
L-PRF Secretome from Both Smokers/Nonsmokers Stimulates Angiogenesis and Osteoblast Differentiation In Vitro
 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. L-PRF Sampling and L-PRF Secretome Recovery
2.2. Enzyme-Linked Immunosorbent Assay (ELISA)
2.3. Tube Formation In Vitro
2.4. Western Blotting
2.5. Osteogenic Differentiation
2.6. Alizarin Red Stain
2.7. Immunostaining
2.8. Statistical Analyses
3. Results
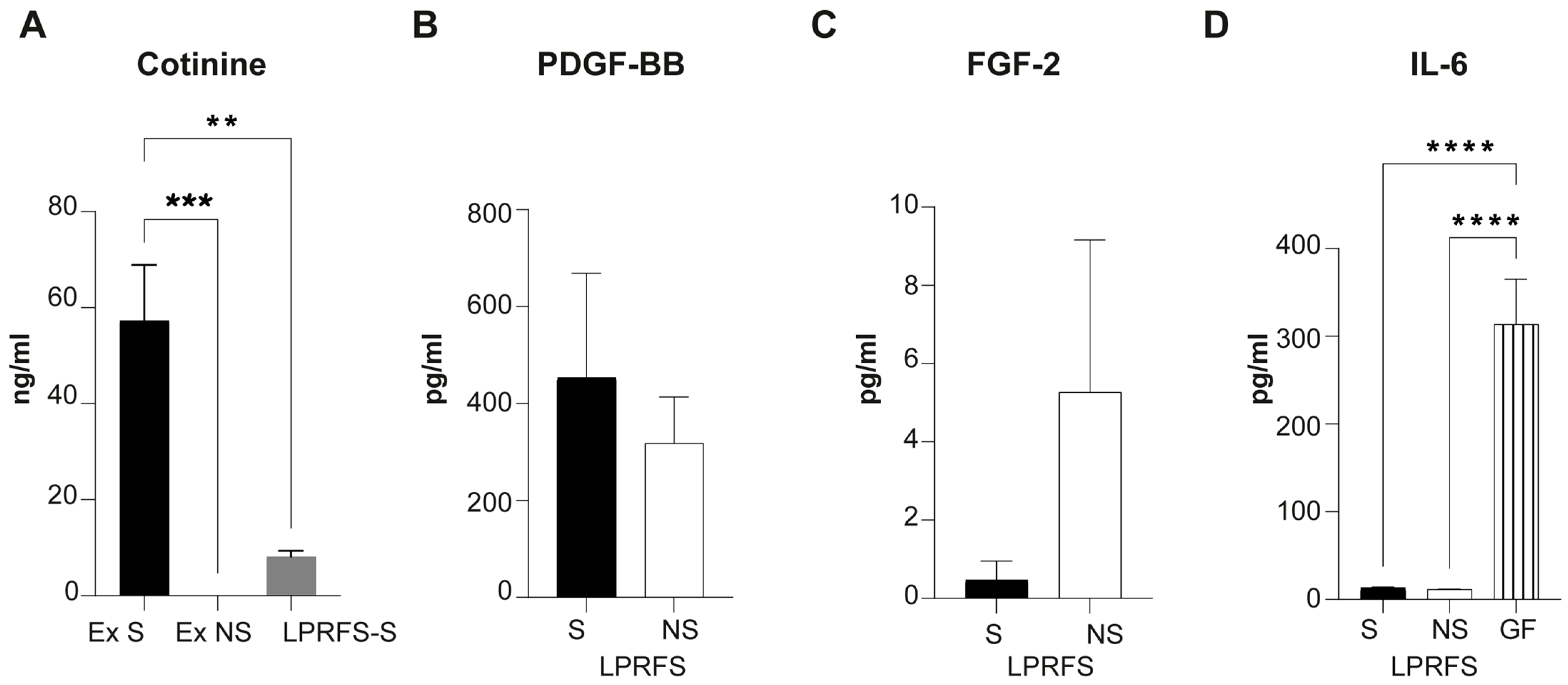
3.1. LPRFS from Smokers and Nonsmokers Releases Similar Levels of PDGF-BB and FGF-b Growth Factors
3.2. LPRFS Derived from Smokers and Nonsmokers Promotes Angiogenesis In Vitro
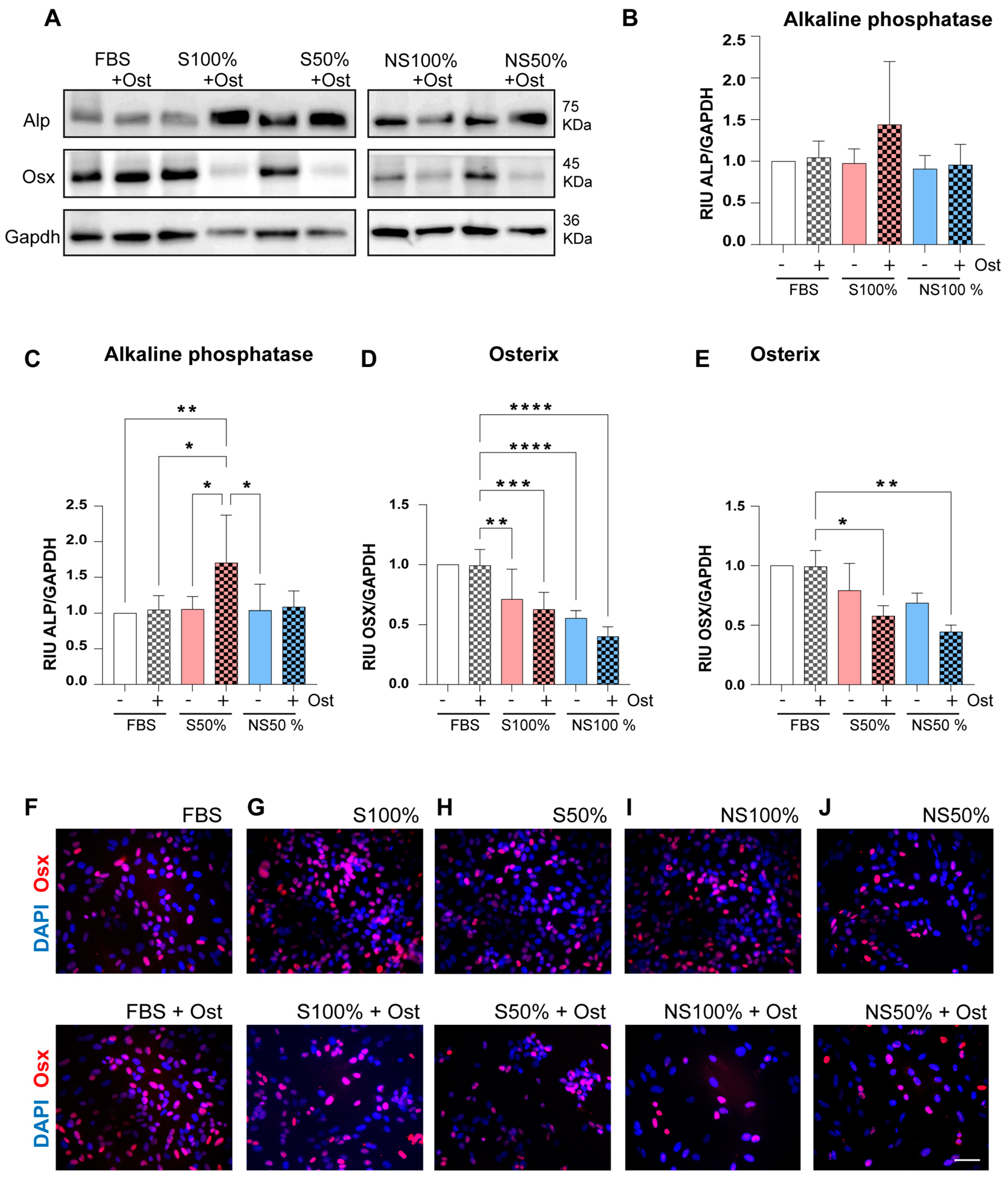
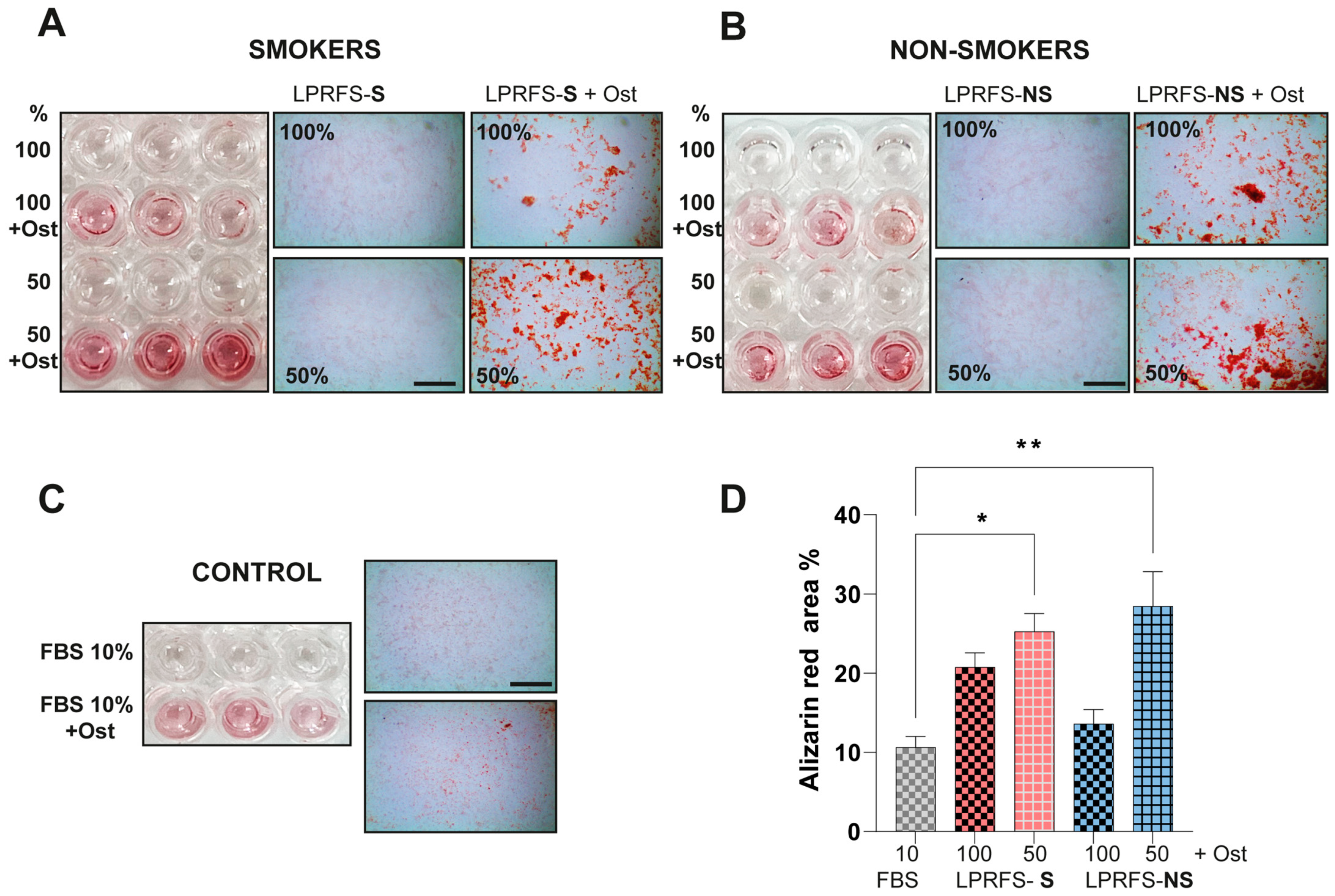
3.3. LPRFS Derived from Smokers/Nonsmokers Increases Bone Differentiation In Vitro
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fan, Y.; Perez, K.; Dym, H. Clinical Uses of Platelet-Rich Fibrin in Oral and Maxillofacial Surgery. Dent. Clin. N. Am. 2020, 64, 291–303. [Google Scholar] [CrossRef]
- Miron, R.J.; Fujioka-Kobayashi, M.; Sculean, A.; Zhang, Y. Optimization of Platelet-rich Fibrin. Periodontology 2000 2023, 1–13. [Google Scholar] [CrossRef]
- Quirynen, M.; Siawasch, S.A.M.; Temmerman, A.; Cortellini, S.; Dhondt, R.; Teughels, W.; Castro, A.B. Do Autologous Platelet Concentrates (APCs) Have a Role in Intra-oral Bone Regeneration? A Critical Review of Clinical Guidelines on Decision-making Process. Periodontology 2000 2023, 93, 254–269. [Google Scholar] [CrossRef]
- Avila-Ortiz, G.; Ambruster, J.; Barootchi, S.; Chambrone, L.; Chen, C.; Dixon, D.R.; Geisinger, M.L.; Giannobile, W.V.; Goss, K.; Gunsolley, J.C.; et al. American Academy of Periodontology Best Evidence Consensus Statement on the Use of Biologics in Clinical Practice. J. Periodontol. 2022, 93, 1763–1770. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.B.; Meschi, N.; Temmerman, A.; Pinto, N.; Lambrechts, P.; Teughels, W.; Quirynen, M. Regenerative Potential of Leucocyte- and Platelet-Rich Fibrin. Part B: Sinus Floor Elevation, Alveolar Ridge Preservation and Implant Therapy. A Systematic Review. J. Clin. Periodontol. 2017, 44, 225–234. [Google Scholar] [CrossRef]
- Pinto, N.R.; Ubilla, M.; Zamora, Y.; Del Rio, V.; Ehrenfest, D.M.D.; Quirynen, M. Leucocyte- and Platelet-Rich Fibrin (L-PRF) as a Regenerative Medicine Strategy for the Treatment of Refractory Leg Ulcers: A Prospective Cohort Study. Platelets 2018, 29, 468–475. [Google Scholar] [CrossRef]
- de Lima Barbosa, R.; Stellet Lourenço, E.; de Azevedo dos Santos, J.V.; Rodrigues Santiago Rocha, N.; Mourão, C.F.; Alves, G.G. The Effects of Platelet-Rich Fibrin in the Behavior of Mineralizing Cells Related to Bone Tissue Regeneration—A Scoping Review of In Vitro Evidence. J. Funct. Biomater. 2023, 14, 503. [Google Scholar] [CrossRef]
- Strauss, F.J.; Nasirzade, J.; Kargarpoor, Z.; Stahli, A.; Gruber, R. Effect of Platelet-Rich Fibrin on Cell Proliferation, Migration, Differentiation, Inflammation, and Osteoclastogenesis: A Systematic Review of in Vitro Studies. Clin. Oral Investig. 2020, 24, 569–584. [Google Scholar] [CrossRef] [PubMed]
- Kargarpour, Z.; Nasirzade, J.; Strauss, F.J.; Di Summa, F.; Hasannia, S.; Müller, H.; Gruber, R. Platelet-rich Fibrin Suppresses in Vitro Osteoclastogenesis. J. Periodontol. 2020, 91, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Ferreira Sávio, D.d.S.; da Silva, L.M.P.; Reis, G.G.D.; Denardi, R.J.; da Costa, N.M.M.; Chaves Furlaneto, F.A.; de Souza, S.L.S.; Mourão, C.F.d.A.B.; Miron, R.J.; Okamoto, R.; et al. Effects of Platelet-Rich Fibrin Produced by Three Centrifugation Protocols on Bone Neoformation in Defects Created in Rat Calvaria. Platelets 2023, 34, 2228417. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, L.T. Wound Healing and Infection in Surgery: The Pathophysiological Impact of Smoking, Smoking Cessation, and Nicotine Replacement Therapy: A Systematic Review. Ann. Surg. 2012, 255, 1069–1079. [Google Scholar] [CrossRef]
- Tatsumi, M.; Yanagita, M.; Yamashita, M.; Hasegawa, S.; Ikegami, K.; Kitamura, M.; Murakami, S. Long-term Exposure to Cigarette Smoke Influences Characteristics in Human Gingival Fibroblasts. J. Periodontal. Res. 2021, 56, 951–963. [Google Scholar] [CrossRef]
- Apatzidou, D.A. The Role of Cigarette Smoking in Periodontal Disease and Treatment Outcomes of Dental Implant Therapy. Periodontol. 2000 2022, 90, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.A.; Wilson, R.F.; Palmer, R.M. The Effect of Smoking on Periodontal Bone Regeneration: A Systematic Review and Meta-Analysis. J. Periodontol. 2012, 83, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Ríos, S.; Álvarez, S.; Smith, P.C.; Sáez, C.G.; Andrade, C.; Pinto, N.; Martínez, C.E. Smoking Habits Do Not Affect Biological Responses Induced by Leucocyte and Platelet-Rich Fibrin in Periodontal Ligament Cells. J. Oral Pathol. Med. 2023, 52, 169–180. [Google Scholar] [CrossRef]
- Lara, C.; Bezmalinovic, A.; García-Herrera, C.; Ríos, S.; Valenzuela, L.M.; Martínez, C.E. Leukocyte- and Platelet-Rich Fibrin (L-PRF) Obtained from Smokers and Nonsmokers Shows a Similar Uniaxial Tensile Response In Vitro. Biomedicines 2023, 11, 3286. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.B.; Andrade, C.; Li, X.; Pinto, N.; Teughels, W.; Quirynen, M. Impact of g Force and Timing on the Characteristics of Platelet-Rich Fibrin Matrices. Sci. Rep. 2021, 11, 6038. [Google Scholar] [CrossRef] [PubMed]
- Arnaoutova, I.; Kleinman, H.K. In Vitro Angiogenesis: Endothelial Cell Tube Formation on Gelled Basement Membrane Extract. Nat. Protoc. 2010, 5, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Strzelecka-Kiliszek, A.; Romiszewska, M.; Bozycki, L.; Mebarek, S.; Bandorowicz-Pikula, J.; Buchet, R.; Pikula, S. Src and ROCK Kinases Differentially Regulate Mineralization of Human Osteosarcoma Saos-2 Cells. Int. J. Mol. Sci. 2019, 20, 2872. [Google Scholar] [CrossRef]
- Talhout, R.; Schulz, T.; Florek, E.; Van Benthem, J.; Wester, P.; Opperhuizen, A. Hazardous Compounds in Tobacco Smoke. Int. J. Environ. Res. Public Health 2011, 8, 613–628. [Google Scholar] [CrossRef]
- Qin, Y.; Liu, Y.; Jiang, Y.; Mei, S.; Liu, Y.; Feng, J.; Guo, L.; Du, J.; Graves, D.T.; Liu, Y. Cigarette Smoke Exposure Inhibits Osteoclast Apoptosis via the MtROS Pathway. J. Dent. Res. 2021, 100, 1378–1386. [Google Scholar] [CrossRef]
- Tarantino, U.; Cariati, I.; Greggi, C.; Gasbarra, E.; Belluati, A.; Ciolli, L.; Maccauro, G.; Momoli, A.; Ripanti, S.; Falez, F.; et al. Skeletal System Biology and Smoke Damage: From Basic Science to Medical Clinic. Int. J. Mol. Sci. 2021, 22, 6629. [Google Scholar] [CrossRef] [PubMed]
- Garip, M.; Van Dessel, J.; Grosjean, L.; Politis, C.; Bila, M. The Impact of Smoking on Surgical Complications after Head and Neck Reconstructive Surgery with a Free Vascularised Tissue Flap: A Systematic Review and Meta-Analysis. Br. J. Oral Maxillofac. Surg. 2021, 59, e79–e98. [Google Scholar] [CrossRef] [PubMed]
- Ye, D.; Gajendra, S.; Lawyer, G.; Jadeja, N.; Pishey, D.; Pathagunti, S.; Lyons, J.; Veazie, P.; Watson, G.; McIntosh, S.; et al. Inflammatory Biomarkers and Growth Factors in Saliva and Gingival Crevicular Fluid of E-Cigarette Users, Cigarette Smokers, and Dual Smokers: A Pilot Study. J. Periodontol. 2020, 91, 1274–1283. [Google Scholar] [CrossRef]
- Smith, P.C.; Martinez, C.; Caceres, M.; Martinez, J. Research on Growth Factors in Periodontology. Periodontology 2000 2015, 67, 234–250. [Google Scholar] [CrossRef] [PubMed]
- Qasim, M.; Chae, D.S.; Lee, N.Y. Bioengineering Strategies for Bone and Cartilage Tissue Regeneration Using Growth Factors and Stem Cells. J. Biomed. Mater. Res. A 2020, 108, 394–411. [Google Scholar] [CrossRef]
- Aldaham, S.; Foote, J.A.; Chow, H.-H.S.; Hakim, I.A. Smoking Status Effect on Inflammatory Markers in a Randomized Trial of Current and Former Heavy Smokers. Int. J. Inflam. 2015, 2015, 439396. [Google Scholar] [CrossRef] [PubMed]
- Blatt, S.; Burkhardt, V.; Kämmerer, P.W.; Pabst, A.M.; Sagheb, K.; Heller, M.; Al-Nawas, B.; Schiegnitz, E. Biofunctionalization of Porcine-Derived Collagen Matrices with Platelet Rich Fibrin: Influence on Angiogenesis in Vitro and in Vivo. Clin. Oral Investig. 2020, 24, 3425–3436. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, J.; Vangansewinkel, T.; Gervois, P.; Merckx, G.; Hilkens, P.; Quirynen, M.; Lambrichts, I.; Bronckaers, A. Angiogenic Properties of ‘Leukocyte- and Platelet-Rich Fibrin’. Sci. Rep. 2018, 8, 14632. [Google Scholar] [CrossRef]
- You, J.-S.; Kim, S.-G.; Oh, J.-S.; Kim, J.-S. Effects of Platelet-Derived Material (Platelet-Rich Fibrin) on Bone Regeneration. Implant. Dent. 2019, 28, 244–255. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Li, M.; Wang, S.; Xiao, Z.; Xiong, Y.; Wang, G. Recent Advances of Osterix Transcription Factor in Osteoblast Differentiation and Bone Formation. Front. Cell Dev. Biol. 2020, 8, 601224. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, H.; Zhang, Z.; Yan, Z.; Lv, H.; Zhang, Y.; Wu, B. Platelet-rich Fibrin Exudate Promotes the Proliferation and Osteogenic Differentiation of Human Periodontal Ligament Cells In vitro. Mol. Med. Rep. 2018, 18, 4477–4485. [Google Scholar] [CrossRef] [PubMed]
- Hoshikawa, S.; Shimizu, K.; Watahiki, A.; Chiba, M.; Saito, K.; Wei, W.; Fukumoto, S.; Inuzuka, H. Phosphorylation-dependent Osterix Degradation Negatively Regulates Osteoblast Differentiation. FASEB J. 2020, 34, 14930–14945. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ríos, S.; González, L.G.; Saez, C.G.; Smith, P.C.; Escobar, L.M.; Martínez, C.E. L-PRF Secretome from Both Smokers/Nonsmokers Stimulates Angiogenesis and Osteoblast Differentiation In Vitro. Biomedicines 2024, 12, 874. https://doi.org/10.3390/biomedicines12040874
Ríos S, González LG, Saez CG, Smith PC, Escobar LM, Martínez CE. L-PRF Secretome from Both Smokers/Nonsmokers Stimulates Angiogenesis and Osteoblast Differentiation In Vitro. Biomedicines. 2024; 12(4):874. https://doi.org/10.3390/biomedicines12040874
Chicago/Turabian StyleRíos, Susana, Lina Gabriela González, Claudia Gilda Saez, Patricio Cristian Smith, Lina M. Escobar, and Constanza Eugenia Martínez. 2024. "L-PRF Secretome from Both Smokers/Nonsmokers Stimulates Angiogenesis and Osteoblast Differentiation In Vitro" Biomedicines 12, no. 4: 874. https://doi.org/10.3390/biomedicines12040874