
The Sigma-1 Receptor Exacerbates Cardiac Dysfunction Induced by Obstructive Nephropathy: A Role for Sexual Dimorphism

 , , ,
, , ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Study Animals
2.2. UUO Surgical Model and Pharmacological Administration of Sigmar1 Agonists and Antagonist
2.3. Colorimetric Tests for Renal Function
2.4. RNA Extraction and RT-qPCR
2.5. Histology and Immunohistochemistry (IHC)
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. RNA Sequencing (RNA-Seq) and Bioinformatic Analysis
2.8. Statistical Analysis
3. Results
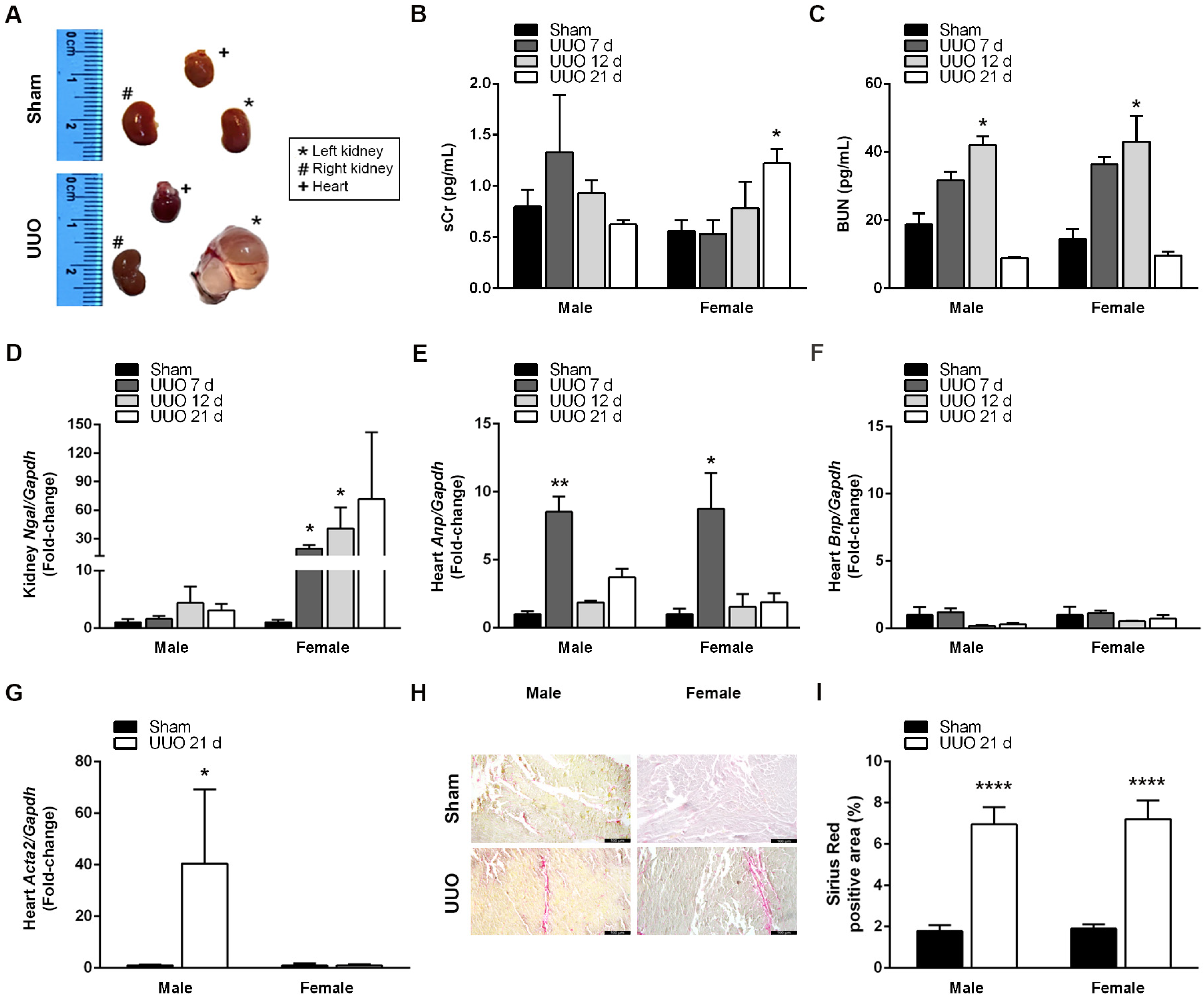
3.1. UUO Effectively Induces CRS4 Development
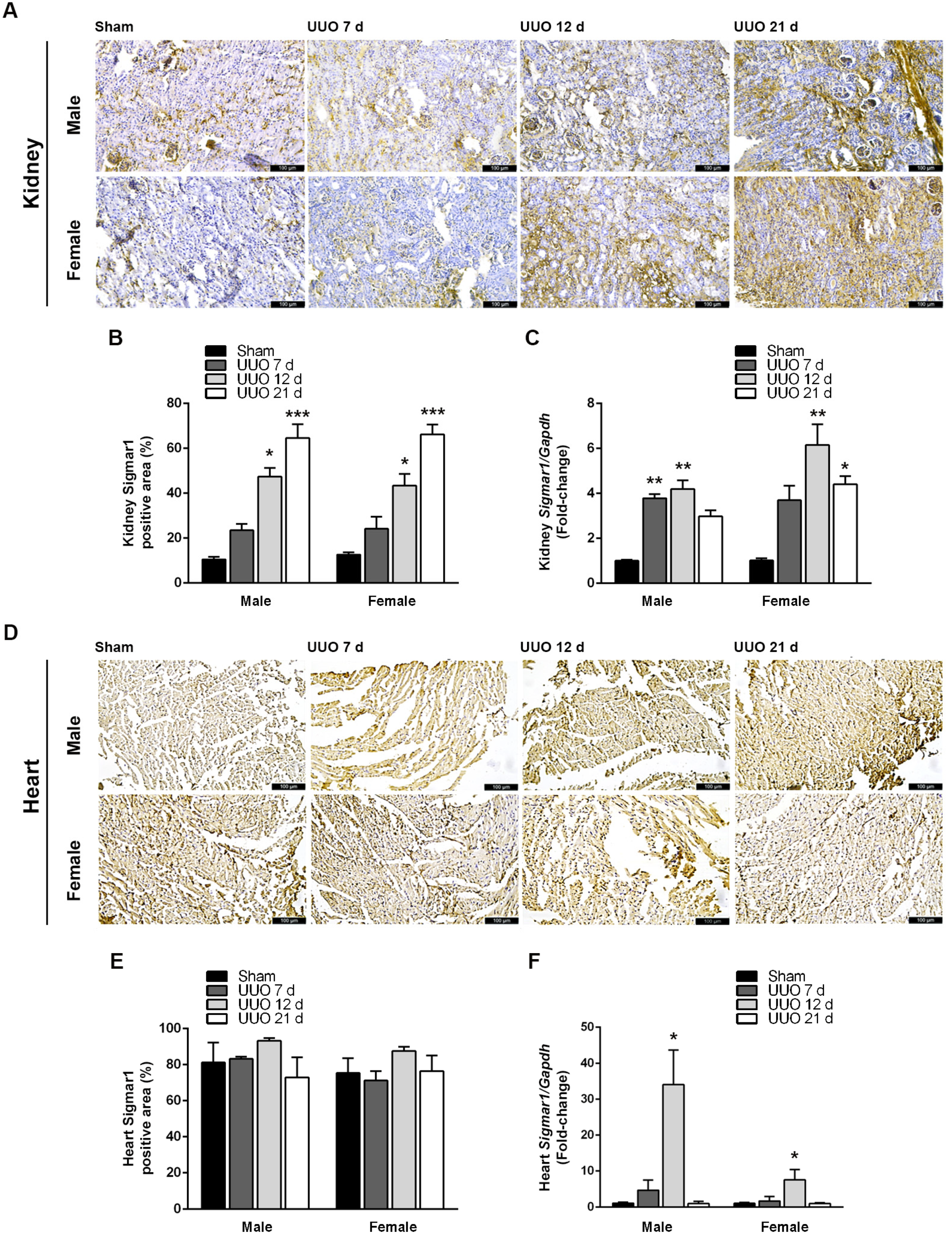
3.2. Obstructive Nephropathy Elevates Sigmar1 Expression in the Kidneys and Heart
3.3. Male Sex Potentiates Sigmar1 Effects on Renal and Cardiac Function Markers after UUO
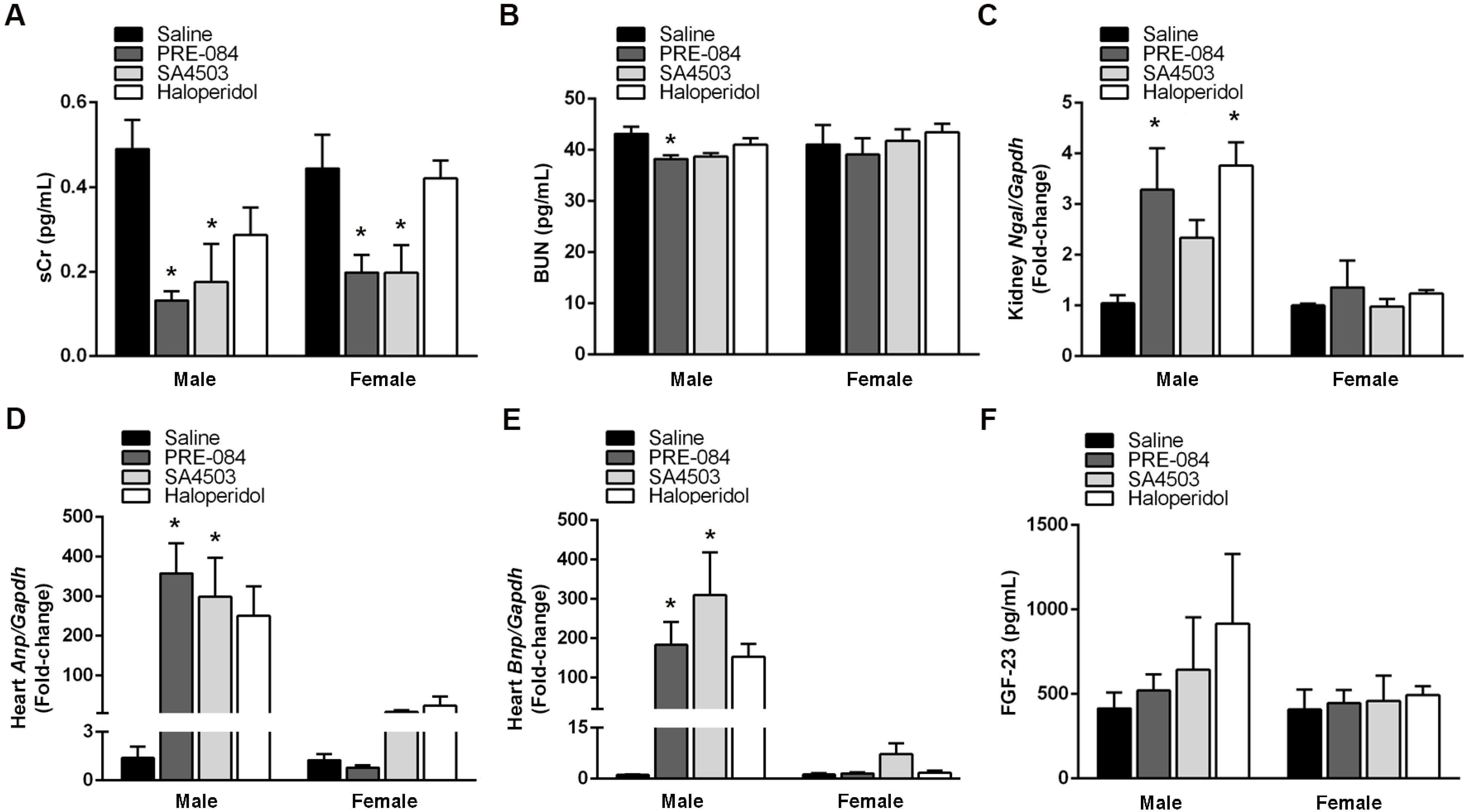
3.4. Sigmar1 Agonists and Antagonists Potentiate UUO-Induced Cardiac Fibrosis Regardless of Sex
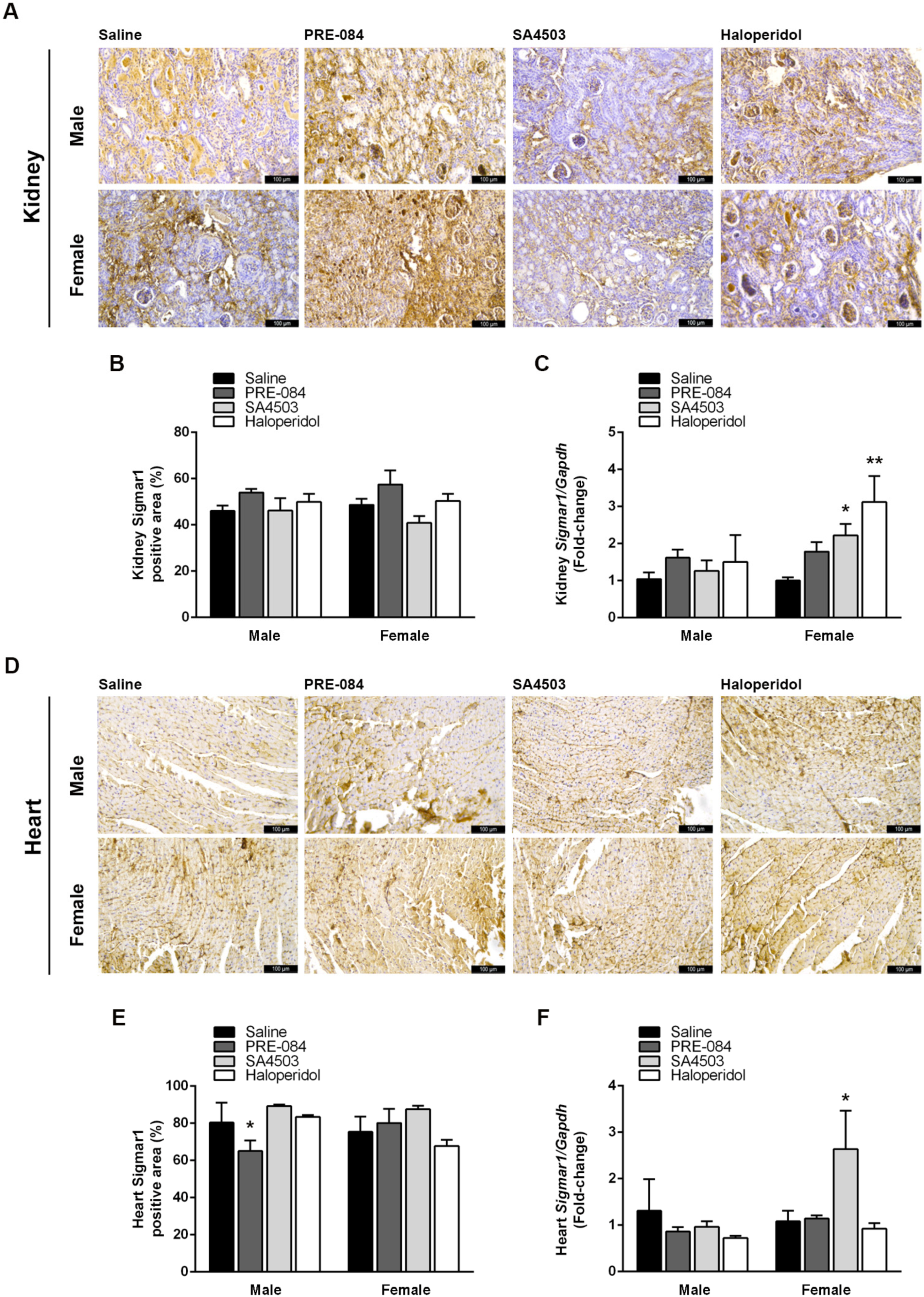
3.5. Sigmar1 Expression in the Kidneys and Heart Is Modulated by Its Agonists and Antagonists after UUO
3.6. Sigmar1 Agonist PRE-084 Induces Transcriptomic Changes Associated with Pathological Cardiac Remodeling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jankowski, J.; Floege, J.; Fliser, D.; Bohm, M.; Marx, N. Cardiovascular Disease in Chronic Kidney Disease: Pathophysiological Insights and Therapeutic Options. Circulation 2021, 143, 1157–1172. [Google Scholar] [CrossRef]
- Minciunescu, A.; Genovese, L.; deFilippi, C. Cardiovascular Alterations and Structural Changes in the Setting of Chronic Kidney Disease: A Review of Cardiorenal Syndrome Type 4. SN Compr. Clin. Med. 2023, 5, 15. [Google Scholar] [CrossRef]
- Rangaswami, J.; Bhalla, V.; Blair, J.E.A.; Chang, T.I.; Costa, S.; Lentine, K.L.; Lerma, E.V.; Mezue, K.; Molitch, M.; Mullens, W.; et al. Cardiorenal Syndrome: Classification, Pathophysiology, Diagnosis, and Treatment Strategies: A Scientific Statement from the American Heart Association. Circulation 2019, 139, e840–e878. [Google Scholar] [CrossRef] [PubMed]
- Gallazzini, M.; Pallet, N. Endoplasmic reticulum stress and kidney dysfunction. Biol. Cell 2018, 110, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Su, T.P.; Hayashi, T. Understanding the molecular mechanism of sigma-1 receptors: Towards a hypothesis that sigma-1 receptors are intracellular amplifiers for signal transduction. Curr. Med. Chem. 2003, 10, 2073–2080. [Google Scholar] [CrossRef]
- Hayashi, T. The Sigma-1 Receptor in Cellular Stress Signaling. Front. Neurosci. 2019, 13, 733. [Google Scholar] [CrossRef] [PubMed]
- Mavlyutov, T.A.; Ruoho, A.E. Ligand-dependent localization and intracellular stability of sigma-1 receptors in CHO-K1 cells. J. Mol. Signal 2007, 2, 8. [Google Scholar] [CrossRef]
- Su, T.P.; Hayashi, T.; Maurice, T.; Buch, S.; Ruoho, A.E. The sigma-1 receptor chaperone as an inter-organelle signaling modulator. Trends Pharmacol. Sci. 2010, 31, 557–566. [Google Scholar] [CrossRef]
- Gomez-Soler, M.; Fernandez-Duenas, V.; Portillo-Salido, E.; Perez, P.; Zamanillo, D.; Vela, J.M.; Burgueno, J.; Ciruela, F. Predicting the antinociceptive efficacy of σ1 receptor ligands by a novel receptor fluorescence resonance energy transfer (FRET) based biosensor. J. Med. Chem. 2014, 57, 238–242. [Google Scholar] [CrossRef]
- Mishra, A.K.; Mavlyutov, T.; Singh, D.R.; Biener, G.; Yang, J.; Oliver, J.A.; Ruoho, A.; Raicu, V. The sigma-1 receptors are present in monomeric and oligomeric forms in living cells in the presence and absence of ligands. Biochem. J. 2015, 466, 263–271. [Google Scholar] [CrossRef]
- Schmidt, H.R.; Betz, R.M.; Dror, R.O.; Kruse, A.C. Structural basis for σ1 receptor ligand recognition. Nat. Struct. Mol. Biol. 2018, 25, 981–987. [Google Scholar] [CrossRef]
- Munguia-Galaviz, F.J.; Miranda-Diaz, A.G.; Cardenas-Sosa, M.A.; Echavarria, R. Sigma-1 Receptor Signaling: In Search of New Therapeutic Alternatives for Cardiovascular and Renal Diseases. Int. J. Mol. Sci. 2023, 24, 1997. [Google Scholar] [CrossRef]
- Balafa, O.; Fernandez-Fernandez, B.; Ortiz, A.; Dounousi, E.; Ekart, R.; Ferro, C.J.; Mark, P.B.; Valdivielso, J.M.; Del Vecchio, L.; Mallamaci, F. Sex disparities in mortality and cardiovascular outcomes in chronic kidney disease. Clin. Kidney J. 2024, 17, sfae044. [Google Scholar] [CrossRef]
- Cobo Marcos, M.; de la Espriella, R.; Gayan Ordas, J.; Zegri, I.; Pomares, A.; Llacer, P.; Fort, A.; Rodriguez Chavarri, A.; Mendez, A.; Blazquez, Z.; et al. Sex differences in Cardiorenal Syndrome: Insights from CARDIOREN Registry. Curr. Heart Fail. Rep. 2023, 20, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Lau, E.S.; Binek, A.; Parker, S.J.; Shah, S.H.; Zanni, M.V.; Van Eyk, J.E.; Ho, J.E. Sexual Dimorphism in Cardiovascular Biomarkers: Clinical and Research Implications. Circ. Res. 2022, 130, 578–592. [Google Scholar] [CrossRef] [PubMed]
- Hosszu, A.; Antal, Z.; Veres-Szekely, A.; Lenart, L.; Balogh, D.B.; Szkibinszkij, E.; Illesy, L.; Hodrea, J.; Banki, N.F.; Wagner, L.; et al. The role of Sigma-1 receptor in sex-specific heat shock response in an experimental rat model of renal ischaemia/reperfusion injury. Transpl. Int. 2018, 31, 1268–1278. [Google Scholar] [CrossRef]
- Wilbert-Lampen, U.; Seliger, C.; Trapp, A.; Straube, F.; Plasse, A. Female sex hormones decrease constitutive endothelin-1 release via endothelial sigma-1/cocaine receptors: An action independent of the steroid hormone receptors. Endothelium 2005, 12, 185–191. [Google Scholar] [CrossRef]
- Munguia-Galaviz, F.J.; Gutierrez-Mercado, Y.K.; Miranda-Diaz, A.G.; Portilla de Buen, E.; Flores-Soto, M.E.; Echavarria, R. Cardiac transcriptomic changes induced by early CKD in mice reveal novel pathways involved in the pathogenesis of Cardiorenal syndrome type 4. Heliyon 2024, 10, e27468. [Google Scholar] [CrossRef]
- Hesketh, E.E.; Vernon, M.A.; Ding, P.; Clay, S.; Borthwick, G.; Conway, B.; Hughes, J. A murine model of irreversible and reversible unilateral ureteric obstruction. J. Vis. Exp. 2014, 94, e52559. [Google Scholar] [CrossRef]
- Kerkela, R.; Ulvila, J.; Magga, J. Natriuretic Peptides in the Regulation of Cardiovascular Physiology and Metabolic Events. J. Am. Heart Assoc. 2015, 4, e002423. [Google Scholar] [CrossRef]
- Delgado-Valero, B.; Cachofeiro, V.; Martinez-Martinez, E. Fibrosis, the Bad Actor in Cardiorenal Syndromes: Mechanisms Involved. Cells 2021, 10, 1824. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.M.; Nikolic-Paterson, D.J.; Lan, H.Y. TGF-beta: The master regulator of fibrosis. Nat. Rev. Nephrol. 2016, 12, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Omi, T.; Tanimukai, H.; Kanayama, D.; Sakagami, Y.; Tagami, S.; Okochi, M.; Morihara, T.; Sato, M.; Yanagida, K.; Kitasyoji, A.; et al. Fluvoxamine alleviates ER stress via induction of Sigma-1 receptor. Cell Death Dis. 2014, 5, e1332. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Liu, X.; Chen, X.; Han, X.; Sun, Y.; Fo, Y.; Wang, X.; Qu, C.; Yang, B. Activation of the sigma-1 receptor exerts cardioprotection in a rodent model of chronic heart failure by stimulation of angiogenesis. Mol. Med. 2022, 28, 87. [Google Scholar] [CrossRef] [PubMed]
- Hosszu, A.; Antal, Z.; Lenart, L.; Hodrea, J.; Koszegi, S.; Balogh, D.B.; Banki, N.F.; Wagner, L.; Denes, A.; Hamar, P.; et al. sigma1-Receptor Agonism Protects against Renal Ischemia-Reperfusion Injury. J. Am. Soc. Nephrol. 2017, 28, 152–165. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.M.; Cifuentes-Diaz, C.; Devaux, J.; Garcia, M.; Bureau, J.; Thomasseau, S.; Klingler, E.; Girault, J.A.; Goutebroze, L. Schwannomin-interacting Protein 1 Isoform IQCJ-SCHIP1 Is a Multipartner Ankyrin- and Spectrin-binding Protein Involved in the Organization of Nodes of Ranvier. J. Biol. Chem. 2017, 292, 2441–2456. [Google Scholar] [CrossRef]
- Kim, J.; Roh, J.D.; Kim, S.; Kang, H.; Bae, M.; Kim, E. Slc6a20a Heterozygous and Homozygous Mutant Mice Display Differential Behavioral and Transcriptomic Changes. Front. Mol. Neurosci. 2022, 15, 857820. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Wilkie, M.; Smith, G.; Paine, M.J. Identification of a functionally impaired allele of human novel oxidoreductase 1 (NDOR1), NDOR1*1. Pharmacogenet Genom. 2005, 15, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Zhangsun, Z.; Dong, Y.; Tang, J.; Jin, Z.; Lei, W.; Wang, C.; Cheng, Y.; Wang, B.; Yang, Y.; Zhao, H. FPR1: A critical gatekeeper of the heart and brain. Pharmacol. Res. 2024, 202, 107125. [Google Scholar] [CrossRef]
- Su, W.; Wang, R.C.; Lohano, M.K.; Wang, L.; Zhu, P.; Luo, Y.; Guo, L.J.; Lv, Q.; Jiang, H.; Wang, J.H.; et al. Identification of Two Mutations in PCDHGA4 and SLFN14 Genes in an Atrial Septal Defect Family. Curr. Med. Sci. 2018, 38, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, J.; Guttsches, A.K.; Schara-Schmidt, U.; Vorgerd, M.; Heute, C.; Preusse, C.; Stenzel, W.; Roos, A. ANO5-related muscle diseases: From clinics and genetics to pathology and research strategies. Genes Dis. 2022, 9, 1506–1520. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Peng, X.; Cao, Y.; Zhou, Y.; Sun, Y. ADNP promotes neural differentiation by modulating Wnt/beta-catenin signaling. Nat. Commun. 2020, 11, 2984. [Google Scholar] [CrossRef]
- Yang, Z.; Zhou, L.; Si, T.; Chen, S.; Liu, C.; Ng, K.K.; Wang, Z.; Chen, Z.; Qiu, C.; Liu, G.; et al. Lysyl hydroxylase LH1 promotes confined migration and metastasis of cancer cells by stabilizing Septin2 to enhance actin network. Mol. Cancer 2023, 22, 21. [Google Scholar] [CrossRef] [PubMed]
- Grayson, C.; Mailloux, R.J. Coenzyme Q(10) and nicotinamide nucleotide transhydrogenase: Sentinels for mitochondrial hydrogen peroxide signaling. Free Radic. Biol. Med. 2023, 208, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Lu, Z.; Wang, F.; Jiang, Z.; Lu, L.; Miao, N.; Wang, N. Renalase attenuates hypertension, renal injury and cardiac remodelling in rats with subtotal nephrectomy. J. Cell Mol. Med. 2016, 20, 1106–1117. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Madduma Hewage, S.; Wijerathne, C.U.B.; Siow, Y.L.; Isaak, C.K.; O, K. Kidney Ischemia-Reperfusion Elicits Acute Liver Injury and Inflammatory Response. Front. Med. 2020, 7, 201. [Google Scholar] [CrossRef] [PubMed]
- Hewitson, T.D.; Ono, T.; Becker, G.J. Small animal models of kidney disease: A review. Methods Mol. Biol. 2009, 466, 41–57. [Google Scholar] [CrossRef]
- Chinnappa, S.; White, E.; Lewis, N.; Baldo, O.; Tu, Y.K.; Glorieux, G.; Vanholder, R.; El Nahas, M.; Mooney, A. Early and asymptomatic cardiac dysfunction in chronic kidney disease. Nephrol. Dial. Transpl. 2018, 33, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zuo, L.; Wang, F.; Wang, M.; Wang, S.; Lv, J.; Liu, L.; Wang, H. Cardiovascular disease in early stages of chronic kidney disease in a Chinese population. J. Am. Soc. Nephrol. 2006, 17, 2617–2621. [Google Scholar] [CrossRef] [PubMed]
- Becker, G.J.; Hewitson, T.D. Animal models of chronic kidney disease: Useful but not perfect. Nephrol. Dial. Transpl. 2013, 28, 2432–2438. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Klimova, E.; Aparicio-Trejo, O.E.; Tapia, E.; Pedraza-Chaverri, J. Unilateral Ureteral Obstruction as a Model to Investigate Fibrosis-Attenuating Treatments. Biomolecules 2019, 9, 141. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, Z.; Chen, Y.; Wu, H.; Zhang, S.; Chen, X. Prediction Value of Serum NGAL in the Diagnosis and Prognosis of Experimental Acute and Chronic Kidney Injuries. Biomolecules 2020, 10, 981. [Google Scholar] [CrossRef]
- Castillo-Rodriguez, E.; Fernandez-Prado, R.; Martin-Cleary, C.; Pizarro-Sanchez, M.S.; Sanchez-Nino, M.D.; Sanz, A.B.; Fernandez-Fernandez, B.; Ortiz, A. Kidney Injury Marker 1 and Neutrophil Gelatinase-Associated Lipocalin in Chronic Kidney Disease. Nephron 2017, 136, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Lai, S.; Dimko, M.; Galani, A.; Coppola, B.; Innico, G.; Frassetti, N.; Mazzei, E.D.; Mariotti, A. Early markers of cardiovascular risk in chronic kidney disease. Ren. Fail. 2015, 37, 254–261. [Google Scholar] [CrossRef]
- Chen, C.; Xie, C.; Wu, H.; Wu, L.; Zhu, J.; Mao, H.; Xing, C. Uraemic Cardiomyopathy in Different Mouse Models. Front. Med. 2021, 8, 690517. [Google Scholar] [CrossRef] [PubMed]
- Ham, O.; Jin, W.; Lei, L.; Huang, H.H.; Tsuji, K.; Huang, M.; Roh, J.; Rosenzweig, A.; Lu, H.A.J. Pathological cardiac remodeling occurs early in CKD mice from unilateral urinary obstruction, and is attenuated by Enalapril. Sci. Rep. 2018, 8, 16087. [Google Scholar] [CrossRef] [PubMed]
- Milardovic, I.; Vitlov Uljevic, M.; Vukojevic, K.; Kostic, S.; Filipovic, N. Renal expression of sigma 1 receptors in diabetic rats. Acta Histochem. 2020, 122, 151580. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, C.S.; Alam, S.; Aishwarya, R.; Miriyala, S.; Panchatcharam, M.; Bhuiyan, M.A.N.; Peretik, J.M.; Orr, A.W.; James, J.; Osinska, H.; et al. Cardiac Dysfunction in the Sigma 1 Receptor Knockout Mouse Associated with Impaired Mitochondrial Dynamics and Bioenergetics. J. Am. Heart Assoc. 2018, 7, e009775. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, C.S.; Aishwarya, R.; Alam, S.; Morshed, M.; Remex, N.S.; Nitu, S.; Kolluru, G.K.; Traylor, J.; Miriyala, S.; Panchatcharam, M.; et al. Methamphetamine induces cardiomyopathy by Sigmar1 inhibition-dependent impairment of mitochondrial dynamics and function. Commun. Biol. 2020, 3, 682. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, Q.; Yang, J.Z.; Li, X.W.; Chen, L.J.; Zhang, K.K.; Liu, J.L.; Li, J.H.; Hsu, C.; Chen, L.; et al. Multi-Omics Analysis Reveals the Role of Sigma-1 Receptor in a Takotsubo-like Cardiomyopathy Model. Biomedicines 2023, 11, 2766. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Z.; Zhang, K.K.; Shen, H.W.; Liu, Y.; Li, X.W.; Chen, L.J.; Liu, J.L.; Li, J.H.; Zhao, D.; Wang, Q.; et al. Sigma-1 receptor knockout disturbs gut microbiota, remodels serum metabolome, and exacerbates isoprenaline-induced heart failure. Front. Microbiol. 2023, 14, 1255971. [Google Scholar] [CrossRef] [PubMed]
- Hodrea, J.; Tran, M.N.; Besztercei, B.; Medveczki, T.; Szabo, A.J.; Orfi, L.; Kovacs, I.; Fekete, A. Sigma-1 Receptor Agonist Fluvoxamine Ameliorates Fibrotic Response of Trabecular Meshwork Cells. Int. J. Mol. Sci. 2023, 24, 11646. [Google Scholar] [CrossRef] [PubMed]
- Haritha, C.V.; Lingaraju, M.C.; Mathesh, K.; Jadhav, S.E.; Shyamkumar, T.S.; Aneesha, V.A.; Parida, S.; Singh, T.U.; Kumar, D. PRE-084 ameliorates adenine-induced renal fibrosis in rats. Tissue Cell 2022, 79, 101905. [Google Scholar] [CrossRef]
- Volpe, M. Natriuretic peptides and cardio-renal disease. Int. J. Cardiol. 2014, 176, 630–639. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, D.; Kudoh, S.; Shiojima, I.; Zou, Y.; Harada, K.; Shimoyama, M.; Imai, Y.; Monzen, K.; Yamazaki, T.; Yazaki, Y.; et al. Atrial natriuretic peptide inhibits cardiomyocyte hypertrophy through mitogen-activated protein kinase phosphatase-1. Biochem. Biophys. Res. Commun. 2004, 322, 310–319. [Google Scholar] [CrossRef]
- Calvieri, C.; Rubattu, S.; Volpe, M. Molecular mechanisms underlying cardiac antihypertrophic and antifibrotic effects of natriuretic peptides. J. Mol. Med. 2012, 90, 5–13. [Google Scholar] [CrossRef]
- Kapoun, A.M.; Liang, F.; O’Young, G.; Damm, D.L.; Quon, D.; White, R.T.; Munson, K.; Lam, A.; Schreiner, G.F.; Protter, A.A. B-type natriuretic peptide exerts broad functional opposition to transforming growth factor-beta in primary human cardiac fibroblasts: Fibrosis, myofibroblast conversion, proliferation, and inflammation. Circ. Res. 2004, 94, 453–461. [Google Scholar] [CrossRef]
- Vinnakota, S.; Chen, H.H. The Importance of Natriuretic Peptides in Cardiometabolic Diseases. J. Endocr. Soc. 2020, 4, bvaa052. [Google Scholar] [CrossRef]
- Hansell, P.; Fasching, A.; Sjoquist, M.; Anden, N.E.; Ulfendahl, H.R. The dopamine receptor antagonist haloperidol blocks natriuretic but not hypotensive effects of the atrial natriuretic factor. Acta Physiol. Scand. 1987, 130, 401–407. [Google Scholar] [CrossRef]
- Choi, M.R.; Rukavina Mikusic, N.L.; Kouyoumdzian, N.M.; Kravetz, M.C.; Fernandez, B.E. Atrial natriuretic peptide and renal dopaminergic system: A positive friendly relationship? Biomed. Res. Int. 2014, 2014, 710781. [Google Scholar] [CrossRef]
- Madularu, D.; Kulkarni, P.; Yee, J.R.; Kenkel, W.M.; Shams, W.M.; Ferris, C.F.; Brake, W.G. High estrogen and chronic haloperidol lead to greater amphetamine-induced BOLD activation in awake, amphetamine-sensitized female rats. Horm. Behav. 2016, 82, 56–63. [Google Scholar] [CrossRef]
- Qu, J.; Li, M.; Li, D.; Xin, Y.; Li, J.; Lei, S.; Wu, W.; Liu, X. Stimulation of Sigma-1 Receptor Protects against Cardiac Fibrosis by Alleviating IRE1 Pathway and Autophagy Impairment. Oxid. Med. Cell Longev. 2021, 2021, 8836818. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wan, W.; Zhao, X.; Han, X.; Ye, T.; Chen, X.; Ran, Q.; Wang, X.; Liu, X.; Qu, C.; et al. Chronic Sigma 1 receptor activation alleviates right ventricular dysfunction secondary to pulmonary arterial hypertension. Bioengineered 2022, 13, 10843–10856. [Google Scholar] [CrossRef] [PubMed]
- Fo, Y.; Zhang, C.; Chen, X.; Liu, X.; Ye, T.; Guo, Y.; Qu, C.; Shi, S.; Yang, B. Chronic sigma-1 receptor activation ameliorates ventricular remodeling and decreases susceptibility to ventricular arrhythmias after myocardial infarction in rats. Eur. J. Pharmacol. 2020, 889, 173614. [Google Scholar] [CrossRef]
- Ye, T.; Liu, X.; Qu, C.; Zhang, C.; Fo, Y.; Guo, Y.; Chen, X.; Shi, S.; Yang, B. Chronic inhibition of the sigma-1 receptor exacerbates atrial fibrillation susceptibility in rats by promoting atrial remodeling. Life Sci. 2019, 235, 116837. [Google Scholar] [CrossRef] [PubMed]
- Franks, A.; Airoldi, E.; Slavov, N. Post-transcriptional regulation across human tissues. PLoS Comput. Biol. 2017, 13, e1005535. [Google Scholar] [CrossRef] [PubMed]
- Rehman, M.; Vodret, S.; Braga, L.; Guarnaccia, C.; Celsi, F.; Rossetti, G.; Martinelli, V.; Battini, T.; Long, C.; Vukusic, K.; et al. High-throughput screening discovers antifibrotic properties of haloperidol by hindering myofibroblast activation. JCI Insight 2019, 4, e123987. [Google Scholar] [CrossRef] [PubMed]
- Shinoda, Y.; Tagashira, H.; Bhuiyan, M.S.; Hasegawa, H.; Kanai, H.; Fukunaga, K. Haloperidol aggravates transverse aortic constriction-induced heart failure via mitochondrial dysfunction. J. Pharmacol. Sci. 2016, 131, 172–183. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munguia-Galaviz, F.J.; Miranda-Diaz, A.G.; Gutierrez-Mercado, Y.K.; Ku-Centurion, M.; Gonzalez-Gonzalez, R.A.; Portilla-de Buen, E.; Echavarria, R. The Sigma-1 Receptor Exacerbates Cardiac Dysfunction Induced by Obstructive Nephropathy: A Role for Sexual Dimorphism. Biomedicines 2024, 12, 1908. https://doi.org/10.3390/biomedicines12081908
Munguia-Galaviz FJ, Miranda-Diaz AG, Gutierrez-Mercado YK, Ku-Centurion M, Gonzalez-Gonzalez RA, Portilla-de Buen E, Echavarria R. The Sigma-1 Receptor Exacerbates Cardiac Dysfunction Induced by Obstructive Nephropathy: A Role for Sexual Dimorphism. Biomedicines. 2024; 12(8):1908. https://doi.org/10.3390/biomedicines12081908
Chicago/Turabian StyleMunguia-Galaviz, Francisco Javier, Alejandra Guillermina Miranda-Diaz, Yanet Karina Gutierrez-Mercado, Marco Ku-Centurion, Ricardo Arturo Gonzalez-Gonzalez, Eliseo Portilla-de Buen, and Raquel Echavarria. 2024. "The Sigma-1 Receptor Exacerbates Cardiac Dysfunction Induced by Obstructive Nephropathy: A Role for Sexual Dimorphism" Biomedicines 12, no. 8: 1908. https://doi.org/10.3390/biomedicines12081908