

Astilbin Alleviates Radiation-Induced Pulmonary Fibrosis via circPRKCE Targeting the TGF-β/Smad7 Pathway to Inhibit Epithelial–Mesenchymal Transition


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Irradiation
2.2. Animals and Irradiation
2.3. AST Treatment
2.4. Western Blotting (WB)
2.5. Real-Time RT-PCR
2.6. Cell Counting Kit 8 Assays (CCK-8)
2.7. Histopathology
2.8. Immunohistochemistry
2.9. Bioinformatics Analysis
2.10. Sanger Sequencing, RNase R Digestion, and DNA Gel Electrophoresis
2.11. Fluorescent In Situ Hybridization (FISH)
2.12. RNA Immunoprecipitation (RIP) Assay
2.13. Dual-Luciferase Reporter Assay
2.14. Construction of Stable-Infected Cell Lines
2.15. Flow Cytometry
2.16. Statistical Analysis Doses and Times
3. Results
3.1. AST Reduces Radiation-Induced Inhibition of Proliferation in Mouse-Derived Lung Epithelial Cells In Vitro
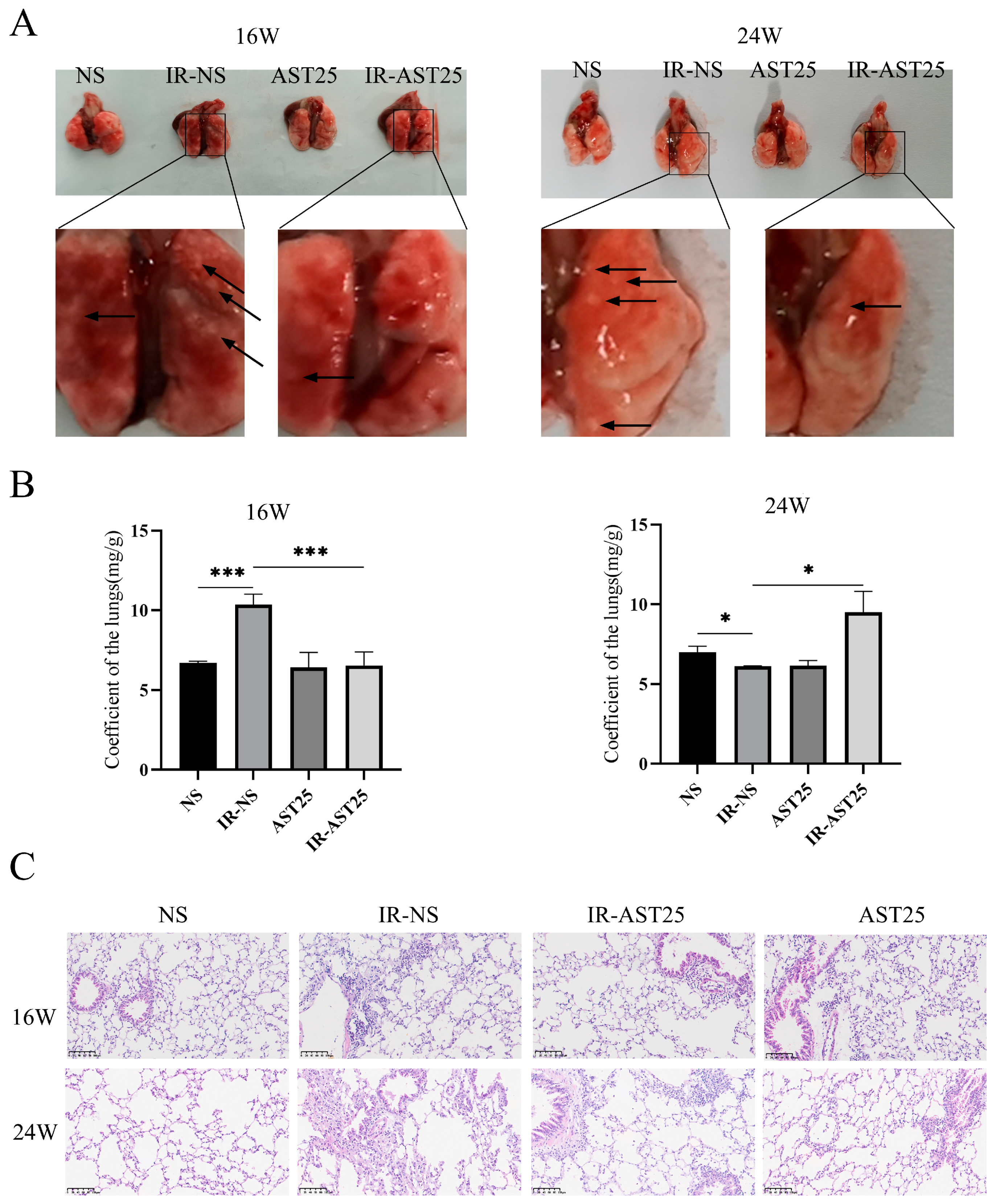
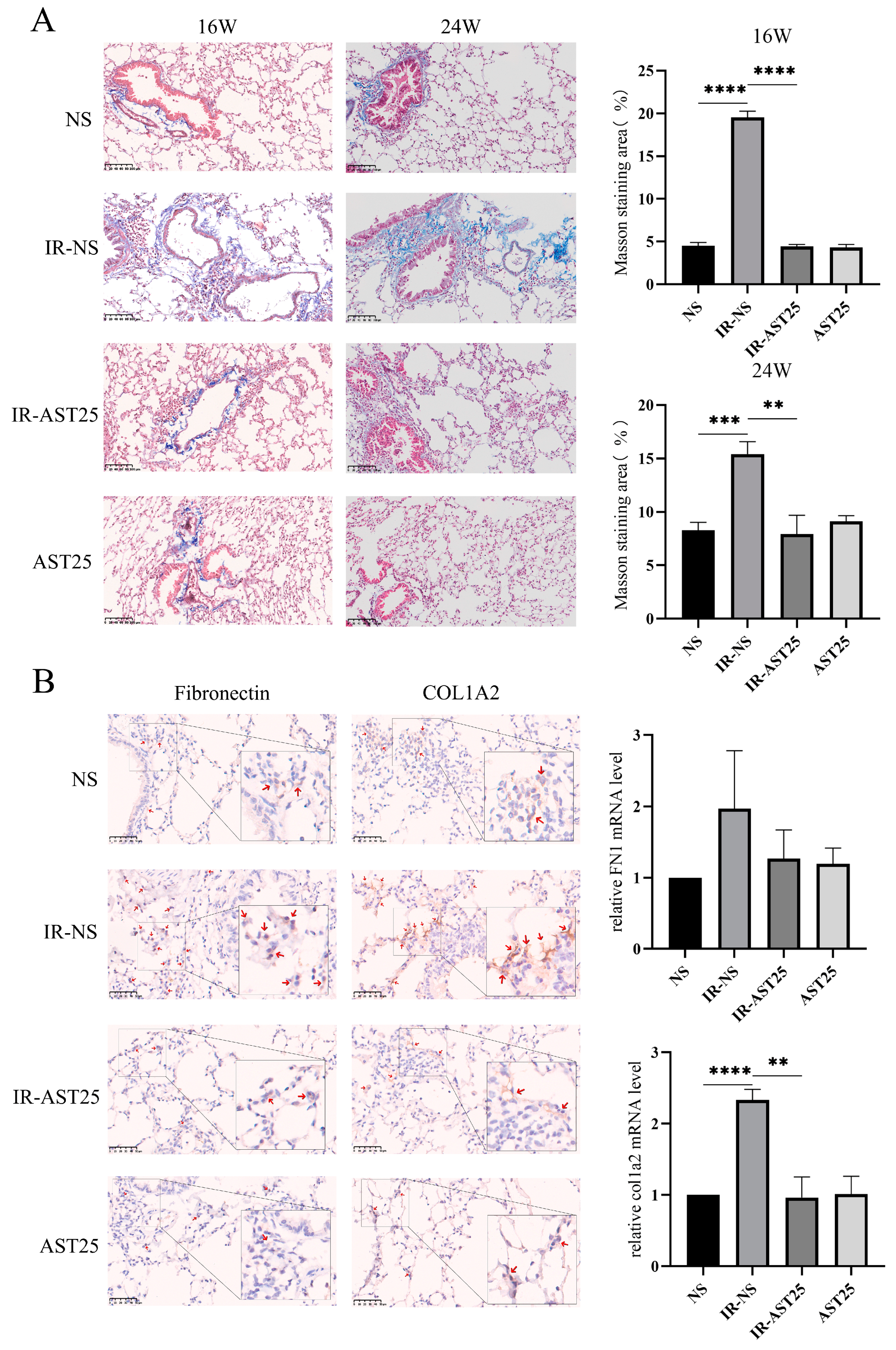
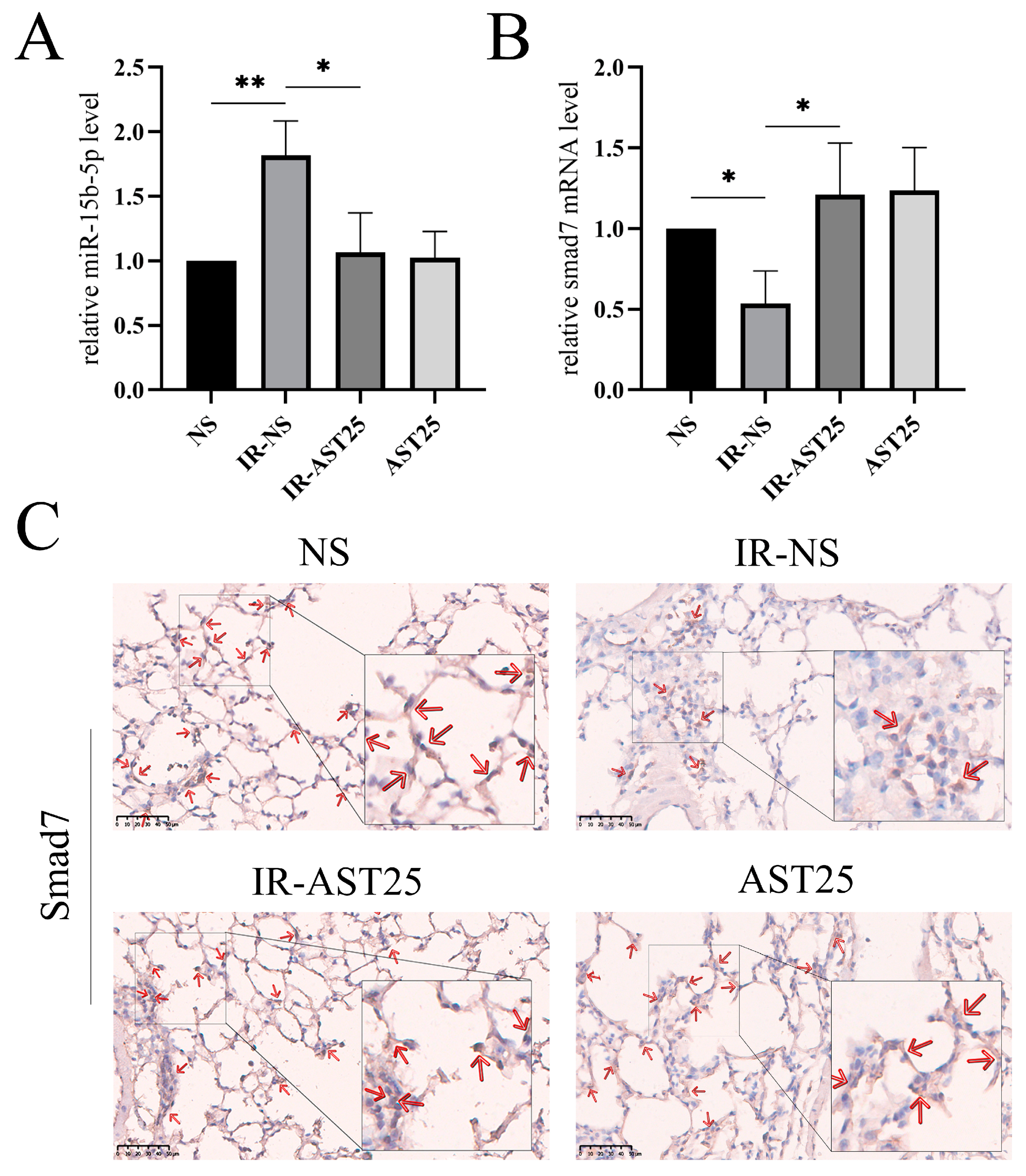
3.2. AST Alleviates Radiation-Induced Pulmonary Fibrosis In Vivo
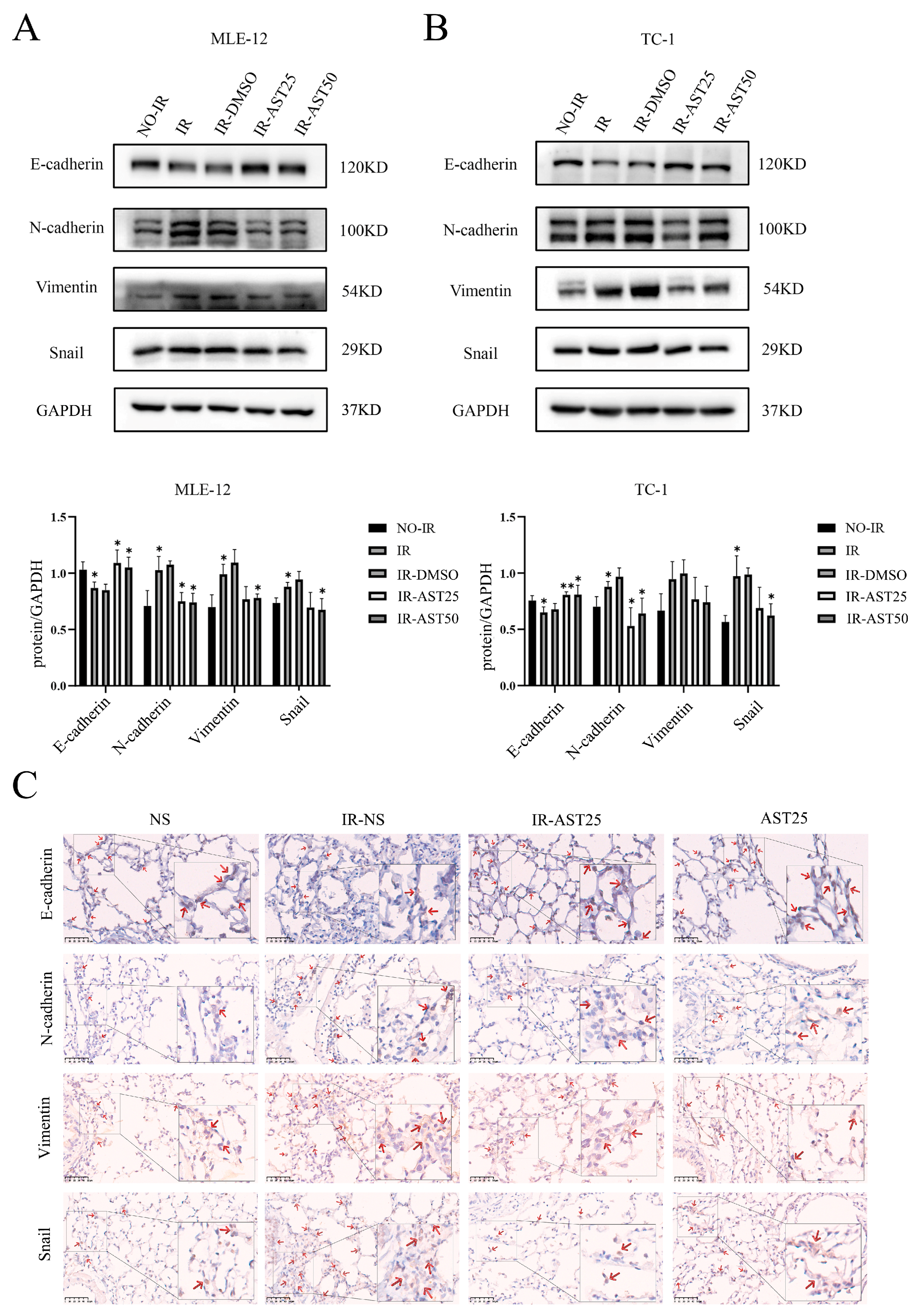
3.3. AST Inhibits Radiation-Mediated Epithelial-to-Mesenchymal Transition Progression
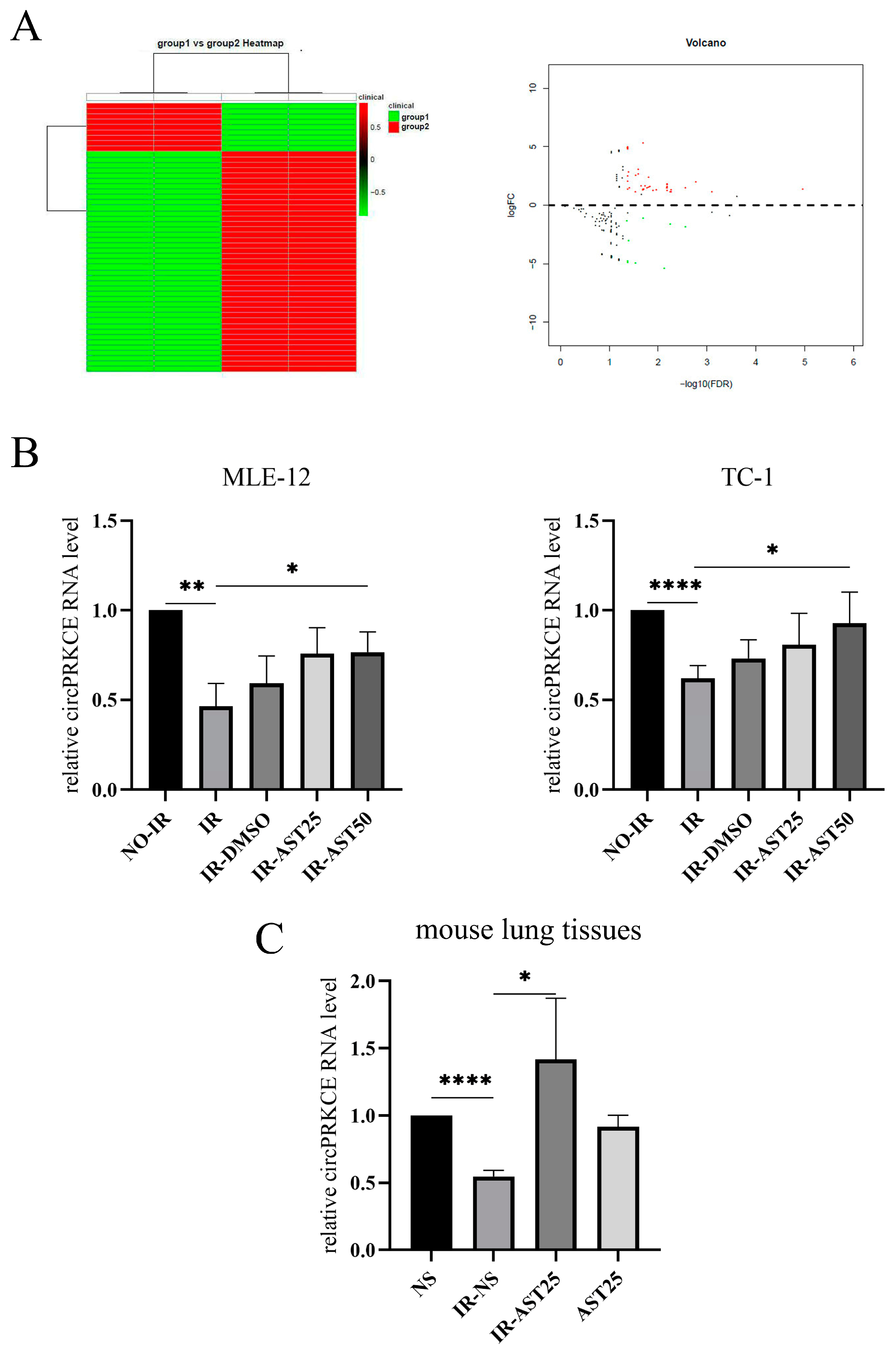
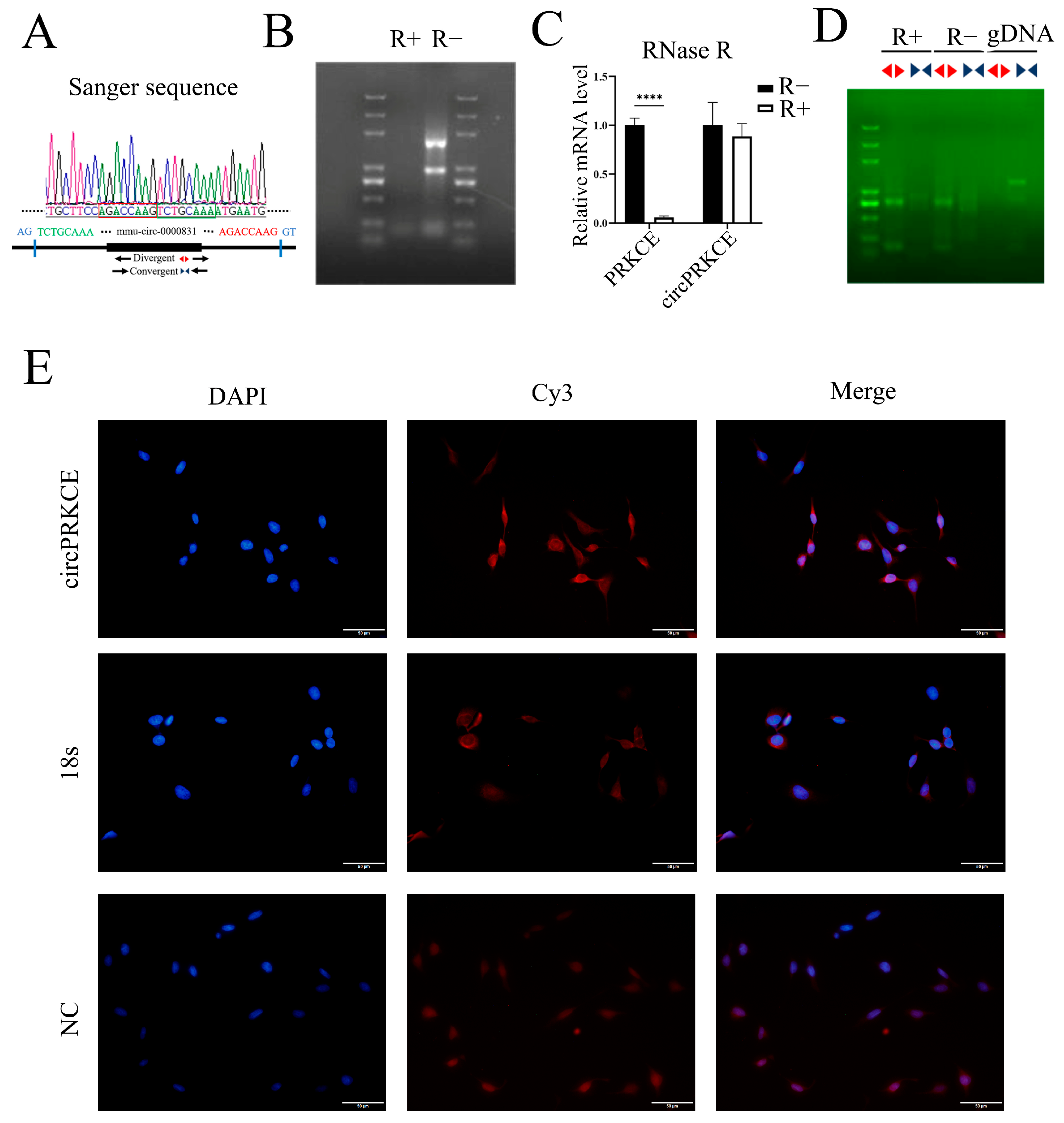
3.4. AST Inhibits Radiation-Induced Pulmonary Fibrosis by Enhancing circPRKCE Expression
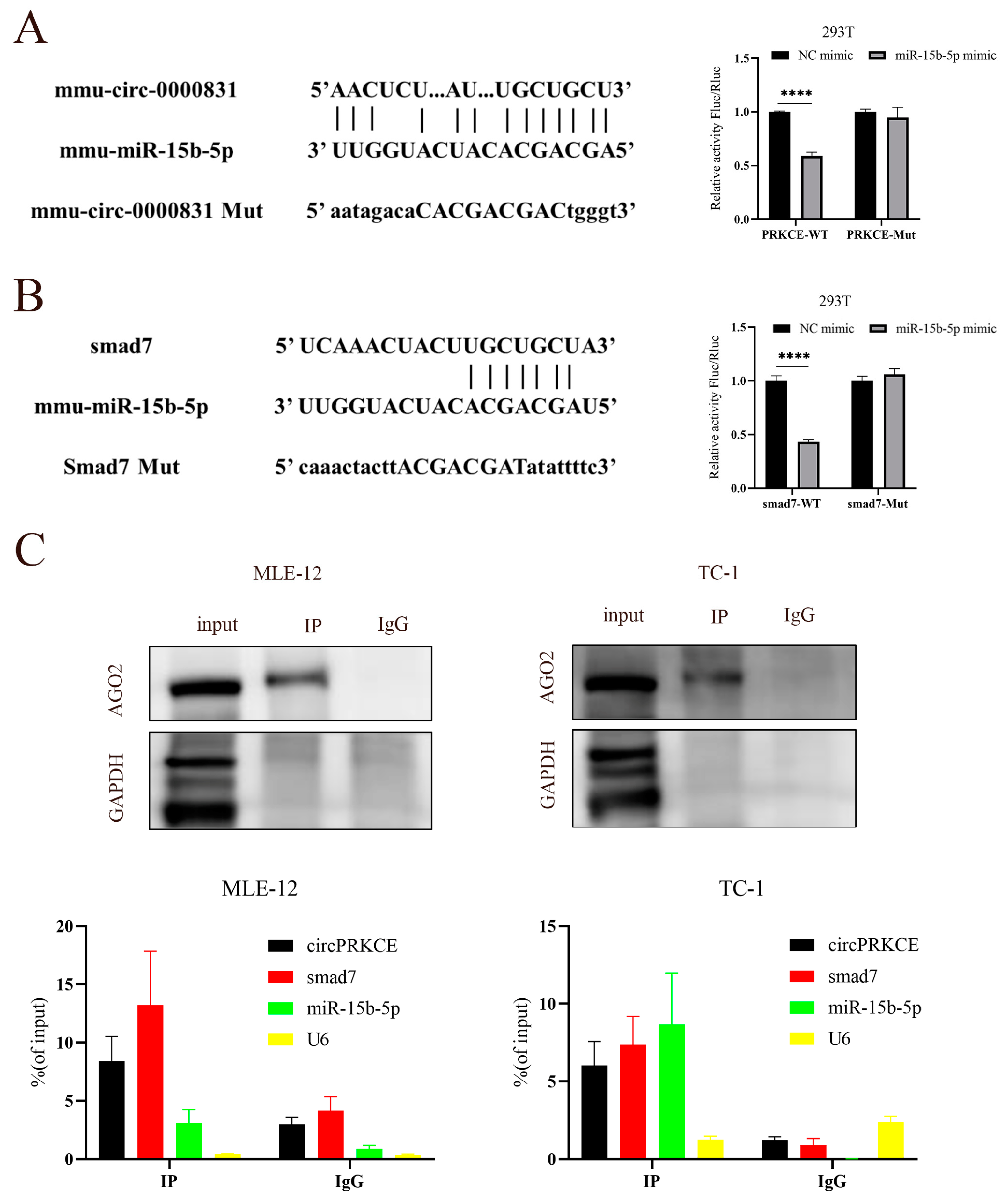
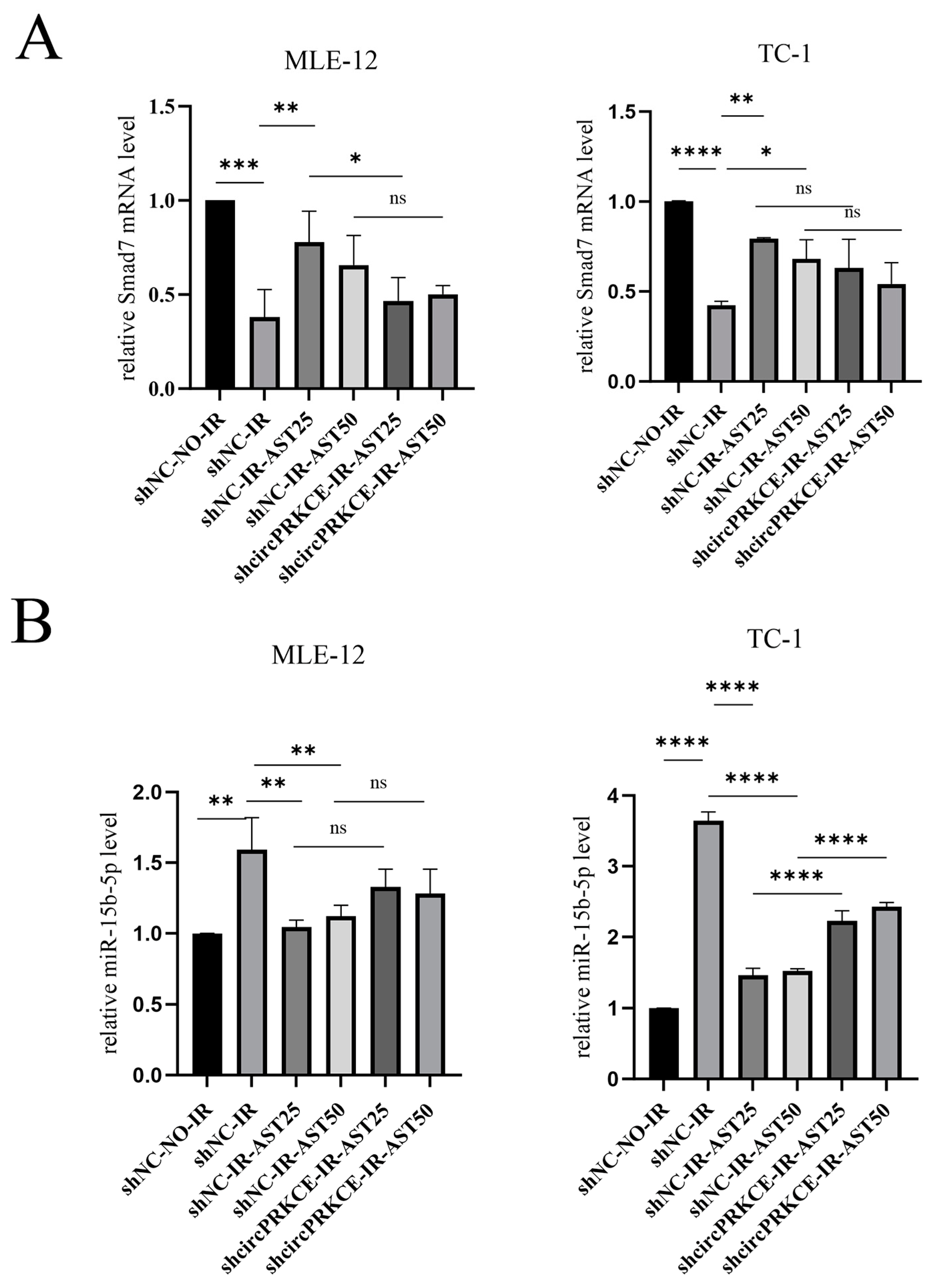
3.5. AST Inhibited Radiation Induced Epithelial-to-Mesenchymal Transition by Regulating circPEKCE/miR-15b-5p/smad7 Axis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Vinod, S.K.; Hau, E. Radiotherapy treatment for lung cancer: Current status and future directions. Respirology 2020, 25, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Bensenane, R.; Helfre, S.; Cao, K.; Carton, M.; Champion, L.; Girard, N.; Glorion, M.; Vieira, T.; Waissi, W.; Crehange, G.; et al. Optimizing lung cancer radiation therapy: A systematic review of multifactorial risk assessment for radiation-induced lung toxicity. Cancer Treat. Rev. 2024, 124, 102684. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef]
- Das, V.; Bhattacharya, S.; Chikkaputtaiah; Channakeshavaiah; Hazra, S.; Pal, M. The basics of epithelial–mesenchymal transition (EMT): A study from a structure, dynamics, and functional perspective. J. Cell. Physiol. 2019, 234, 14535–14555. [Google Scholar] [CrossRef]
- Marconi, G.D.; Fonticoli, L.; Rajan, T.S.; Pierdomenico, S.D.; Trubiani, O.; Pizzicannella, J.; Diomede, F. Epithelial-Mesenchymal Transition (EMT): The Type-2 EMT in Wound Healing, Tissue Regeneration and Organ Fibrosis. Cells 2021, 10, 1587. [Google Scholar] [CrossRef]
- Reilkoff, R.A.; Bucala, R.; Herzog, E.L. Fibrocytes: Emerging effector cells in chronic inflammation. Nat. Rev. Immunol. 2011, 11, 427–435. [Google Scholar] [CrossRef]
- Andarawewa, K.L.; Erickson, A.C.; Chou, W.S.; Costes, S.V.; Gascard, P.; Mott, J.D.; Bissell, M.J.; Barcellos-Hoff, M.H. Ionizing Radiation Predisposes Nonmalignant Human Mammary Epithelial Cells to Undergo Transforming Growth Factor β–Induced Epithelial to Mesenchymal Transition. Cancer Res. 2007, 67, 8662–8670. [Google Scholar] [CrossRef]
- Lu, J.; Zhong, Y.; Chen, J.; Lin, X.; Lin, Z.; Wang, N.; Lin, S. Radiation Enhances the Epithelial– Mesenchymal Transition of A549 Cells via miR3591-5p/USP33/PPM1A. Cell. Physiol. Biochem. 2018, 50, 721–733. [Google Scholar] [CrossRef]
- Xu, M.; Xie, F.; Tang, X.; Wang, T.; Wang, S. Insights into the role of circular RNA in macrophage activation and fibrosis disease. Pharmacol. Res. 2020, 156, 104777. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Abak, A.; Talebi, S.F.; Shoorei, H.; Branicki, W.; Taheri, M.; Akbari Dilmaghani, N. Role of miRNA and lncRNAs in organ fibrosis and aging. Biomed. Pharmacother. 2021, 143, 112132. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Li, Y.; Han, L.; Ji, X.; Pan, H.; Liu, Y.; Yuan, J.; Yan, W.; Ni, C. The CDR1as/miR-7/TGFBR2 Axis Modulates EMT in Silica-Induced Pulmonary Fibrosis. Toxicol. Sci. Off. J. Soc. Toxicol. 2018, 166, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-x.; Lu, J.; Xie, H.; Wang, D.-p.; Ni, H.-e.; Zhu, Y.; Ren, L.-h.; Meng, X.-x.; Wang, R.-l. circHIPK3 regulates lung fibroblast-to-myofibroblast transition by functioning as a competing endogenous RNA. Cell Death Dis. 2019, 10, 182. [Google Scholar] [CrossRef]
- Zhou, Y.; Kwon, O.-S.; Kim, K.-T.; Lee, E.; Kim, M.; Choi, S.-H.; Li, H.; Fornace, A.J.; Cho, J.-H.; Lee, Y.-S.; et al. Induction of MiR-21 by Stereotactic Body Radiotherapy Contributes to the Pulmonary Fibrotic Response. PLoS ONE 2016, 11, e0154942. [Google Scholar] [CrossRef]
- Ibáñez, B.; Melero, A.; Montoro, A.; Merino-Torres, J.F.; Soriano, J.M.; San Onofre, N. A Narrative Review of the Herbal Preparation of Ayurvedic, Traditional Chinese, and Kampō Medicines Applied as Radioprotectors. Antioxidants 2023, 12, 1437. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Hu, K.; Jiang, J.; Chen, Y.; Liu, W.; Wang, R. Sarcandra glabra inhibits the epithelial-mesenchymal transition in radiation-induced lung injury in minipigs. World Sci. Technol.-Mod. Tradit. Chin. Med. 2016, 18, 846–853. [Google Scholar]
- Sun, S.; Yan, Z.; Shui, X.; Qi, W.; Chen, Y.; Xu, X.; Hu, Y.; Guo, W.; Shang, P. Astilbin prevents osteoarthritis development through the TLR4/MD-2 pathway. J. Cell. Mol. Med. 2020, 24, 13104–13114. [Google Scholar] [CrossRef]
- Han, S.; Lin, Z.; Wen, J.; Wu, K.; Xu, Y.; Zhang, Y.; Lu, G.; Xiao, W.; Ding, Y.; Jia, X.; et al. Astilbin promotes the induction of regulatory NK1.1− CD4+ NKG2D+ T cells through the PI3K, STAT3, and MAPK signaling pathways. Int. Immunopharmacol. 2020, 81, 106143. [Google Scholar] [CrossRef]
- Ding, S.; Lu, G.; Wang, B.; Xiang, J.; Hu, C.; Lin, Z.; Ding, Y.; Xiao, W.; Gong, W. Astilbin Activates the Reactive Oxidative Species/PPARγ Pathway to Suppress Effector CD4+ T Cell Activities via Direct Binding with Cytochrome P450 1B1. Front. Pharmacol. 2022, 13, 848957. [Google Scholar] [CrossRef]
- Fang, Z.; Wang, G.; Huang, R.; Liu, C.; Yushanjiang, F.; Mao, T.; Li, J. Astilbin protects from sepsis-induced cardiac injury through the NRF2/HO-1 and TLR4/NF-κB pathway. Phytother. Res. 2023, 38, 1044–1058. [Google Scholar] [CrossRef]
- Wang, T.; Ye, Y.; Ji, J.; Zhang, S.; Yang, X.; Xu, J.; Wang, J.-S.; Chen, Z.; Xia, B.; Shen, H.; et al. Astilbin fromSmilax glabraRoxb. alleviates high-fat diet-induced metabolic dysfunction. Food Funct. 2022, 13, 5023–5036. [Google Scholar] [CrossRef]
- Lu, G.; Zhang, J.; Liu, X.; Liu, W.; Cao, G.; Lv, C.; Zhang, X.; Xu, P.; Li, M.; Song, X. Regulatory network of two circRNAs and an miRNA with their targeted genes under astilbin treatment in pulmonary fibrosis. J. Cell. Mol. Med. 2019, 23, 6720–6729. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, H.; Song, C.; Zhang, J.; Wang, Y.; Lv, C.; Song, X. Astilbin ameliorates pulmonary fibrosis via blockade of Hedgehog signaling pathway. Pulm. Pharmacol. Ther. 2018, 50, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Sun, S.; Zhao, J.; Wu, Q.; He, W.; Sun, W. Yishen-Qingli-Huoxue formula attenuates renal fibrosis by inhibiting indoxyl sulfate via AhR/snai1 signaling. Phytomedicine 2023, 108, 154546. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.-H.; Zhang, H.; Fan, X.-P.; Wang, Z.-H. Astilbin Protects Against Carbon Tetrachloride-Induced Liver Fibrosis in Rats. Pharmacology 2021, 106, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Huang, Y.; Chen, L.; Shi, Z.; Wang, H.; Zhang, T.; Li, Z.; Mi, J.; Fan, T.; Lu, Y.; et al. Radioprotective efficacy of Astilbin in mitigating radiation-induced lung injury through inhibition of p53 acetylation. Environ. Toxicol. 2023, 38, 2967–2980. [Google Scholar] [CrossRef]
- Zhang, T.; Mi, J.; Qin, X.; Ouyang, Z.; Wang, Y.; Li, Z.; He, S.; Hu, K.; Wang, R.; Huang, W. Rosmarinic Acid Alleviates Radiation-Induced Pulmonary Fibrosis by Downregulating the tRNA N7-Methylguanosine Modification-Regulated Fibroblast-to-Myofibroblast Transition Through the Exosome Pathway. J. Inflamm. Res. 2024, 17, 5567–5586. [Google Scholar] [CrossRef]
- Yang, Y.; Gao, X.; Zhang, M.; Yan, S.; Sun, C.; Xiao, F.; Huang, N.; Yang, X.; Zhao, K.; Zhou, H.; et al. Novel Role of FBXW7 Circular RNA in Repressing Glioma Tumorigenesis. JNCI J. Natl. Cancer Inst. 2018, 110, 304–315. [Google Scholar] [CrossRef]
- Liu, Y.; Shi, M.; He, X.; Cao, Y.; Liu, P.; Li, F.; Zou, S.; Wen, C.; Zhan, Q.; Xu, Z.; et al. LncRNA-PACERR induces pro-tumour macrophages via interacting with miR-671-3p and m6A-reader IGF2BP2 in pancreatic ductal adenocarcinoma. J. Hematol. Oncol. 2022, 15, 52. [Google Scholar] [CrossRef]
- Yan, Y.; Fu, J.; Kowalchuk, R.O.; Wright, C.M.; Zhang, R.; Li, X.; Xu, Y. Exploration of radiation-induced lung injury, from mechanism to treatment: A narrative review. Transl. Lung Cancer Res. 2022, 11, 307–322. [Google Scholar] [CrossRef]
- Xiong, S.; Pan, X.; Xu, L.; Yang, Z.; Guo, R.; Gu, Y.; Li, R.; Wang, Q.; Xiao, F.; Du, L.; et al. Regulatory T Cells Promote β-Catenin–Mediated Epithelium-to-Mesenchyme Transition During Radiation-Induced Pulmonary Fibrosis. Int. J. Radiat. Oncol. *Biol. *Phys. 2015, 93, 425–435. [Google Scholar] [CrossRef]
- Tofilon, P.J.; Almeida, C.; Nagarajan, D.; Tian, J.; Leal, S.W.; Wheeler, K.; Munley, M.; Blackstock, W.; Zhao, W. The Role of Alveolar Epithelium in Radiation-Induced Lung Injury. PLoS ONE 2013, 8, e53628. [Google Scholar] [CrossRef]
- Kartha, R.V.; Subramanian, S. Competing endogenous RNAs (ceRNAs): New entrants to the intricacies of gene regulation. Front. Genet. 2014, 5, 8. [Google Scholar] [CrossRef] [PubMed]
- Peng, D.; Fu, M.; Wang, M.; Wei, Y.; Wei, X. Targeting TGF-β signal transduction for fibrosis and cancer therapy. Mol. Cancer 2022, 21, 104. [Google Scholar] [CrossRef] [PubMed]
- Hanania, A.N.; Mainwaring, W.; Ghebre, Y.T.; Hanania, N.A.; Ludwig, M. Radiation-Induced Lung Injury. Chest 2019, 156, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-y.; Zhu, J.-y.; Ye, Y.; Zhang, M.; Zhang, L.-j.; Wang, S.-j.; Song, Y.-n.; Zhang, H. Erhuang Formula ameliorates renal damage in adenine–induced chronic renal failure rats via inhibiting inflammatory and fibrotic responses. Biomed. Pharmacother. 2017, 95, 520–528. [Google Scholar] [CrossRef]
- Li, Y.; Shen, Z.; Jiang, X.; Wang, Y.; Yang, Z.; Mao, Y.; Wu, Z.; Li, G.; Chen, H. Mouse mesenchymal stem cell-derived exosomal miR-466f-3p reverses EMT process through inhibiting AKT/GSK3β pathway via c-MET in radiation-induced lung injury. J. Exp. Clin. Cancer Res. 2022, 41, 128. [Google Scholar] [CrossRef]
- Yang, C.; Song, C.; Wang, Y.; Zhou, W.; Zheng, W.; Zhou, H.; Deng, G.; Li, H.; Xiao, W.; Yang, Z.; et al. Re-Du-Ning injection ameliorates radiation-induced pneumonitis and fibrosis by inhibiting AIM2 inflammasome and epithelial-mesenchymal transition. Phytomedicine 2022, 102, 154184. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Ao, X.; Liang, X.; Chen, Z.; Liu, Y.; Wang, P.; Wang, D.; Liu, Z.; Liu, X.; Zhu, J.; et al. Transcriptional inhibition of miR-486-3p by BCL6 upregulates Snail and induces epithelial–mesenchymal transition during radiation-induced pulmonary fibrosis. Respir. Res. 2022, 23, 104. [Google Scholar] [CrossRef]
- Park, H.-R.; Jo, S.-K.; Jung, U. Ionizing Radiation Promotes Epithelial–to–Mesenchymal Transition in Lung Epithelial Cells by TGF-β-producing M2 Macrophages. Vivo 2019, 33, 1773–1784. [Google Scholar] [CrossRef]
- Ying, H.; Fang, M.; Hang, Q.; Chen, Y.; Qian, X.; Chen, M. Pirfenidone modulates macrophage polarization and ameliorates radiation-induced lung fibrosis by inhibiting the TGF-β1/Smad3 pathway. J. Cell. Mol. Med. 2021, 25, 8662–8675. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-H.; Kim, J.-Y.; Kim, J.-M.; Yoo, B.R.; Han, S.Y.; Jung, Y.J.; Bae, H.; Cho, J. PM014 attenuates radiation-induced pulmonary fibrosis via regulating NF-kB and TGF-b1/NOX4 pathways. Sci. Rep. 2020, 10, 16112. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Jian, X.; Zhang, Z.; Ning, Q.; Kan, B.; Kong, L. Effects of tacrolimus on the TGF-β1/SMAD signaling pathway in paraquat-exposed rat alveolar type II epithelial cells. Mol. Med. Rep. 2020, 22, 3687–3694. [Google Scholar] [CrossRef]
- Gu, X.; Jiang, Y.-N.; Wang, W.-J.; Zhang, J.; Shang, D.-S.; Sun, C.-B.; Tian, J.-T.; Tian, J.-W.; Yu, B.; Zhang, Y. Comprehensive circRNA expression profile and construction of circRNA-related ceRNA network in cardiac fibrosis. Biomed. Pharmacother. 2020, 125, 109944. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Wei, J.; Zeng, Y.; Liu, J.; Xiao, E.; Kang, Y.; Kang, Y. Mesenchymal stem cell-originated exosomal circDIDO1 suppresses hepatic stellate cell activation by miR-141-3p/PTEN/AKT pathway in human liver fibrosis. Drug Deliv. 2022, 29, 440–453. [Google Scholar] [CrossRef]
- Peng, F.; Gong, W.; Li, S.; Yin, B.; Zhao, C.; Liu, W.; Chen, X.; Luo, C.; Huang, Q.; Chen, T.; et al. circRNA_010383 Acts as a Sponge for miR-135a, and Its Downregulated Expression Contributes to Renal Fibrosis in Diabetic Nephropathy. Diabetes 2021, 70, 603–615. [Google Scholar] [CrossRef]
- Xu, Q.; Cheng, D.; Li, G.; Liu, Y.; Li, P.; Sun, W.; Ma, D.; Ni, C. CircHIPK3 regulates pulmonary fibrosis by facilitating glycolysis in miR-30a-3p/FOXK2-dependent manner. Int. J. Biol. Sci. 2021, 17, 2294–2307. [Google Scholar] [CrossRef]
- Jiang, X.-P.; Ai, W.-B.; Wan, L.-Y.; Zhang, Y.-Q.; Wu, J.-F. The roles of microRNA families in hepatic fibrosis. Cell Biosci. 2017, 7, 34. [Google Scholar] [CrossRef]
- Rawal, S.; Munasinghe, P.E.; Nagesh, P.T.; Lew, J.K.S.; Jones, G.T.; Williams, M.J.A.; Davis, P.; Bunton, D.; Galvin, I.F.; Manning, P.; et al. Down-regulation of miR-15a/b accelerates fibrotic remodelling in the Type 2 diabetic human and mouse heart. Clin. Sci. 2017, 131, 847–863. [Google Scholar] [CrossRef]
- Tijsen, A.J.; van der Made, I.; van den Hoogenhof, M.M.; Wijnen, W.J.; van Deel, E.D.; de Groot, N.E.; Alekseev, S.; Fluiter, K.; Schroen, B.; Goumans, M.-J.; et al. The microRNA-15 family inhibits the TGFβ-pathway in the heart. Cardiovasc. Res. 2014, 104, 61–71. [Google Scholar] [CrossRef]
- Prasanna, P.G.S.; Aryankalayil, M.; Citrin, D.E.; Coleman, C.N. Radiation-induced pulmonary fibrosis: Roles of therapy-induced senescence and microRNAs. Int. J. Radiat. Biol. 2023, 99, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Zhang, Q.-F. The natural source, physicochemical properties, biological activities and metabolism of astilbin. Crit. Rev. Food Sci. Nutr. 2022, 63, 9506–9518. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Z.; Liu, J.; Qin, J.; Liang, X.; Ou, X.; Zhang, T.; Yan, X.; Hu, Q.; Huang, W.; Hu, K. Astilbin Alleviates Radiation-Induced Pulmonary Fibrosis via circPRKCE Targeting the TGF-β/Smad7 Pathway to Inhibit Epithelial–Mesenchymal Transition. Biomedicines 2025, 13, 689. https://doi.org/10.3390/biomedicines13030689
Shi Z, Liu J, Qin J, Liang X, Ou X, Zhang T, Yan X, Hu Q, Huang W, Hu K. Astilbin Alleviates Radiation-Induced Pulmonary Fibrosis via circPRKCE Targeting the TGF-β/Smad7 Pathway to Inhibit Epithelial–Mesenchymal Transition. Biomedicines. 2025; 13(3):689. https://doi.org/10.3390/biomedicines13030689
Chicago/Turabian StyleShi, Zhiling, Jing Liu, Jing Qin, Xian Liang, Xue Ou, Tingting Zhang, Xueting Yan, Qianxin Hu, Weimei Huang, and Kai Hu. 2025. "Astilbin Alleviates Radiation-Induced Pulmonary Fibrosis via circPRKCE Targeting the TGF-β/Smad7 Pathway to Inhibit Epithelial–Mesenchymal Transition" Biomedicines 13, no. 3: 689. https://doi.org/10.3390/biomedicines13030689
APA StyleShi, Z., Liu, J., Qin, J., Liang, X., Ou, X., Zhang, T., Yan, X., Hu, Q., Huang, W., & Hu, K. (2025). Astilbin Alleviates Radiation-Induced Pulmonary Fibrosis via circPRKCE Targeting the TGF-β/Smad7 Pathway to Inhibit Epithelial–Mesenchymal Transition. Biomedicines, 13(3), 689. https://doi.org/10.3390/biomedicines13030689