorders in humans. FASEB J. 2001, 15, 2623–2630. [Google Scholar] [CrossRef] [PubMed]
- Koopal, C.; Geerlings, M.I.; Muller, M.; de Borst, G.-J.; Algra, A.; van der Graaf, Y.; Visseren, F.L.; Group, S.S. The relation between apolipoprotein E (APOE) genotype and peripheral artery disease in patients at high risk for cardiovascular disease. Atherosclerosis 2016, 246, 187–192. [Google Scholar] [CrossRef]
- Riddell, D.R.; Graham, A.; Owen, J.S. Apolipoprotein E Inhibits Platelet Aggregation through the L-Arginine:Nitric Oxide Pathway: Implications for Vascular Disease. J. Biol. Chem. 1997, 272, 89–95. [Google Scholar] [CrossRef]
- Barbagallo, C.M.; Rizzo, M.; Noto, D.; Frasheri, A.; Pernice, V.; Rubino, A.; Pieri, D.; Pinto, V.; Cefalù, A.B.; Giordano, C.; et al. Accumulation of apoE-enriched triglyceride-rich lipoproteins in patients with coronary artery disease. Metabolism 2006, 55, 662–668. [Google Scholar] [CrossRef]
- Marais, A.D. Apolipoprotein E in lipoprotein metabolism, health and cardiovascular disease. Pathology 2019, 51, 165–176. [Google Scholar] [CrossRef]
- Majack, R.A.; Castle, C.K.; Goodman, L.V.; Weisgraber, K.H.; Mahley, R.W.; Shooter, E.M.; Gebicke-Haerter, P.J. Expression of apolipoprotein E by cultured vascular smooth muscle cells is controlled by growth state. J. Cell Biol. 1988, 107, 1207–1213. [Google Scholar] [CrossRef]
- Eberhardt, R.T.; Raffetto, J.D. Chronic venous insufficiency. Circulation 2014, 130, 333–346. [Google Scholar] [CrossRef]
- Brand, F.N.; Dannenberg, A.L.; Abbott, R.D.; Kannel, W.B. The epidemiology of varicose veins: The Framingham Study. Am. J. Prev. Med. 1988, 4, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Abondio, P.; Bruno, F.; Luiselli, D. Apolipoprotein E (APOE) Haplotypes in Healthy Subjects from Worldwide Macroareas: A Population Genetics Perspective for Cardiovascular Disease, Neurodegeneration, and Dementia. Curr. Issues Mol. Biol. 2023, 45, 2817–2831. [Google Scholar] [CrossRef]
- Modaghegh, M.H.S.; Saberianpour, S.; Amoueian, S.; Kamyar, M.M. Signaling pathways associated with structural changes in varicose veins: A case-control study. Phlebology 2022, 37, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Wang, Z.; Wu, X.; Shu, Y.; Lu, D. Apolipoprotein E polymorphism is associated with lower extremity deep venous thrombosis: Color-flow Doppler ultrasound evaluation. Lipids Health Dis. 2014, 13, 21. [Google Scholar] [CrossRef] [PubMed]
- Katrancioglu, N.; Manduz, S.; Ozen, F.; Yilmaz, M.B.; Atahan, E.; Ozdemir, O.; Berkan, O. Association between ApoE4 allele and deep venous thrombosis: A pilot study. Clin. Appl. Thromb. Hemost. 2011, 17, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Lahoz, C.; Schaefer, E.J.; Cupples, L.A.; Wilson, P.W.F.; Levy, D.; Osgood, D.; Parpos, S.; Pedro-Botet, J.; Daly, J.A.; Ordovas, J.M. Apolipoprotein E genotype and cardiovascular disease in the Framingham Heart Study. Atherosclerosis 2001, 154, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Raz, N.; Rodrigue, K.M.; Kennedy, K.M.; Land, S. Genetic and vascular modifiers of age-sensitive cognitive skills: Effects of COMT, BDNF, ApoE, and hypertension. Neuropsychology 2009, 23, 105–116. [Google Scholar] [CrossRef]
- Song, Y.; Stampfer, M.J.; Liu, S. Meta-analysis: Apolipoprotein E genotypes and risk for coronary heart disease. Ann. Intern. Med. 2004, 141, 137–147. [Google Scholar] [CrossRef]
- Larifla, L.; Armand, C.; Bangou, J.; Blanchet-Deverly, A.; Numeric, P.; Fonteau, C.; Michel, C.T.; Ferdinand, S.; Bourrhis, V.; Vélayoudom-Céphise, F.L. Association of APOE gene polymorphism with lipid profile and coronary artery disease in Afro-Caribbeans. PLoS ONE 2017, 12, e0181620. [Google Scholar] [CrossRef]
- Semaev, S.; Shakhtshneider, E.; Shcherbakova, L.; Ivanoshchuk, D.; Orlov, P.; Malyutina, S.; Gafarov, V.; Ragino, Y.; Voevoda, M. Associations of APOE Gene Variants rs429358 and rs7412 with Parameters of the Blood Lipid Profile and the Risk of Myocardial Infarction and Death in a White Population of Western Siberia. Curr. Issues Mol. Biol. 2022, 44, 1713–1724. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, K.L.; Tybjærg-Hansen, A.; Nordestgaard, B.G.; Frikke-Schmidt, R. Data on plasma levels of apolipoprotein E, correlations with lipids and lipoproteins stratified by APOE genotype, and risk of ischemic heart disease. Data Brief 2016, 6, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Frikke-Schmidt, R.; Tybjærg-Hansen, A.; Steffensen, R.; Jensen, G.; Nordestgaard, B.G. Apolipoprotein E genotype: Epsilon32 women are protected while epsilon43 and epsilon44 men are susceptible to ischemic heart disease: The Copenhagen City Heart Study. J. Am. Coll. Cardiol. 2000, 35, 1192–1199. [Google Scholar] [CrossRef] [PubMed]
- Kulminski, A.M.; Raghavachari, N.; Arbeev, K.G.; Culminskaya, I.; Arbeeva, L.; Wu, D.; Ukraintseva, S.V.; Christensen, K.; Yashin, A.I. Protective role of the apolipoprotein E2 allele in age-related disease traits and survival: Evidence from the Long Life Family Study. Biogerontology 2016, 17, 893–905. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Matsunaga, A.; Fukunaga, M.; Nagahama, K.; Hara, S.; Muso, E. Apolipoprotein E–related glomerular disorders. Kidney Int. 2020, 97, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Hu, M.; Sun, Y.; Lin, Y.; Zhu, C. Associations Between the Prevalence, Treatment, Control of Hypertension and Cognitive Trajectories Among Chinese Middle-Aged and Older Adults. Am. J. Geriatr. Psychiatry 2022, 30, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Misra, U.K.; Kalita, J.; Somarajan, B.I. Recurrent intracerebral hemorrhage in patients with hypertension is associated with APOE gene polymorphism: A preliminary study. J. Stroke Cerebrovasc. Dis. 2013, 22, 758–763. [Google Scholar] [CrossRef]
- Rao, H.; Wu, H.; Yu, Z.; Huang, Q. APOE Genetic Polymorphism rs7412 T/T Genotype May Be a Risk Factor for Essential Hypertension among Hakka People in Southern China. Int. J. Hypertens. 2022, 2022, 8145896. [Google Scholar] [CrossRef]
- Lan, X.; Wang, Z.; Zeng, Z.; Yao, H.; Xu, W.; Zhang, Y. Association of Different Combinations of ALDH2 rs671, APOE rs429358, rs7412 Polymorphisms with Hypertension in Middle-Aged and Elderly People: A Case-Control Study. Int. J. Gen. Med. 2023, 16, 915–927. [Google Scholar] [CrossRef]
- Liu, S.; Liu, J.; Weng, R.; Gu, X.; Zhong, Z. Apolipoprotein E gene polymorphism and the risk of cardiovascular disease and type 2 diabetes. BMC Cardiovasc. Disord. 2019, 19, 213. [Google Scholar] [CrossRef]
- Lumsden, A.L.; Mulugeta, A.; Zhou, A.; Hyppönen, E. Apolipoprotein E (APOE) genotype-associated disease risks: A phenome-wide, registry-based, case-control study utilising the UK Biobank. EBioMedicine 2020, 59, 102954. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.V.; Frikke-Schmidt, R.; Melis, D.; Luben, R.; Asselbergs, F.W.; Boer, J.M.; Cooper, J.; Palmen, J.; Horvat, P.; Engmann, J.; et al. A systematic review and meta-analysis of 130,000 individuals shows smoking does not modify the association of APOE genotype on risk of coronary heart disease. Atherosclerosis 2014, 237, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.D.; Gu, W.; Qiao, S.B.; Zhu, E.J.; Zhao, Q.M.; Lv, S.Z. Apolipoprotein E gene polymorphism and risk for coronary heart disease in the Chinese population: A meta-analysis of 61 studies including 6634 cases and 6393 controls. PLoS ONE 2014, 9, e95463. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tang, H.Q.; Peng, W.J.; Zhang, B.B.; Liu, M. Meta-analysis for the Association of Apolipoprotein E ε2/ε3/ε4 Polymorphism with Coronary Heart Disease. Chin. Med. J. 2015, 128, 1391–1398. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Group | Gender | Sample Size | Age |
|---|---|---|---|
| Patient | Total | n = 62 | Mean = 57 |
| Median = 49 | |||
| Mode = 41 | |||
| Female | n = 31 | Mean = 68 | |
| Median = 52 | |||
| Mode = 66 | |||
| Male | n = 31 | Mean = 46 | |
| Median = 43 | |||
| Mode = 55 | |||
| Control | Total | n = 53 | Mean = 59 |
| Median = 63 | |||
| Mode = 64 | |||
| Female | n = 18 | Mean = 62 | |
| Median = 68 | |||
| Mode = 77 | |||
| Male | n = 35 | Mean = 58 | |
| Median = 60 | |||
| Mode = 64 |
| rs429358 T>C-ε4 | Sequence | TM | Product Size |
|---|---|---|---|
| rs429358_Forward T | GCGGACATGGAGGACGTGT | 61–62 °C | 130 bp |
| rs429358_Reverse | GAGCCGCTTACGCAGCTTG | 61 °C | |
| rs7412 C>T-ε2 | |||
| rs7412_Forward C | GCCGATGACCTGCAGAAGC | 60–61 °C | 216 bp |
| rs7412_Reverse | GCTGCCCATCTCCTCCATC | 58–59 °C |
| Stage | Temperature | Time | Cycles |
|---|---|---|---|
| Initial denaturation | 95 °C | 2 min | 1 cycle |
| Denaturation | 95 °C | 30 s | |
| Annealing | 60–61 °C | 30 s | 40 cycles |
| Termination | 72 °C | 45 s | 1 cycle |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ismail, A.B.; Balcıoğlu, Ö.; Özcem, B.; Ergoren, M.Ç. APOE Gene Variation’s Impact on Cardiovascular Health: A Case-Control Study. Biomedicines 2024, 12, 695. https://doi.org/10.3390/biomedicines12030695
Ismail AB, Balcıoğlu Ö, Özcem B, Ergoren MÇ. APOE Gene Variation’s Impact on Cardiovascular Health: A Case-Control Study. Biomedicines. 2024; 12(3):695. https://doi.org/10.3390/biomedicines12030695
Chicago/Turabian StyleIsmail, Aya Badeea, Özlem Balcıoğlu, Barçın Özcem, and Mahmut Çerkez Ergoren. 2024. "APOE Gene Variation’s Impact on Cardiovascular Health: A Case-Control Study" Biomedicines 12, no. 3: 695. https://doi.org/10.3390/biomedicines12030695
APA StyleIsmail, A. B., Balcıoğlu, Ö., Özcem, B., & Ergoren, M. Ç. (2024). APOE Gene Variation’s Impact on Cardiovascular Health: A Case-Control Study. Biomedicines, 12(3), 695. https://doi.org/10.3390/biomedicines12030695