
Changes in AmotL2 Expression in Cells of the Human Enteral Nervous System in Oxaliplatin-Induced Enteric Neuropathy
 , , ,
, , ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. Immunohistochemistry
2.3. Image Quantitative Analysis and Statistics
3. Results
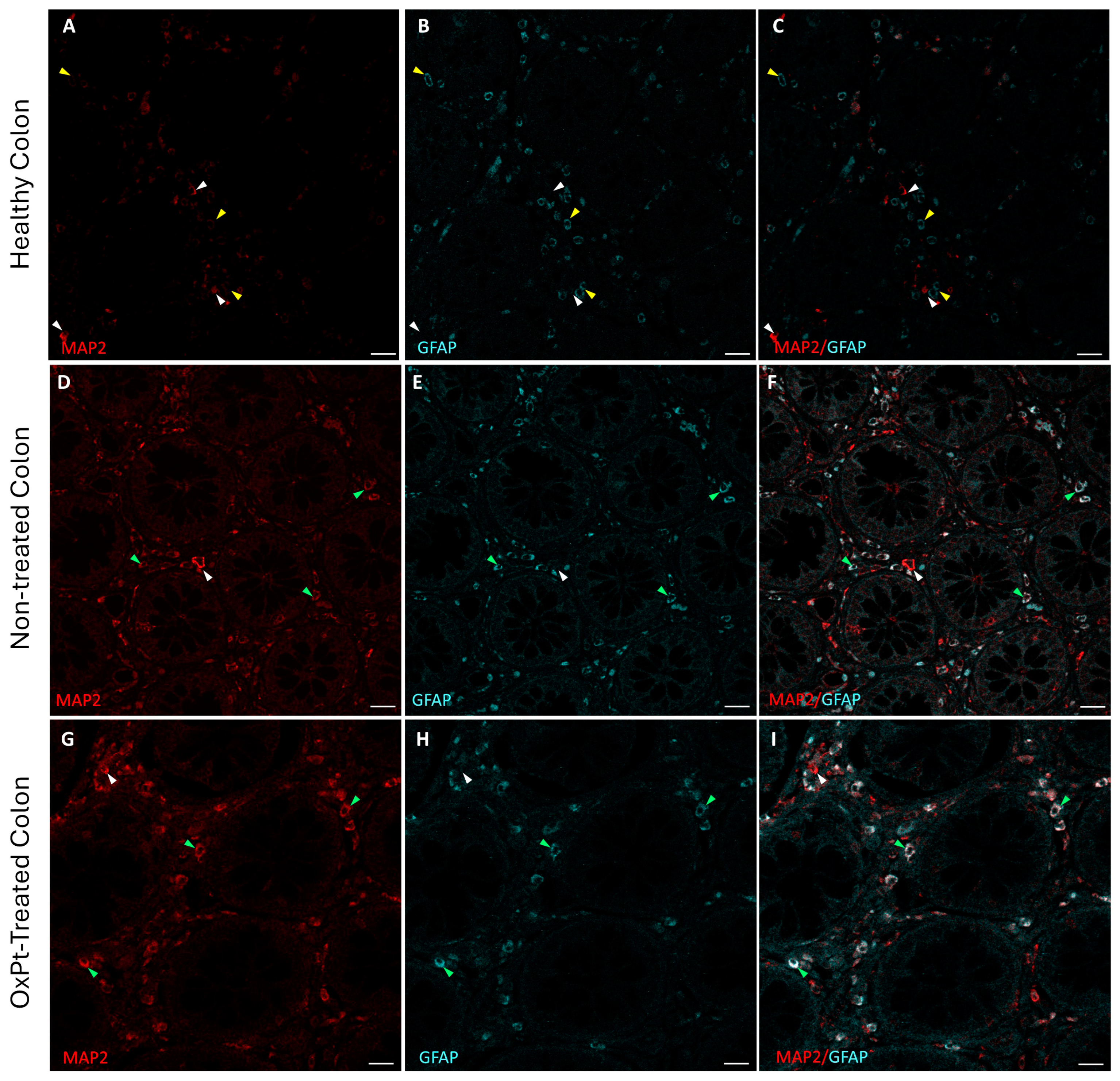
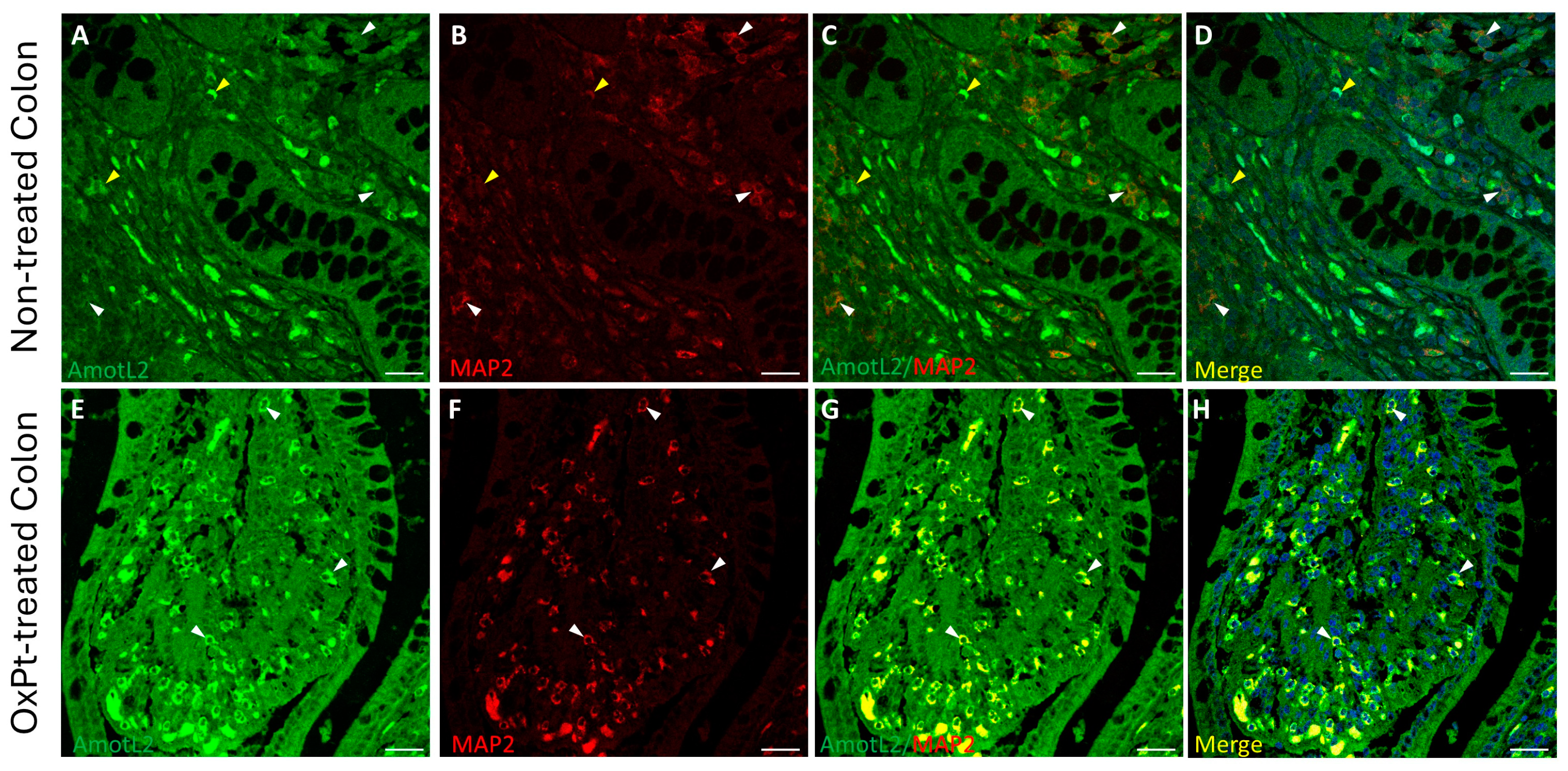
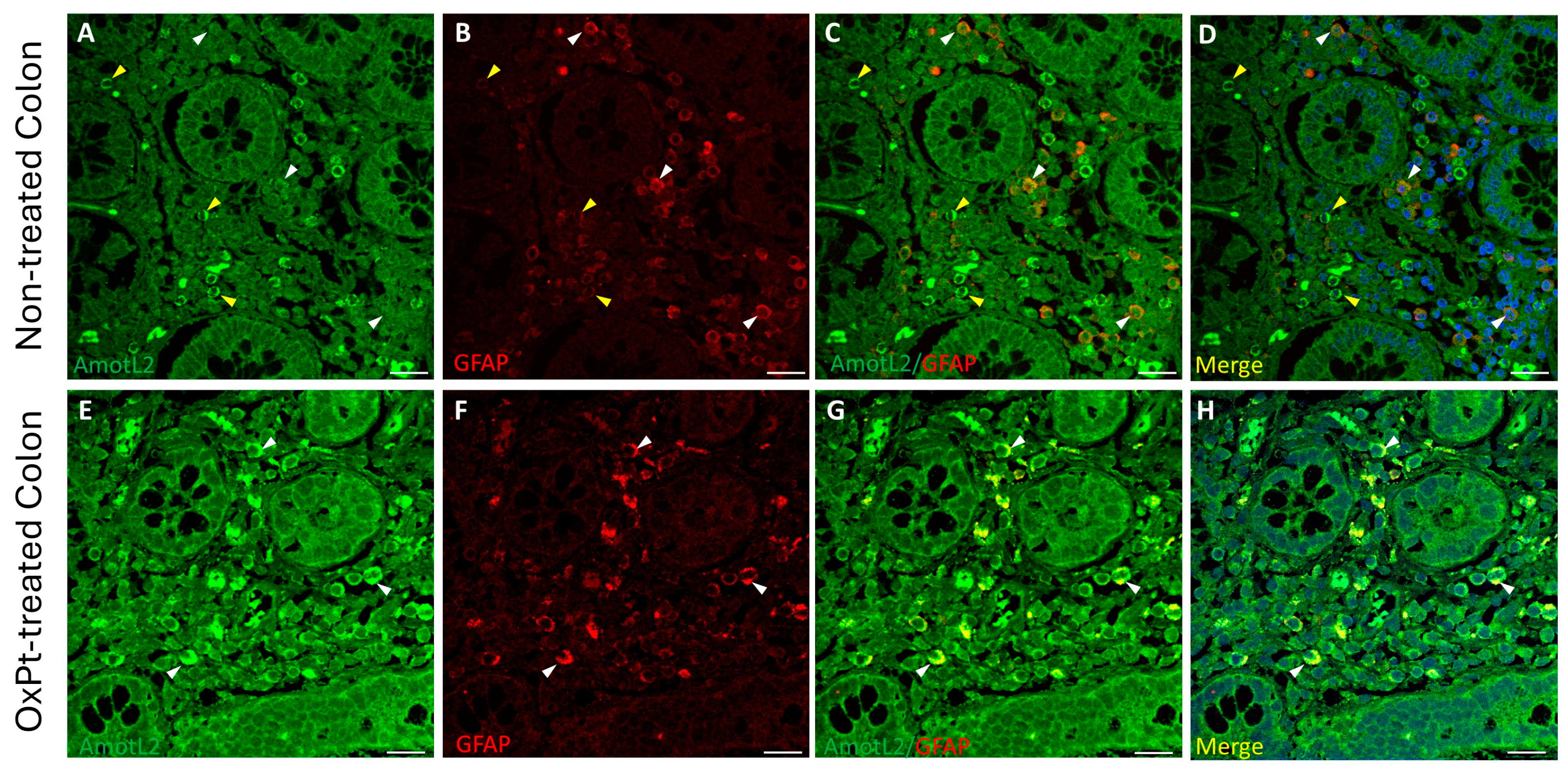
3.1. ENS Neurons and Glial Identification
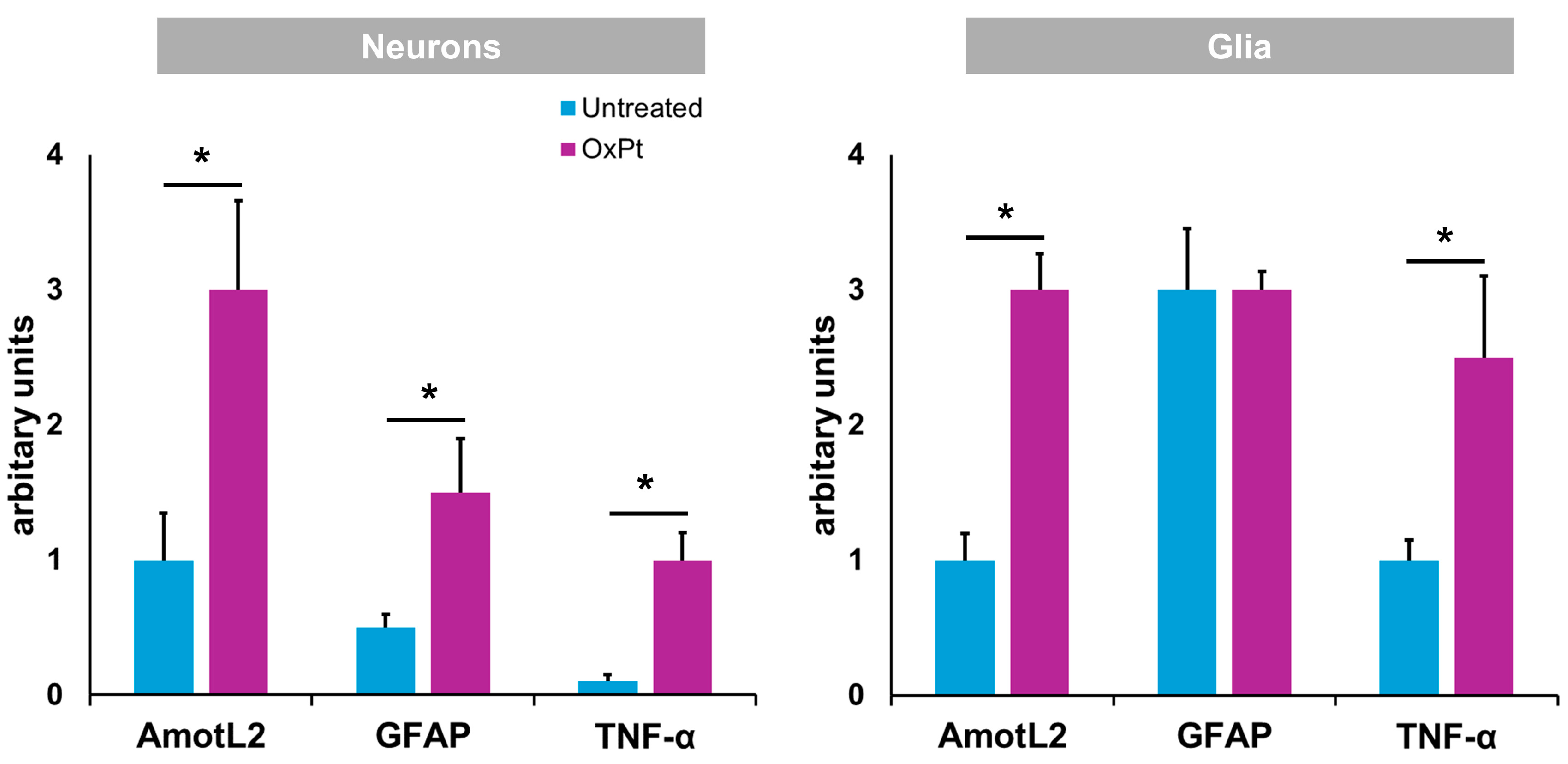
3.2. Quantification of Neurons and Glial Cells of the ENS
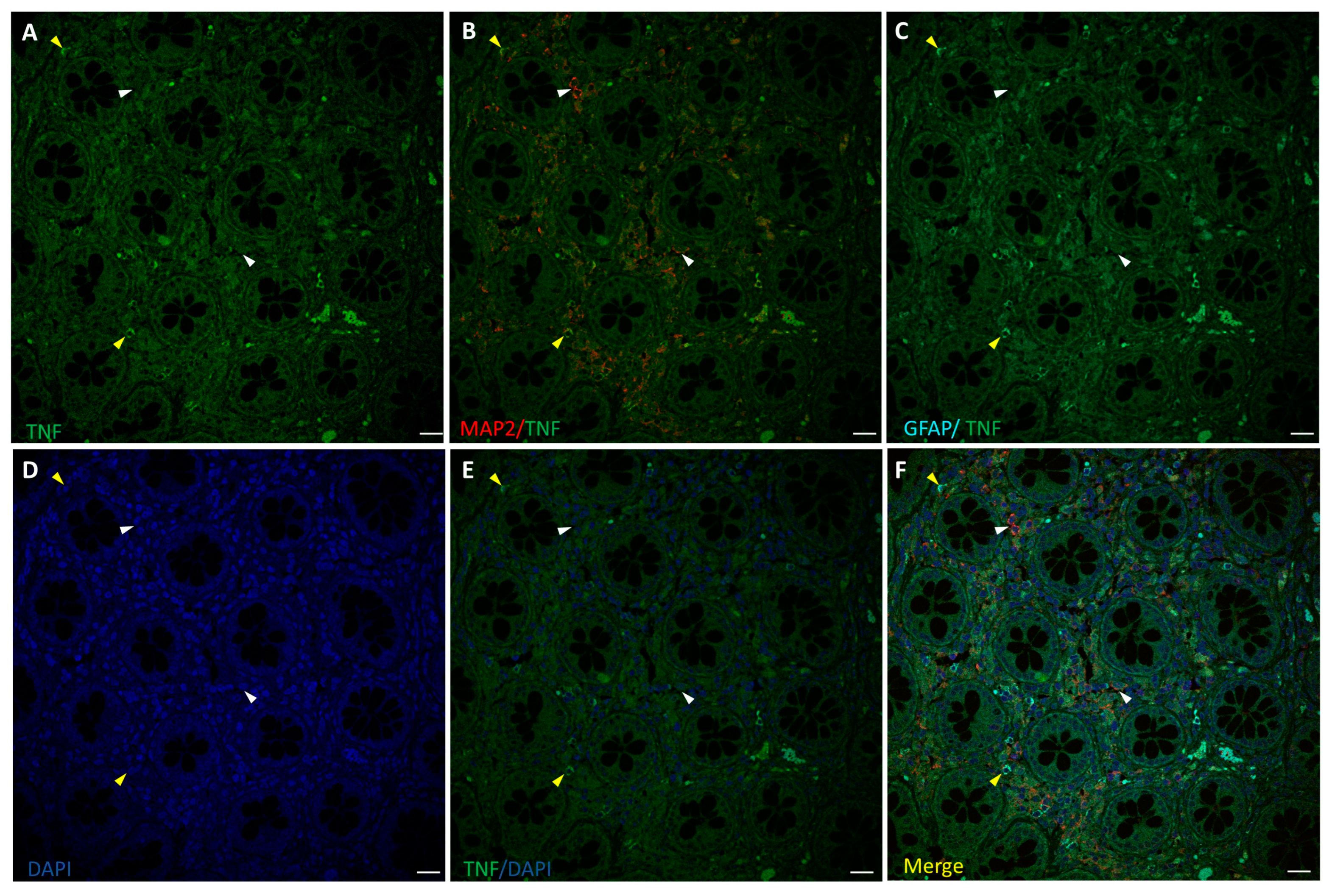
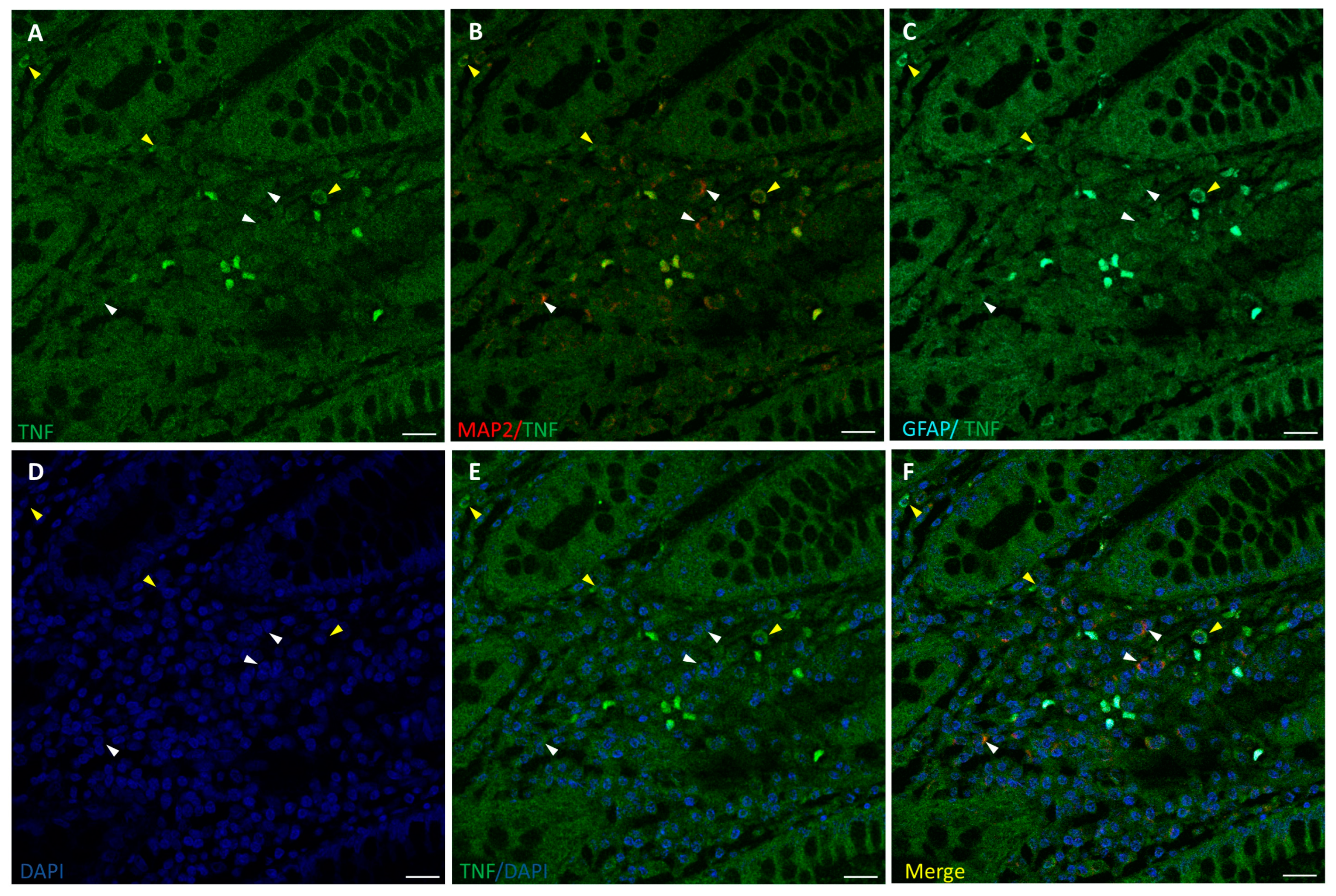
3.3. TNF-α Expression in the ENS
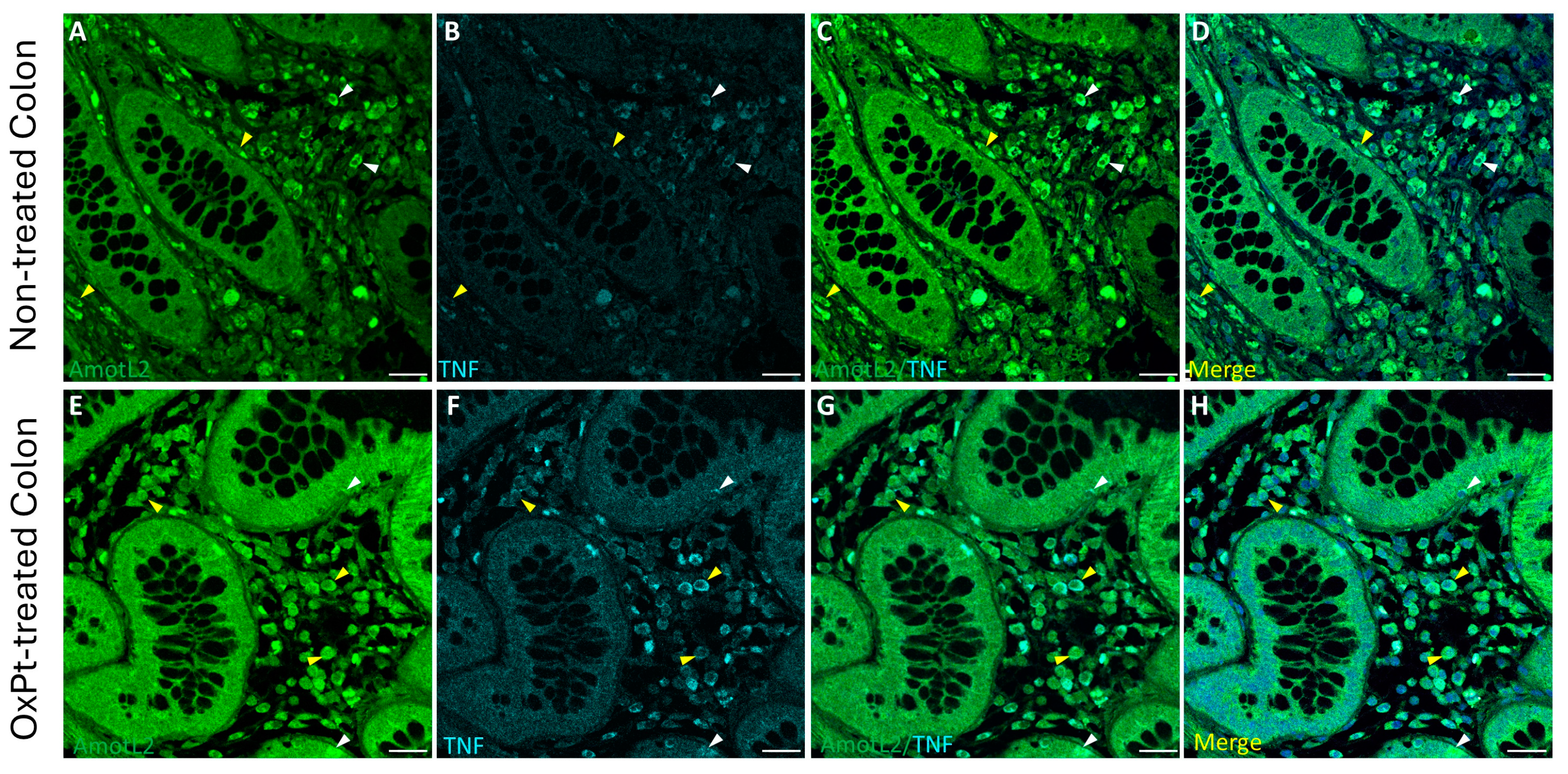
3.4. OxPt Treatment Effects over AmotL2 Expression
3.5. Variations in AmotL2 Expression in the Enteric Nervous System in Samples from Patients Treated with OxPt-Based Chemotherapy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Cutsem, E.; Cervantes, A.; Nordlinger, B.; Arnold, D. Metastatic Colorectal Cancer: ESMO Clinical Practice Guidelines for Diagnosis, Treatment and Follow-Up. Ann. Oncol. 2014, 25, iii1–iii9. [Google Scholar] [CrossRef]
- Argilés, G.; Tabernero, J.; Labianca, R.; Hochhauser, D.; Salazar, R.; Iveson, T.; Laurent-Puig, P.; Quirke, P.; Yoshino, T.; Taieb, J.; et al. Localised Colon Cancer: ESMO Clinical Practice Guidelines for Diagnosis, Treatment and Follow-Up. Ann. Oncol. 2020, 31, 1291–1305. [Google Scholar] [CrossRef]
- Carbone, S.E.; Jovanovska, V.; Brookes, S.J.H.; Nurgali, K. Electrophysiological and Morphological Changes in Colonic Myenteric Neurons from Chemotherapy-treated Patients: A Pilot Study. Neurogastroenterol. Motil. 2016, 28, 975–984. [Google Scholar] [CrossRef]
- Keefe, D.M.; Elting, L.S.; Nguyen, H.T.; Grunberg, S.M.; Aprile, G.; Bonaventura, A.; Selva-Nayagam, S.; Barsevick, A.; Koczwara, B.; Sonis, S.T. Risk and Outcomes of Chemotherapy-Induced Diarrhea (CID) among Patients with Colorectal Cancer Receiving Multi-Cycle Chemotherapy. Cancer Chemother. Pharmacol. 2014, 74, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Peterson, D.E.; Bensadoun, R.-J.; Roila, F. Management of Oral and Gastrointestinal Mucositis: ESMO Clinical Practice Guidelines. Ann. Oncol. 2011, 22, vi78–vi84. [Google Scholar] [CrossRef]
- McQuade, R.M.; Stojanovska, V.; Abalo, R.; Bornstein, J.C.; Nurgali, K. Chemotherapy-Induced Constipation and Diarrhea: Pathophysiology, Current and Emerging Treatments. Front. Pharmacol. 2016, 7, 414. [Google Scholar] [CrossRef] [PubMed]
- Sonis, S.T.; Elting, L.S.; Keefe, D.; Peterson, D.E.; Schubert, M.; Hauer-Jensen, M.; Bekele, B.N.; Raber-Durlacher, J.; Donnelly, J.P.; Rubenstein, E.B. Perspectives on Cancer Therapy-Induced Mucosal Injury. Cancer 2004, 100, 1995–2025. [Google Scholar] [CrossRef] [PubMed]
- Costa, M. Anatomy and Physiology of the Enteric Nervous System. Gut 2000, 47, iv15–iv19. [Google Scholar] [CrossRef]
- Stojanovska, V.; McQuade, R.M.; Miller, S.; Nurgali, K. Effects of Oxaliplatin Treatment on the Myenteric Plexus Innervation and Glia in the Murine Distal Colon. J. Histochem. Cytochem. 2018, 66, 723–736. [Google Scholar] [CrossRef]
- Shaw, A.S.; Filbert, E.L. Scaffold Proteins and Immune-Cell Signalling. Nat. Rev. Immunol. 2009, 9, 47–56. [Google Scholar] [CrossRef]
- Buday, L.; Tompa, P. Functional Classification of Scaffold Proteins and Related Molecules. FEBS J. 2010, 277, 4348–4355. [Google Scholar] [CrossRef]
- Garbett, D.; Bretscher, A. The Surprising Dynamics of Scaffolding Proteins. Mol. Biol. Cell 2014, 25, 2315–2319. [Google Scholar] [CrossRef]
- Wang, H.; Ye, M.; Jin, X. Role of Angiomotin Family Members in Human Diseases (Review). Exp. Ther. Med. 2024, 27, 258. [Google Scholar] [CrossRef]
- Rotoli, D.; Morales, M.; Ávila, J.; Maeso, M.; García, M.; Mobasheri, A.; Martín-Vasallo, P. Commitment of Scaffold Proteins in the Onco-Biology of Human Colorectal Cancer and Liver Metastases after Oxaliplatin-Based Chemotherapy. Int. J. Mol. Sci. 2017, 18, 891. [Google Scholar] [CrossRef]
- Moleirinho, S.; Guerrant, W.; Kissil, J.L. The Angiomotins—From Discovery to Function. FEBS Lett. 2014, 588, 2693–2703. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, F. Angiomotin Family Proteins in the Hippo Signaling Pathway. BioEssays 2024, 46, 2400076. [Google Scholar] [CrossRef] [PubMed]
- Amirifar, P.; Kissil, J. The Role of Motin Family Proteins in Tumorigenesis—An Update. Oncogene 2023, 42, 1265–1271. [Google Scholar] [CrossRef]
- Wigerius, M.; Quinn, D.; Fawcett, J.P. Emerging Roles for Angiomotin in the Nervous System. Sci. Signal 2020, 13, eabc0635. [Google Scholar] [CrossRef] [PubMed]
- Wigerius, M.; Quinn, D.; Diab, A.; Clattenburg, L.; Kolar, A.; Qi, J.; Krueger, S.R.; Fawcett, J.P. The Polarity Protein Angiomotin P130 Controls Dendritic Spine Maturation. J. Cell Biol. 2018, 217, 715. [Google Scholar] [CrossRef]
- Rojek, K.O.; Krzemień, J.; Doleżyczek, H.; Boguszewski, P.M.; Kaczmarek, L.; Konopka, W.; Rylski, M.; Jaworski, J.; Holmgren, L.; Prószyński, T.J. Amot and Yap1 Regulate Neuronal Dendritic Tree Complexity and Locomotor Coordination in Mice. PLoS Biol. 2019, 17, e3000253. [Google Scholar] [CrossRef]
- Zaltsman, Y.; Masuko, S.; Bensen, J.J.; Kiessling, L.L. Angiomotin Regulates YAP Localization during Neural Differentiation of Human Pluripotent Stem Cells. Stem Cell Rep. 2019, 12, 869–877. [Google Scholar] [CrossRef]
- Proszynski, T.J.; Sanes, J.R. Amotl2 Interacts with LL5β, Localizes to Podosomes and Regulates Postsynaptic Differentiation in Muscle. J. Cell Sci. 2013, 126, 2225–2235. [Google Scholar] [CrossRef]
- Furness, J.B. The Enteric Nervous System and Neurogastroenterology. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Stauffer, W.; Sheng, H.; Lim, H.N. EzColocalization: An ImageJ Plugin for Visualizing and Measuring Colocalization in Cells and Organisms. Sci. Rep. 2018, 8, 15764. [Google Scholar] [CrossRef] [PubMed]
- Sasselli, V.; Pachnis, V.; Burns, A.J. The Enteric Nervous System. Dev. Biol. 2012, 366, 1106–1115. [Google Scholar] [CrossRef] [PubMed]
- Cabezos, P.A.; Vera, G.; Martín-fontelles, M.I.; Fernández-pujol, R.; Abalo, R. Cisplatin-Induced Gastrointestinal Dysmotility Is Aggravated after Chronic Administration in the Rat. Comparison with Pica. Neurogastroenterol. Motil. 2010, 22, 797-e225. [Google Scholar] [CrossRef]
- Wafai, L.; Taher, M.; Jovanovska, V.; Bornstein, J.C.; Dass, C.R.; Nurgali, K. Effects of Oxaliplatin on Mouse Myenteric Neurons and Colonic Motility. Front. Neurosci. 2013, 7, 30. [Google Scholar] [CrossRef]
- Vera, G.; Castillo, M.; Cabezos, P.A.; Chiarlone, A.; Martín, M.I.; Gori, A.; Pasquinelli, G.; Barbara, G.; Stanghellini, V.; Corinaldesi, R.; et al. Enteric Neuropathy Evoked by Repeated Cisplatin in the Rat. Neurogastroenterol. Motil. 2011, 23, 370-e163. [Google Scholar] [CrossRef]
- Zhang, D.K.; He, F.Q.; Li, T.K.; Pang, X.H.; Cui, D.J.; Xie, Q.; Huang, X.L.; Gan, H.T. Glial-Derived Neurotrophic Factor Regulates Intestinal Epithelial Barrier Function and Inflammation and Is Therapeutic for Murine Colitis. J. Pathol. 2010, 222, 213–222. [Google Scholar] [CrossRef]
- Bach-Ngohou, K.; Mahé, M.M.; Aubert, P.; Abdo, H.; Boni, S.; Bourreille, A.; Denis, M.G.; Lardeux, B.; Neunlist, M.; Masson, D. Enteric Glia Modulate Epithelial Cell Proliferation and Differentiation through 15-Deoxy-12,14-Prostaglandin J2. J. Physiol. 2010, 588, 2533–2544. [Google Scholar] [CrossRef]
- Casper, K.B.; McCarthy, K.D. GFAP-Positive Progenitor Cells Produce Neurons and Oligodendrocytes throughout the CNS. Mol. Cell. Neurosci. 2006, 31, 676–684. [Google Scholar] [CrossRef]
- Cooper, J.E.; Natarajan, D.; McCann, C.J.; Choudhury, S.; Godwin, H.; Burns, A.J.; Thapar, N. In Vivo Transplantation of Fetal Human Gut-derived Enteric Neural Crest Cells. Neurogastroenterol. Motil. 2017, 29, e12900. [Google Scholar] [CrossRef] [PubMed]
- Lasrado, R.; Boesmans, W.; Kleinjung, J.; Pin, C.; Bell, D.; Bhaw, L.; McCallum, S.; Zong, H.; Luo, L.; Clevers, H.; et al. Lineage-Dependent Spatial and Functional Organization of the Mammalian Enteric Nervous System. Science 2017, 356, 722–726. [Google Scholar] [CrossRef] [PubMed]
- Jessen, K.R.; Mirsky, R.; Arthur-Farraj, P. The Role of Cell Plasticity in Tissue Repair: Adaptive Cellular Reprogramming. Dev. Cell 2015, 34, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Laranjeira, C.; Sandgren, K.; Kessaris, N.; Richardson, W.; Potocnik, A.; Vanden Berghe, P.; Pachnis, V. Glial Cells in the Mouse Enteric Nervous System Can Undergo Neurogenesis in Response to Injury. J. Clin. Investig. 2011, 121, 3412–3424. [Google Scholar] [CrossRef]
- Belkind-Gerson, J.; Graham, H.K.; Reynolds, J.; Hotta, R.; Nagy, N.; Cheng, L.; Kamionek, M.; Shi, H.N.; Aherne, C.M.; Goldstein, A.M. Colitis Promotes Neuronal Differentiation of Sox2+ and PLP1+ Enteric Cells. Sci. Rep. 2017, 7, 2525. [Google Scholar] [CrossRef]
- Heinhuis, K.M.; Ros, W.; Kok, M.; Steeghs, N.; Beijnen, J.H.; Schellens, J.H.M. Enhancing Antitumor Response by Combining Immune Checkpoint Inhibitors with Chemotherapy in Solid Tumors. Ann. Oncol. 2019, 30, 219–235. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, L.; Forschner, A.; Loquai, C.; Goldinger, S.M.; Zimmer, L.; Ugurel, S.; Schmidgen, M.I.; Gutzmer, R.; Utikal, J.S.; Göppner, D.; et al. Cutaneous, Gastrointestinal, Hepatic, Endocrine, and Renal Side-Effects of Anti-PD-1 Therapy. Eur. J. Cancer 2016, 60, 190–209. [Google Scholar] [CrossRef]
- Nardini, P.; Pini, A.; Bessard, A.; Duchalais, E.; Niccolai, E.; Neunlist, M.; Vannucchi, M.G. GLP-2 Prevents Neuronal and Glial Changes in the Distal Colon of Mice Chronically Treated with Cisplatin. Int. J. Mol. Sci. 2020, 21, 8875. [Google Scholar] [CrossRef]
- Stojanovska, V.; Sakkal, S.; Nurgali, K. Platinum-Based Chemotherapy: Gastrointestinal Immunomodulation and Enteric Nervous System Toxicity. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G223–G232. [Google Scholar] [CrossRef]
- Brown, I.A.M.; McClain, J.L.; Watson, R.E.; Patel, B.A.; Gulbransen, B.D. Enteric Glia Mediate Neuron Death in Colitis Through Purinergic Pathways That Require Connexin-43 and Nitric Oxide. Cell Mol. Gastroenterol. Hepatol. 2016, 2, 77–91. [Google Scholar] [CrossRef]
- Pochard, C.; Coquenlorge, S.; Freyssinet, M.; Naveilhan, P.; Bourreille, A.; Neunlist, M.; Rolli-Derkinderen, M. The Multiple Faces of Inflammatory Enteric Glial Cells: Is Crohn’s Disease a Gliopathy? Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G1–G11. [Google Scholar] [CrossRef] [PubMed]
- Bush, T.G.; Savidge, T.C.; Freeman, T.C.; Cox, H.J.; Campbell, E.A.; Mucke, L.; Johnson, M.H.; Sofroniew, M.V. Fulminant Jejuno-Ileitis Following Ablation of Enteric Glia in Adult Transgenic Mice. Cell 1998, 93, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Delvalle, N.M.; Dharshika, C.; Morales-Soto, W.; Fried, D.E.; Gaudette, L.; Gulbransen, B.D. Communication Between Enteric Neurons, Glia, and Nociceptors Underlies the Effects of Tachykinins on Neuroinflammation. Cell Mol. Gastroenterol. Hepatol. 2018, 6, 321–344. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.S.; de Andrade Gomes, Y.; de Sousa Cavalcante, M.L.; Telles, P.V.N.; da Silva, A.C.A.; Severo, J.S.; de Oliveira Santos, R.; dos Santos, B.L.B.; Cavalcante, G.L.; Rocha, C.H.L.; et al. Exercise and Pyridostigmine Prevents Gastric Emptying Delay and Increase Blood Pressure and Cisplatin-Induced Baroreflex Sensitivity in Rats. Life Sci. 2021, 267, 118972. [Google Scholar] [CrossRef]
- Rosenberg, H.J.; Rao, M. Enteric Glia in Homeostasis and Disease: From Fundamental Biology to Human Pathology. iScience 2021, 24, 102863. [Google Scholar] [CrossRef]
- Lv, M.; Shen, Y.; Yang, J.; Li, S.; Wang, B.; Chen, Z.; Li, P.; Liu, P.; Yang, J. Angiomotin Family Members: Oncogenes or Tumor Suppressors? Int. J. Biol. Sci. 2017, 13, 772. [Google Scholar] [CrossRef]
- Woznicki, J.A.; Saini, N.; Flood, P.; Rajaram, S.; Lee, C.M.; Stamou, P.; Skowyra, A.; Bustamante-Garrido, M.; Regazzoni, K.; Crawford, N.; et al. TNF-α Synergises with IFN-γ to Induce Caspase-8-JAK1/2-STAT1-Dependent Death of Intestinal Epithelial Cells. Cell Death Dis. 2021, 12, 864. [Google Scholar] [CrossRef]
- Melfi, S.; Colciago, A.; Giannotti, G.; Bonalume, V.; Caffino, L.; Fumagalli, F.; Magnaghi, V. Stressing out the Hippo/YAP Signaling Pathway: Toward a New Role in Schwann Cells. Cell Death Dis. 2015, 6, e1915. [Google Scholar] [CrossRef]
- Mindos, T.; Dun, X.; North, K.; Doddrell, R.D.S.; Schulz, A.; Edwards, P.; Russell, J.; Gray, B.; Roberts, S.L.; Shivane, A.; et al. Merlin Controls the Repair Capacity of Schwann Cells after Injury by Regulating Hippo/YAP Activity. J. Cell Biol. 2017, 216, 495–510. [Google Scholar] [CrossRef]
- Salah, N.; Eissa, S.; Mansour, A.; El Magd, N.M.A.; Hasanin, A.H.; El Mahdy, M.M.; Hassan, M.K.; Matboli, M. Evaluation of the Role of Kefir in Management of Non-Alcoholic Steatohepatitis Rat Model via Modulation of NASH Linked MRNA-MiRNA Panel. Sci. Rep. 2023, 13, 236. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Fernández, R.; Martín-Ramírez, R.; Maeso, M.-d.-C.; Lázaro, A.; Ávila, J.; Martín-Vasallo, P.; Morales, M. Changes in AmotL2 Expression in Cells of the Human Enteral Nervous System in Oxaliplatin-Induced Enteric Neuropathy. Biomedicines 2024, 12, 1952. https://doi.org/10.3390/biomedicines12091952
González-Fernández R, Martín-Ramírez R, Maeso M-d-C, Lázaro A, Ávila J, Martín-Vasallo P, Morales M. Changes in AmotL2 Expression in Cells of the Human Enteral Nervous System in Oxaliplatin-Induced Enteric Neuropathy. Biomedicines. 2024; 12(9):1952. https://doi.org/10.3390/biomedicines12091952
Chicago/Turabian StyleGonzález-Fernández, Rebeca, Rita Martín-Ramírez, María-del-Carmen Maeso, Alberto Lázaro, Julio Ávila, Pablo Martín-Vasallo, and Manuel Morales. 2024. "Changes in AmotL2 Expression in Cells of the Human Enteral Nervous System in Oxaliplatin-Induced Enteric Neuropathy" Biomedicines 12, no. 9: 1952. https://doi.org/10.3390/biomedicines12091952
APA StyleGonzález-Fernández, R., Martín-Ramírez, R., Maeso, M.-d.-C., Lázaro, A., Ávila, J., Martín-Vasallo, P., & Morales, M. (2024). Changes in AmotL2 Expression in Cells of the Human Enteral Nervous System in Oxaliplatin-Induced Enteric Neuropathy. Biomedicines, 12(9), 1952. https://doi.org/10.3390/biomedicines12091952