
MicroRNA-195-5p Attenuates Intracerebral-Hemorrhage-Induced Brain Damage by Inhibiting MMP-9/MMP-2 Expression

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Preparation and Surgical Preparation: Induction of Intracerebral Hemorrhage
2.2. Neurological Tests
2.2.1. Corner Turn Test
2.2.2. Forelimb Grip Strength Test
2.3. Measurement of Brain Water Content
2.4. Evaluation of Blood–Brain Barrier (BBB) Permeability
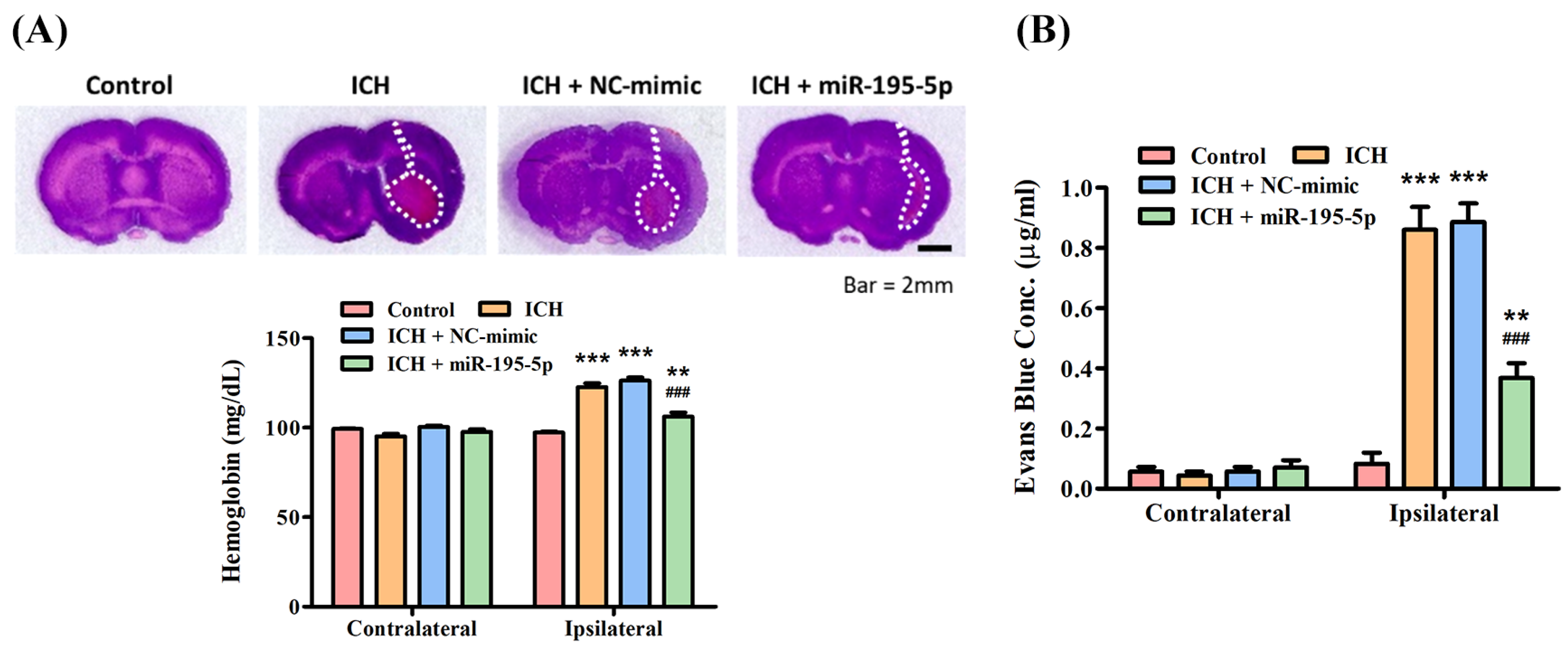
2.5. Evaluation of Hematoma Value (Spectrophotometric Assay)
2.6. Tissue Preparation for TUNEL Stain
2.7. Western Blotting Analysis
2.8. Matrix Metalloproteinase Gelatin Zymography
2.9. Statistical Analysis
3. Results
3.1. Neurological Tests
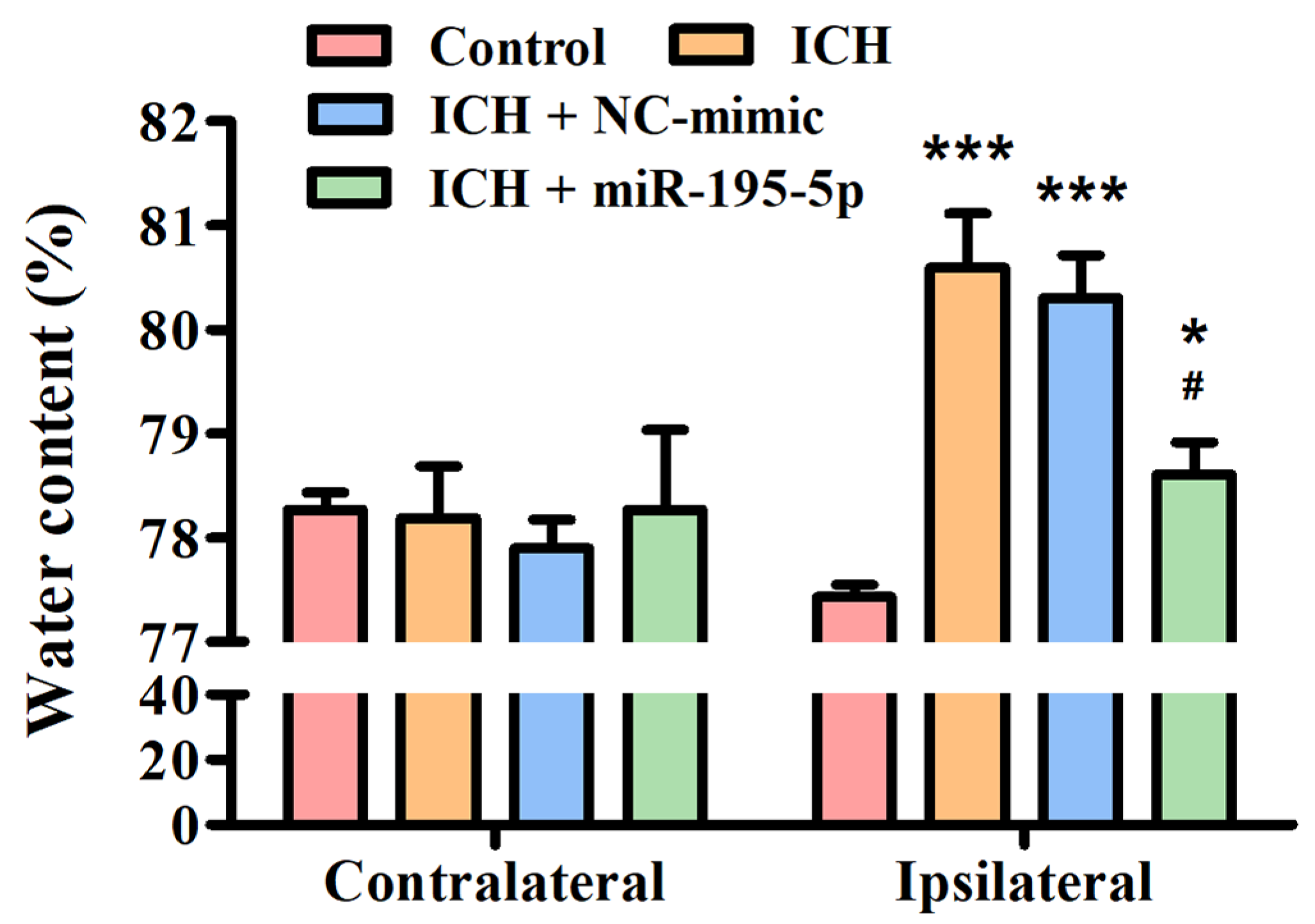
3.2. Brain Water Content
3.3. miR-195-5p Improves the Blood–Brain Barrier Permeability after ICH
3.4. miR-195-5p Inhibits Neuron Apoptosis after ICH
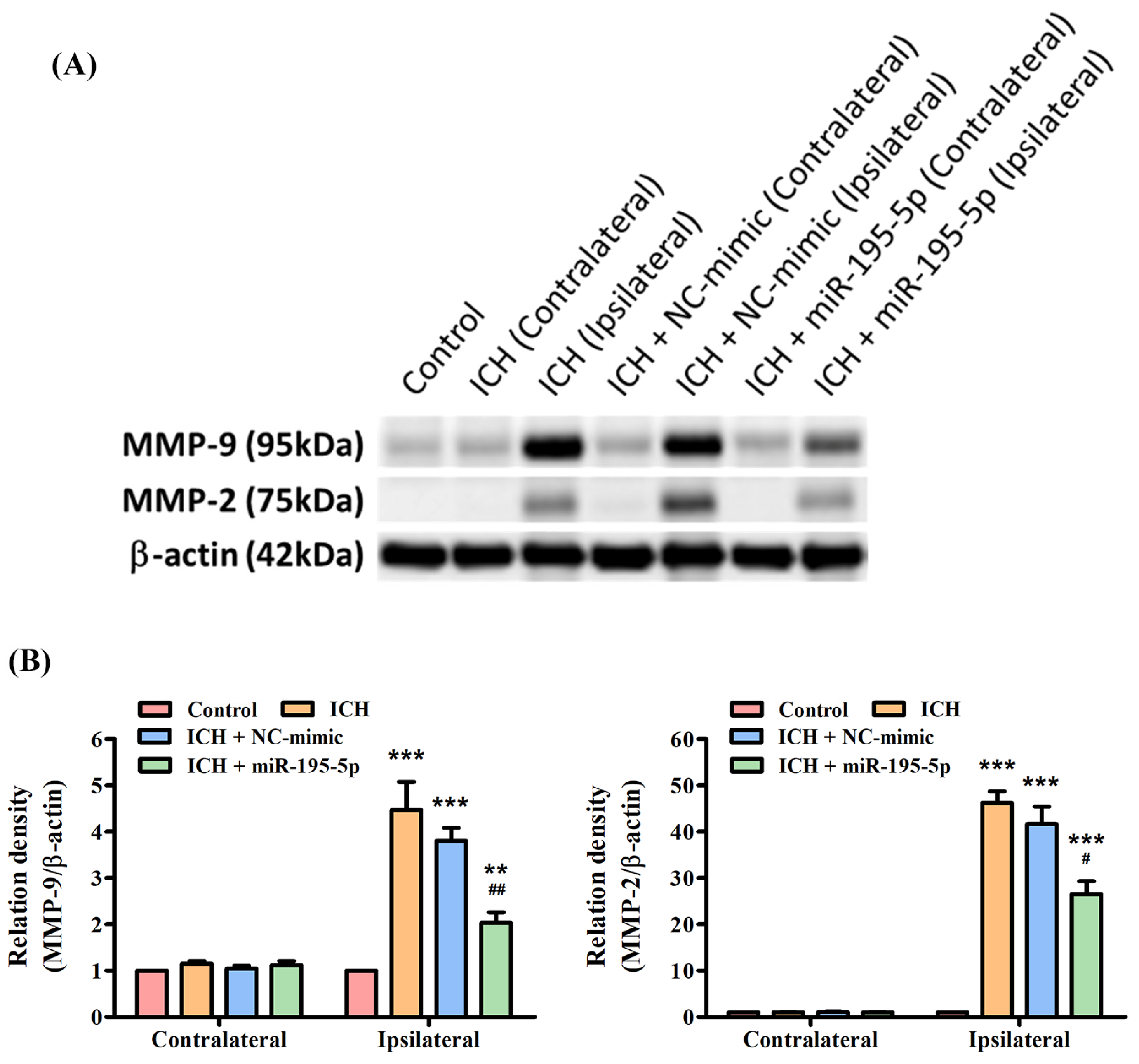
3.5. miR-195-5p Inhibits MMP-9/MMP-2 Enzymatic Activity after ICH
3.6. miR-195-5p Reduces the Protein Expression of ICH-Induced MMP-9 and MMP-2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Broderick, J.P.; Brott, T.G.; Duldner, J.E.; Tomsick, T.; Huster, G. Volume of intracerebral hemorrhage. A powerful and easy-to-use predictor of 30-day mortality. Stroke 1993, 24, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Wagner, K.R.; Xi, G.; Hua, Y.; Kleinholz, M.; de Courten-Myers, G.M.; Myers, R.E. Early metabolic alterations in edematous peri-hematomal brain regions following experimental intracerebral hemorrhage. J. Neurosurg. 1998, 88, 1058–1065. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, G.A. Matrix metalloproteinases and their multiple roles in neurodegenerative diseases. Lancet Neurol. 2009, 8, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Estrada, E.Y.; Thompson, J.F.; Liu, W.; Rosenberg, G.A. Matrix metalloproteinase-mediated disruption of tight junction proteins in cerebral vessels is reversed by synthetic matrix metalloproteinase inhibitor in focal ischemia in rat. J. Cereb. Blood Flow Metab. 2007, 27, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Tsirka, S.E. Neuroprotection by inhibition of matrix metalloproteinases in a mouse model of intracerebral haemor-rhage. Brain 2005, 128, 1622–1633. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Chen, X.; Wang, Z.; Zhang, M.; Meng, H.; Gao, Y.; Luo, B.; Tao, L.; Chen, Y. Poloxamer-188 can attenuate blood-brain barrier damage to exert neuroprotective effect in mice intracerebral hemorrhage model. J. Mol. Neurosci. 2015, 55, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Li, S.; Xiong, Q.; Zhou, L.; Luo, A. Effect of Propofol on the Expression of MMP-9 and Its Relevant Inflammatory Factors in Brain of Rat with Intracerebral Hemorrhage. Cell Biochem. Biophys. 2015, 72, 675–679. [Google Scholar] [CrossRef] [PubMed]
- Lagos-Quintana, M.; Rauhut, R.; Yalcin, A.; Meyer, J.; Lendeckel, W.; Tuschl, T. Identification of tissue-specific microRNAs from mouse. Curr. Biol. 2002, 12, 735–739. [Google Scholar] [CrossRef] [PubMed]
- Costinean, S.; Zanesi, N.; Pekarsky, Y.; Tili, E.; Volinia, S.; Heerema, N.; Croce, C.M. Pre-B cell proliferation and lymphoblastic leukemia/high-grade lymphoma in E(mu)-miR155 transgenic mice. Proc. Natl. Acad. Sci. USA 2006, 103, 7024–7029. [Google Scholar] [CrossRef]
- Kim, V.N. MicroRNA biogenesis: Coordinated cropping and dicing. Nat. Rev. Mol. Cell Biol. 2005, 6, 376–385. [Google Scholar] [CrossRef]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.W.; Mendell, J.T. MicroRNAs in cell proliferation, cell death, and tumorigenesis. Br. J. Cancer 2006, 94, 776–780. [Google Scholar] [CrossRef] [PubMed]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—microRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.C. OncomiRs: The discovery and progress of microRNAs in cancers. Mol. Cancer 2007, 6, 60. [Google Scholar] [CrossRef] [PubMed]
- Suárez, Y.; Fernández-Hernando, C.; Yu, J.; Gerber, S.A.; Harrison, K.D.; Pober, J.S.; Iruela-Arispe, M.L.; Merkenschlager, M.; Sessa, W.C. Dicer-dependent endothelial microRNAs are necessary for postnatal angiogenesis. Proc. Natl. Acad. Sci. USA 2008, 105, 14082–14087. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-S.; Wang, H.-Y.J.; Liao, Y.-C.; Tsai, P.-C.; Chen, K.-C.; Cheng, H.-Y.; Lin, R.-T.; Juo, S.-H.H. MicroRNA-195 regulates vascular smooth muscle cell phenotype and prevents neointimal formation. Cardiovasc. Res. 2012, 95, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Poliseno, L.; Tuccoli, A.; Mariani, L.; Evangelista, M.; Citti, L.; Woods, K.; Mercatanti, A.; Hammond, S.; Rainaldi, G. MicroRNAs modulate the angiogenic properties of HUVECs. Blood 2006, 108, 3068–3071. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.; Cheng, Y.; Yue, J.; Yang, J.; Liu, X.; Chen, H.; Dean, D.B.; Zhang, C. MicroRNA expression signature and antisense-mediated depletion reveal an essential role of MicroRNA in vascular neointimal lesion formation. Circ. Res. 2007, 100, 1579–1588. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.-Z.; Tian, Y.; Ander, B.P.; Xu, H.; Stamova, B.S.; Zhan, X.; Turner, R.J.; Jickling, G.; Sharp, F.R. Brain and blood microRNA expression profiling of ischemic stroke, intracerebral hemorrhage, and kainate seizures. J. Cereb. Blood Flow Metab. 2010, 30, 92–101. [Google Scholar] [CrossRef]
- Chen, K.C.; Wang, Y.S.; Hu, C.Y.; Chang, W.C.; Liao, Y.C.; Dai, C.Y.; Juo, S.H. OxLDL up-regulates microRNA-29b, leading to epigenetic modifications of MMP-2/MMP-9 genes: A novel mechanism for cardiovascular diseases. FASEB J. 2011, 25, 1718–1728. [Google Scholar] [CrossRef]
- Tan, J.R.; Koo, Y.X.; Kaur, P.; Liu, F.; Armugam, A.; Wong, P.T.; Jeyaseelan, K. microRNAs in stroke pathogenesis. Curr. Mol. Med. 2011, 11, 76–92. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Juo, S.H. MicroRNAs in atherosclerosis. Kaohsiung J. Med. Sci. 2012, 28, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Wang, Y.; Lin, J.; Li, N.; Zhao, X.; Liu, G.; Liu, L.; Jiao, Y.; Gu, W.; Wang, D.Z.; et al. Circulating MicroRNAs as potential risk biomarkers for hematoma enlargement after intracerebral hemorrhage. CNS Neurosci. Ther. 2012, 18, 1003–1011. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.; Liu, J.; Wang, W.; Hao, F.; Sun, X.; Wu, X.; Bu, P.; Zhang, Y.; Liu, Y.; Liu, F.; et al. Alteration in abundance and compartmentalization of inflammation-related miRNAs in plasma after intracerebral hemorrhage. Stroke 2013, 44, 1739–1742. [Google Scholar] [CrossRef] [PubMed]
- Wasserman, J.K.; Zhu, X.; Schlichter, L.C. Evolution of the inflammatory response in the brain following intracerebral hemorrhage and effects of delayed minocycline treatment. Brain Res. 2007, 1180, 140–154. [Google Scholar] [CrossRef] [PubMed]
- Lu, A.; Tang, Y.; Ran, R.; Ardizzone, T.L.; Wagner, K.R.; Sharp, F.R. Brain genomics of intracerebral hemorrhage. J. Cereb. Blood Flow Metab. 2006, 26, 230–252. [Google Scholar] [CrossRef] [PubMed]
- Rosell, A.; Vilalta, A.; Garcia-Berrocoso, T.; Fernandez-Cadenas, I.; Domingues-Montanari, S.; Cuadrado, E.; Delgado, P.; Ribó, M.; Martínez-Sáez, E.; Ortega-Aznar, A.; et al. Brain perihema-toma genomic profile following spontaneous human intracerebral hemorrhage. PLoS ONE 2011, 6, e16750. [Google Scholar] [CrossRef] [PubMed]
- Bu, L.; Tian, Y.; Wen, H.; Jia, W.; Yang, S. miR-195-5p exerts tumor-suppressive functions in human lung cancer cells through targeting TrxR2. Acta Biochim. Biophys. Sin. 2021, 53, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Li, Q.; Zhang, L.; Zhong, L.; Gu, M.; He, B.; Qu, Q.; Lao, Y.; Gu, K.; Zheng, B.; et al. Serum miR-195-5p Exhibits Clinical Sig-nificance in the Diagnosis of Essential Hypertension with Type 2 Diabetes Mellitus by Targeting DRD1. Clinics 2021, 76, e2502. [Google Scholar] [CrossRef]
- Xu, P.; Zhao, Y.; Liu, M.; Wang, Y.; Wang, H.; Li, Y.X.; Zhu, X.; Yao, Y.; Wang, H.; Qiao, J.; et al. Variations of microRNAs in human placentas and plasma from preeclamptic pregnancy. Hypertension 2014, 63, 1276–1284. [Google Scholar] [CrossRef]
- Xu, Y.; Jiang, W.; Zhong, L.; Li, H.; Bai, L.; Chen, X.; Lin, Y.; Zheng, D. miR-195-5p alleviates acute kidney injury through repression of inflammation and oxidative stress by targeting vascular endothelial growth factor A. Aging 2020, 12, 10235–10245. [Google Scholar] [CrossRef]
- Jeong, S.-W.; Chu, K.; Jung, K.-H.; Kim, S.U.; Kim, M.; Roh, J.-K. Human neural stem cell transplantation promotes functional recovery in rats with experimental intracerebral hemorrhage. Stroke 2003, 34, 2258–2263. [Google Scholar] [CrossRef]
- Jung, K.-H.; Chu, K.; Jeong, S.-W.; Han, S.-Y.; Lee, S.-T.; Kim, J.-Y.; Kim, M.; Roh, J.-K. HMG-CoA reductase inhibitor, atorvastatin, promotes sensorimotor recovery, suppressing acute inflammatory reaction after experimental intracerebral hemorrhage. Stroke 2004, 35, 1744–1749. [Google Scholar] [CrossRef]
- Asahi, M.; Asahi, K.; Wang, X.; Lo, E.H. Reduction of tissue plasminogen activator-induced hemorrhage and brain injury by free radical spin trapping after embolic focal cerebral ischemia in rats. J. Cereb. Blood Flow Metab. 2000, 20, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Xia, M.; Guo, C.; Jia, Z.; Wang, J.; Li, C.; Li, M.; Tang, X.; Hu, R.; Chen, Y.; et al. Modified behavioural tests to detect white matter injury- induced motor deficits after intracerebral haemorrhage in mice. Sci. Rep. 2019, 9, 16958. [Google Scholar] [CrossRef]
- Zausinger, S.; Hungerhuber, E.; Baethmann, A.; Reulen, H.-J.; Schmid-Elsaesser, R. Neurological impairment in rats after transient middle cerebral artery occlusion: A comparative study under various treatment paradigms. Brain Res. 2000, 863, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Visser, J.E.; Bloem, B.R. Role of the basal ganglia in balance control. Neural Plast. 2005, 12, 161–174. [Google Scholar] [CrossRef]
- Florczak-Rzepka, M.; Grond-Ginsbach, C.; Montaner, J.; Steiner, T. Matrix metalloproteinases in human spontaneous intracerebral hemorrhage: An update. Cerebrovasc. Dis. 2012, 34, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Lucivero, V.; Prontera, M.; Mezzapesa, D.M.; Petruzzellis, M.; Sancilio, M.; Tinelli, A.; Di Noia, D.; Ruggieri, M.; Federico, F. Different roles of matrix metallopro-teinases-2 and -9 after human ischaemic stroke. Neurol. Sci. 2007, 28, 165–170. [Google Scholar] [CrossRef]
- Lakhan, S.E.; Kirchgessner, A.; Tepper, D.; Leonard, A. Matrix metalloproteinases and blood-brain barrier disruption in acute ischemic stroke. Front. Neurol. 2013, 4, 32. [Google Scholar] [CrossRef]
- Wasserman, J.K.; Schlichter, L.C. Minocycline protects the blood–brain barrier and reduces edema following intracerebral hemorrhage in the rat. Exp. Neurol. 2007, 207, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Keep, R.F.; Zhou, N.; Xiang, J.; Andjelkovic, A.V.; Hua, Y.; Xi, G. Vascular disruption and blood-brain barrier dysfunction in intrac-erebral hemorrhage. Fluids Barriers CNS 2014, 11, 18. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Chen, C.; Zhang, J.; Hu, Z. Mechanism and Therapy of Brain Edema after Intracerebral Hemorrhage. Cerebrovasc. Dis. 2016, 42, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.; Wang, Y.-C.; Bai, Y.-Y.; Lu, C.-Q.; Xu, T.-T.; Zhu, L.; Ju, S. Role of P38 MAPK on MMP Activity in Photothrombotic Stroke Mice as Measured using an Ultrafast MMP Activatable Probe. Sci. Rep. 2015, 5, 16951. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Gao, Q.; Ma, Y.; Wang, F.; Tan, X.; Song, D.; Hoo, R.L.; Wang, Z.; Ge, X.; Han, H.; et al. An MMP-9 exclusive neutralizing antibody attenuates blood-brain barrier breakdown in mice with stroke and reduces stroke patient-derived MMP-9 activity. Pharmacol. Res. 2023, 190, 106720. [Google Scholar] [CrossRef]
- Rosenberg, G.A.; Yang, Y. Vasogenic edema due to tight junction disruption by matrix metalloproteinases in cerebral ischemia. Neurosurg. Focus 2007, 22, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Li, Z.; Wu, T.; Ding, S. Role of rho kinase in microvascular damage following cerebral ischemia reperfusion in rats. Int. J. Mol. Sci. 2011, 12, 1222–1231. [Google Scholar] [CrossRef]
- Clark, A.W.; Krekoski, C.A.; Bou, S.-S.; Chapman, K.R.; Edwards, D.R. Increased gelatinase A (MMP-2) and gelatinase B (MMP-9) activities in human brain after focal ischemia. Neurosci. Lett. 1997, 238, 53–56. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, Y.-C.; Chang, C.-H.; Chong, Y.B.; Wu, C.-H.; Tsai, H.-P.; Cheng, T.-L.; Lin, C.-L. MicroRNA-195-5p Attenuates Intracerebral-Hemorrhage-Induced Brain Damage by Inhibiting MMP-9/MMP-2 Expression. Biomedicines 2024, 12, 1373. https://doi.org/10.3390/biomedicines12061373
Tsai Y-C, Chang C-H, Chong YB, Wu C-H, Tsai H-P, Cheng T-L, Lin C-L. MicroRNA-195-5p Attenuates Intracerebral-Hemorrhage-Induced Brain Damage by Inhibiting MMP-9/MMP-2 Expression. Biomedicines. 2024; 12(6):1373. https://doi.org/10.3390/biomedicines12061373
Chicago/Turabian StyleTsai, Yi-Cheng, Chih-Hui Chang, Yoon Bin Chong, Chieh-Hsin Wu, Hung-Pei Tsai, Tian-Lu Cheng, and Chih-Lung Lin. 2024. "MicroRNA-195-5p Attenuates Intracerebral-Hemorrhage-Induced Brain Damage by Inhibiting MMP-9/MMP-2 Expression" Biomedicines 12, no. 6: 1373. https://doi.org/10.3390/biomedicines12061373
APA StyleTsai, Y.-C., Chang, C.-H., Chong, Y. B., Wu, C.-H., Tsai, H.-P., Cheng, T.-L., & Lin, C.-L. (2024). MicroRNA-195-5p Attenuates Intracerebral-Hemorrhage-Induced Brain Damage by Inhibiting MMP-9/MMP-2 Expression. Biomedicines, 12(6), 1373. https://doi.org/10.3390/biomedicines12061373