
The Role of miRNAs as Early Biomarkers in Obesity-Related Glomerulopathy: Implications for Early Detection and Treatment


Abstract
:1. Introduction
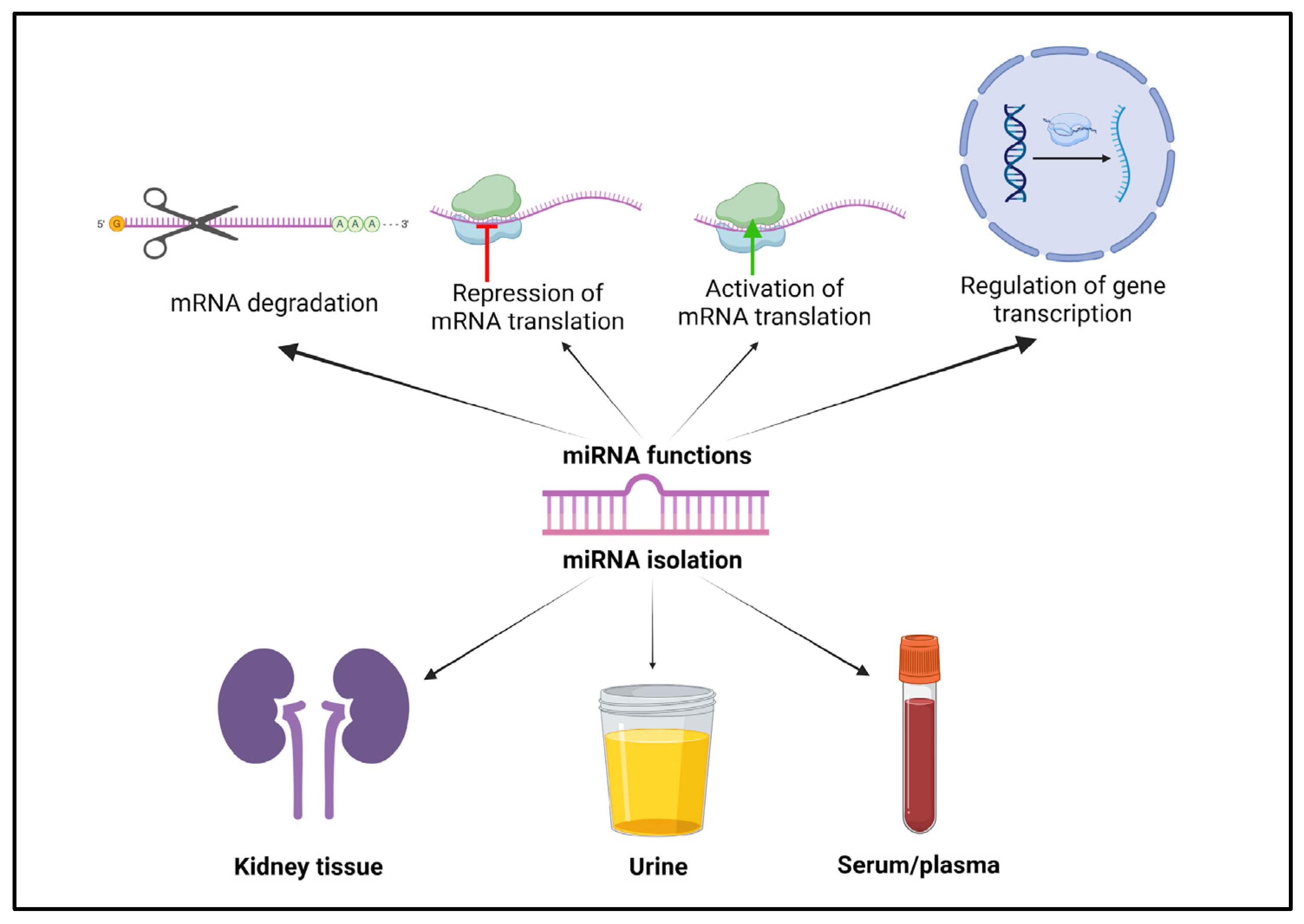
2. MicroRNAs: Biological Functions and Roles in Obesity-Associated Pathophysiology
3. An Overview of miRNA Changes in Different Diseases, with an Emphasis on Renal Pathologies
4. The Emerging Role of miRNAs in Obesity-Related Glomerulopathy
- -
- miR-802 [52]: The IkB kinase/NF-kB (IKK/NF-kB) signaling pathway is involved in the synthesis and secretion of chemokines. The miR-802 has previously been related with pro-inflammatory characteristics. As inflammation is one of the main cornerstones of obesity-related kidney disease development, Sun D et al. aimed to evaluate the implications of miR-802 in ORG. After 16 weeks of feeding C57BL/6J mice with normal chow (NC) or a high-fat diet (HFD), the HFD group showed significantly increased miR-802 levels in the kidneys. The increase in miR-802 was also positively related with serum levels of BUN and creatinine. Moreover, the authors demonstrated that miR-802 activated IKK/NF-kB pathways through interaction with the 3′ UTR of NF-Kn-repressing factor. Another group of obese rodents was treated with 1 × 109 lentivirus particles encoding either a miR-802 sponge, which would bind and neutralize miR-802, or a control, to investigate the effects of miR-802 inhibition. Treatment with the miR-802 sponge led to a significant reduction in weight, blood urea nitrogen (BUN), inflammatory markers, fibrosis factors, glomerular size and basement membrane thickness. Finally, the circulating levels of miR-802 were higher in subjects with obesity compared with lean individuals. Although miR-802 was positively correlated with creatinine levels in humans, it was negatively correlated with creatinine clearance.
- -
- miR-130b [53]: Adiponectin is an adipokine with anti-inflammatory properties that is usually decreased in patients with obesity. These low levels of adiponectin are thought to encourage overactivation of the renal sympathetic nervous system and also promote albuminuria through podocyte injury [4]. miR-130b has previously been related to renal fibrosis and other kidney diseases. After 16 weeks of a control diet or an HFD in adipoKO and wildtype (WT) mice with a C57BL/6J background, Pereira BMV et al. confirmed that there was overexpression of miR-130b in the kidney of the group of adipoKO mice with an HFD. Both adipoKO and WT mice fed with an HFD showed non-detectable levels of adiponectin compared with groups. However, an in vitro analysis of cultured podocytes of WT mice disproved the adiponectin treatment’s regulation of miR-130 expression [53].
- -
- miR-21 [54]: In order to study the role of inflammation, fibrosis and adiponectin in obesity-related kidney disease, Morrison MC et al. fed a human CRP transgenic (huCRPtg) mouse model with a C57BL/6J background with an NC diet or an HFD for 50 weeks, and also evaluated the effects of rosiglitazone and rosuvastatin based on their anti-inflammatory effects. Since miR-21 has previously been associated with fibrosis, inflammation and albuminuria, its expression was also measured. Renal miR-21 was overexpressed in the HFD group compared with the NC group. Also, miR-21 expression was significantly correlated with the kidney damage marker KIM-1 and with renal fibrosis, while no such correlations were found for adiponectin levels or exposure.
- -
- miR-155 [55]: Circulating miR-155 has already been associated with eGFR and proteinuria in CKD, and with histological kidney damage in patients with IgA nephropathy. Zheng C et al. fed C57BL/6J mice with either an HFD or a control diet (CD) for 20 weeks. Afterwards, the HFD mice were injected with 1 × 1010 IU viral particles of lentivirus encoding the miR-155 sponge or a control vector. To silence SHIP1/INNP5D, which is a negative regulator of inflammatory response in the NF-κB signaling pathway, 1 × 1010 IU viral particles of lentivirus encoding INPP5D siRNA were also injected. Four weeks after these infusions, renal miR-155 was upregulated 4-fold in HFD mice in comparison to CD mice. There was a linear correlation between renal miR-155 and albuminuria and BUN. Those HFD mice treated with the miR-155 sponge showed attenuation of glomerular enlargement, fibrosis and tubular damage, blocked macrophage infiltration and lipid deposits, and decreased albuminuria and BUN, compared with the control vector-treated HFD mice. The authors were able to confirm that miR-155 significantly decreased the luciferase activity in INPP5D 3′-UTR transfected cells, and real-time PCR analysis showed that the gene level of INPP5D was significantly decreased by overexpression of miR-155 in the kidneys. Finally, the suppression of INPP5D increased the levels of albuminuria and BUN and significantly increased glomerular size; all of this was accompanied by renal inflammatory response and oxidative stress. In summary, miR-155 seems to contribute to ORG development by promoting inflammation and oxidative stress through inhibition of the INNP5D signaling pathway.
- -
- miR-205 [8]: The PTEN gene (phosphatase and tensin homolog) is essential in kidney health, particularly in relation to inflammation and podocyte injury, in which case it is regulated by various miRNAs [56,57]. Its inhibition has been associated with several obesity-related pathways, including enhanced muscle regeneration [58], mitochondrial dysfunction [59], endoplasmic reticulum stress [60], increased renal inflammation and fibrosis [61] and the promotion of insulin resistance [62]. Interestingly, podocytes in patients with ORG have shown increased lipid endocytosis, which has been linked to reduced PTEN expression [63]. In this model, Wistar Han rats were fed with a standard diet or an HFD for 10 weeks. It was proven that the downregulation of PTEN in the kidneys of the HFD group was driven by interactions between miR-205, miR-22-3p, miR-22-5p and miR-144-3p, and was associated with increased lipid uptake in podocytes (Figure 2). Notably, miR-205 was not only upregulated in the kidneys of rats with ORG, but was also found to be differentially expressed in their urine (Figure 2). Furthermore, the increase in the mesangial matrix and podocyte hypertrophy observed in these animals correlated with changes in miRNA expression in both kidney tissue and urine.
5. Future Perspectives and Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ORG | Obesity-related glomerulopathy |
| CKD | Chronic kidney disease |
| miRNAs | MicroRNAs |
| mRNA | Messenger RNA |
| BUN | Blood urea nitrogen |
| CD | Control diet |
| PTEN | Phosphatase and tensin homolog |
| SHIP1 | SH2-containing inositol phosphatase 1 |
| INNP5D | Inositol Polyphosphate-5-Phosphatase D |
| siRNA | Small interfering RNA |
References
- Weisinger, J.R.; Kempson, R.L.; Eldridge, F.L.; Swenson, R.S. The Nephrotic Syndrome: A Complication of Massive Obesity. Ann. Intern. Med. 1974, 81, 440–447. [Google Scholar] [CrossRef]
- Cohen, A.H. Massive obesity and the kidney. A morphologic and statistical study. Am. J. Pathol. 1975, 81, 117–130. [Google Scholar]
- Kambham, N.; Markowitz, G.S.; Valeri, A.M.; Lin, J.; D’agati, V.D. Obesity-related glomerulopathy: An emerging epidemic. Kidney Int. 2001, 59, 1498–1509. [Google Scholar] [CrossRef]
- D’Agati, V.D.; Chagnac, A.; De Vries, A.P.J.; Levi, M.; Porrini, E.; Herman-Edelstein, M.; Praga, M. Obesity-related glomerulopathy: Clinical and pathologic characteristics and pathogenesis. Nat. Rev. Nephrol. 2016, 12, 453–471. [Google Scholar] [CrossRef]
- Bikbov, B.; Purcell, C.A.; Levey, A.S.; Smith, M.; Abdoli, A.; Abebe, M.; Adebayo, O.M.; Afarideh, M.; Agarwal, S.K.; Agudelo-Botero, M.; et al. Global, regional, and national burden of chronic kidney disease, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2020, 395, 709–733. [Google Scholar] [CrossRef]
- Sowers, J.R. Obesity as a cardiovascular risk factor. Am. J. Med. 2003, 115 (Suppl. 8), 37S–41S. [Google Scholar] [CrossRef]
- Serra, A.; Romero, R.; Lopez, D.; Navarro, M.; Esteve, A.; Perez, N.; Alastrue, A.; Ariza, A. Renal injury in the extremely obese patients with normal renal function. Kidney Int. 2008, 73, 947–955. [Google Scholar] [CrossRef]
- López-Martínez, M.; Armengol, M.P.; Pey, I.; Farré, X.; Rodríguez-Martínez, P.; Ferrer, M.; Porrini, E.; Luis-Lima, S.; Díaz-Martín, L.; Rodríguez-Rodríguez, A.E.; et al. Integrated miRNA–mRNA Analysis Reveals Critical miRNAs and Targets in Diet-Induced Obesity-Related Glomerulopathy. Int. J. Mol. Sci. 2024, 25, 6437. [Google Scholar] [CrossRef]
- Jung, C.H.; Lee, M.J.; Kang, Y.M.; Hwang, J.Y.; Kim, E.H.; Park, J.-Y.; Kim, H.-K.; Lee, W.J. The risk of chronic kidney disease in a metabolically healthy obese population. Kidney Int. 2015, 88, 843–850. [Google Scholar] [CrossRef]
- Bancu, I.; Navarro Díaz, M.; Serra, A.; Granada, M.; Lopez, D.; Romero, R.; Bonet, J. Low Insulin-Like Growth Factor-1 Level in Obesity Nephropathy: A New Risk Factor? PLoS ONE 2016, 11, e0154451. [Google Scholar] [CrossRef] [PubMed]
- López-Martínez, M.; Luis-Lima, S.; Morales, E.; Navarro-Díaz, M.; Negrín-Mena, N.; Folgueras, T.; Escamilla, B.; Estupiñán, S.; Delgado-Mallén, P.; Marrero-Miranda, D.; et al. The estimation of GFR and the adjustment for BSA in overweight and obesity: A dreadful combination of two errors. Int. J. Obes. 2020, 44, 1129–1140. [Google Scholar] [CrossRef]
- Azushima, K.; Gurley, S.B.; Coffman, T.M. Modelling diabetic nephropathy in mice. Nat. Rev. Nephrol. 2018, 14, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Teixido-Trujillo, S.; Luis-Lima, S.; López-Martínez, M.; Navarro-Díaz, M.; Díaz-Martín, L.; Escasany-Martínez, E.; Gaspari, F.; Rodríguez-Rodríguez, A.E. Measured GFR in murine animal models: Review on methods, techniques, and procedures. Pflugers Arch. 2023, 475, 1241–1250. [Google Scholar] [CrossRef]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef]
- Cavaliere, A.F.; Perelli, F.; Zaami, S.; Piergentili, R.; Mattei, A.; Vizzielli, G.; Scambia, G.; Gianluca, S.; Restaino, S.; Signore, F. Towards Personalized Medicine: Non-Coding RNAs and Endometrial Cancer. Healthcare 2021, 9, 965. [Google Scholar] [CrossRef] [PubMed]
- Seyhan, A.A. Circulating microRNAs as Potential Biomarkers in Pancreatic Cancer—Advances and Challenges. Int. J. Mol. Sci. 2023, 24, 13340. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wu, C.; Yang, Q.; Ding, M.; Zhong, J.; Zhang, C.-Y.; Ge, J.; Wang, J.; Zhang, C. miR-28-5p acts as a tumor suppressor in renal cell carcinoma for multiple antitumor effects by targeting RAP1B. Oncotarget 2016, 7, 73888–73902. [Google Scholar] [CrossRef]
- Seyhan, A.A. Trials and Tribulations of MicroRNA Therapeutics. Int. J. Mol. Sci. 2024, 25, 1469. [Google Scholar] [CrossRef]
- Caus, M.; Eritja, À.; Bozic, M. Role of micrornas in obesity-related kidney disease. Int. J. Mol. Sci. 2021, 22, 11416. [Google Scholar] [CrossRef]
- Mahtal, N.; Lenoir, O.; Tinel, C.; Anglicheau, D.; Tharaux, P.-L. MicroRNAs in kidney injury and disease. Nat. Rev. Nephrol. 2022, 18, 643–662. [Google Scholar] [CrossRef]
- Ji, C.; Guo, X. The clinical potential of circulating microRNAs in obesity. Nat. Rev. Endocrinol. 2019, 15, 731–743. [Google Scholar] [CrossRef] [PubMed]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Montoro, J.I.; Morales, E.; Cornejo-Pareja, I.; Tinahones, F.J.; Fernández-García, J.C. Obesity-related glomerulopathy: Current approaches and future perspectives. Obes. Rev. 2022, 23, e13450. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Dabbas, W.; Gangemi, A.; Benedetti, E.; Lash, J.; Finn, P.W.; Perkins, D.L. Obesity Management and Chronic Kidney Disease. Semin Nephrol. 2021, 41, 392–402. [Google Scholar] [CrossRef]
- Kiran, S.; Kumar, V.; Kumar, S.; Price, R.L.; Singh, U.P. Adipocyte, Immune Cells, and miRNA Crosstalk: A Novel Regulator of Metabolic Dysfunction and Obesity. Cells 2021, 10, 1004. [Google Scholar] [CrossRef]
- Martinelli, R.; Nardelli, C.; Pilone, V.; Buonomo, T.; Liguori, R.; Castanò, I.; Buono, P.; Masone, S.; Persico, G.; Forestieri, P.; et al. miR-519d overexpression is associated with human obesity. Obesity 2010, 18, 2170–2176. [Google Scholar] [CrossRef]
- Chen, T.; Zhang, Y.; Liu, Y.; Zhu, D.; Yu, J.; Li, G.; Sun, Z.; Wang, W.; Jiang, H.; Hong, Z. MiR-27a promotes insulin resistance and mediates glucose metabolism by targeting PPAR-γ-mediated PI3K/AKT signaling. Aging 2019, 11, 7510–7524. [Google Scholar] [CrossRef]
- Zhang, Y.; Qian, B.; Yang, Y.; Niu, F.; Lin, C.; Yuan, H.; Wang, J.; Wu, T.; Shao, Y.; Shao, S.; et al. Visceral Adipocyte-Derived Extracellular Vesicle miR-27a-5p Elicits Glucose Intolerance by Inhibiting Pancreatic β-Cell Insulin Secretion. Diabetes 2024, 73, 1832–1847. [Google Scholar] [CrossRef]
- Cho, Y.K.; Son, Y.; Kim, S.-N.; Song, H.-D.; Kim, M.; Park, J.-H.; Jung, Y.-S.; Ahn, S.-Y.; Saha, A.; Granneman, J.G.; et al. MicroRNA-10a-5p regulates macrophage polarization and promotes therapeutic adipose tissue remodeling. Mol. Metab. 2019, 29, 86–98. [Google Scholar] [CrossRef]
- Lee, S.; Cho, Y.K.; Kim, H.; Choi, C.; Kim, S.; Lee, Y.-H. miR-10a regulates cell death and inflammation in adipose tissue of male mice with diet-induced obesity. Mol. Metab. 2024, 90, 102039. [Google Scholar] [CrossRef]
- Zhou, J.; Li, Z.; Wu, T.; Zhao, Q.; Zhao, Q.; Cao, Y. LncGBP9/miR-34a axis drives macrophages toward a phenotype conducive for spinal cord injury repair via STAT1/STAT6 and SOCS3. J. Neuroinflammation 2020, 17, 134. [Google Scholar] [CrossRef]
- Jaiswal, A.; Reddy, S.S.; Maurya, M.; Maurya, P.; Barthwal, M.K. MicroRNA-99a mimics inhibit M1 macrophage phenotype and adipose tissue inflammation by targeting TNFα. Cell Mol Immunol. 2019, 16, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Maurya, M.; Barthwal, M.K. MicroRNA-99a: A potential double-edged sword targeting macrophage inflammation and metabolism. Cell Mol. Immunol. 2021, 18, 2290–2292. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.; Danielson, K.M.; Benton, M.C.; Ziegler, O.; Shah, R.; Stubbs, R.S.; Das, S.; Macartney-Coxson, D. miRNA Signatures of Insulin Resistance in Obesity. Obesity 2017, 25, 1734–1744. [Google Scholar] [CrossRef]
- Agbu, P.; Carthew, R.W. MicroRNA-mediated regulation of glucose and lipid metabolism. Nat. Rev. Mol. Cell Biol. 2021, 22, 425–438. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, A.; Lu, S.; Liu, D.; Xuan, X.; Lei, X.; Zhong, M.; Gao, F. Noncoding RNA profiling in omentum adipose tissue from obese patients and the identification of novel metabolic biomarkers. Front. Genet. 2025, 16. [Google Scholar] [CrossRef]
- Ying, W.; Riopel, M.; Bandyopadhyay, G.; Dong, Y.; Birmingham, A.; Seo, J.B.; Ofrecio, J.M.; Wollam, J.; Hernandez-Carretero, A.; Fu, W.; et al. Adipose Tissue Macrophage-Derived Exosomal miRNAs Can Modulate In Vivo and In Vitro Insulin Sensitivity. Cell 2017, 171, 372–384.e12. [Google Scholar] [CrossRef]
- Castaño, C.; Kalko, S.; Novials, A.; Párrizas, M. Obesity-associated exosomal miRNAs modulate glucose and lipid metabolism in mice. Proc. Natl. Acad. Sci. USA 2018, 115, 12158–12163. [Google Scholar] [CrossRef]
- Trajkovski, M.; Hausser, J.; Soutschek, J.; Bhat, B.; Akin, A.; Zavolan, M.; Heim, M.H.; Stoffel, M. MicroRNAs 103 and 107 regulate insulin sensitivity. Nature 2011, 474, 649–653. [Google Scholar] [CrossRef]
- Kornfeld, J.-W.; Baitzel, C.; Könner, A.C.; Nicholls, H.T.; Vogt, M.C.; Herrmanns, K.; Scheja, L.; Haumaitre, C.; Wolf, A.M.; Knippschild, U.; et al. Obesity-induced overexpression of miR-802 impairs glucose metabolism through silencing of Hnf1b. Nature 2013, 494, 111–115. [Google Scholar] [CrossRef]
- Masoumi-Ardakani, Y.; Eghbalian, M.; Fallah, H.; Jafari, A.; Shahouzehi, B. Exploring serum miR-33b as a novel diagnostic marker for hypercholesterolemia and obesity: Insights from a pilot case-control study. BMC Endocr. Disord. 2025, 25, 27. [Google Scholar] [CrossRef] [PubMed]
- Hanusek, K.; Karczmarski, J.; Litwiniuk, A.; Urbańska, K.; Ambrozkiewicz, F.; Kwiatkowski, A.; Martyńska, L.; Domańska, A.; Bik, W.; Paziewska, A. Obesity as a Risk Factor for Breast Cancer—The Role of miRNA. Int. J. Mol. Sci. 2022, 23, 15683. [Google Scholar] [CrossRef]
- Silveira, A.; Gomes, J.; Roque, F.; Fernandes, T.; de Oliveira, E.M. MicroRNAs in Obesity-Associated Disorders: The Role of Exercise Training. Obes. Facts. 2022, 15, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, T.; Casaes, L.; Soci, Ú.; Silveira, A.; Gomes, J.; Barretti, D.; Roque, F.; Oliveira, E. Exercise Training Restores the Cardiac Microrna-16 Levels Preventing Microvascular Rarefaction in Obese Zucker Rats. Obes. Facts 2018, 11, 15–24. [Google Scholar] [CrossRef]
- Szostak, J.; Gorący, A.; Durys, D.; Dec, P.; Modrzejewski, A.; Pawlik, A. The Role of MicroRNA in the Pathogenesis of Diabetic Nephropathy. Int. J. Mol. Sci. 2023, 24, 6214. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Falck Ørntoft, T. Normalization of Real-Time Quantitative Reverse Transcription-PCR Data: A Model-Based Variance Estimation Approach to Identify Genes Suited for Normalization, Applied to Bladder and Colon Cancer Data Sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
- Shankar, M.; Shetty, A.; N.S., M.; C.G., S.; A., K.; Tennankore, K. Urinary exosomal miRNA signature of IgA nephropathy: A case-control study. Sci. Rep. 2023, 13, 21400. [Google Scholar] [CrossRef]
- Trabulus, S.; Zor, M.S.; Alagoz, S.; Dincer, M.T.; Meşe, M.; Yilmaz, E.; Turanli, E.T.; Seyahi, N. Profiling of five urinary exosomal miRNAs for the differential diagnosis of patients with diabetic kidney disease and focal segmental glomerulosclerosis. PLoS ONE 2024, 19, e0312470. [Google Scholar] [CrossRef]
- Gomez, I.G.; MacKenna, D.A.; Johnson, B.G.; Kaimal, V.; Roach, A.M.; Ren, S.; Nakagawa, N.; Xin, C.; Newitt, R.; Pandya, S.; et al. Anti-microRNA-21 oligonucleotides prevent Alport nephropathy progression by stimulating metabolic pathways. J. Clin. Investig. 2015, 125, 141–156. [Google Scholar] [CrossRef]
- Bais, T.; Gansevoort, R.T.; Meijer, E. Drugs in Clinical Development to Treat Autosomal Dominant Polycystic Kidney Disease. Drugs 2022, 82, 1095–1115. [Google Scholar] [CrossRef]
- Ru, Y.; Kechris, K.J.; Tabakoff, B.; Hoffman, P.; Radcliffe, R.A.; Bowler, R.; Mahaffey, S.; Rossi, S.; Calin, G.A.; Lynne Bemis, L.; et al. The multiMiR R package and database: Integration of microRNA-target interactions along with their disease and drug associations. Nucleic Acids Res. 2014, 42, gku631. [Google Scholar] [CrossRef]
- Sun, D.; Chen, J.; Wu, W.; Tang, J.; Luo, L.; Zhang, K.; Jin, L.; Lin, S.; Gao, Y.; Yan, X.; et al. MiR-802 causes nephropathy by suppressing NF-κB-repressing factor in obese mice and human. J. Cell Mol. Med. 2019, 23, 2863–2871. [Google Scholar] [CrossRef] [PubMed]
- Pereira, B.M.V.; Thieme, K.; de Araújo, L.; Rodrigues, A.C. Lack of adiponectin in mice accelerates high-fat diet-induced progression of chronic kidney disease. Life Sci. 2020, 257, 118061. [Google Scholar] [CrossRef]
- Morrison, M.C.; Yakala, G.K.; Liang, W.; Wielinga, P.Y.; Salic, K.; van Koppen, A.; Tomar, T.; Kleemann, R.; Heeringa, P.; Kooistra, T. Protective effect of rosiglitazone on kidney function in high-fat challenged human-CRP transgenic mice: A possible role for adiponectin and miR-21? Sci. Rep. 2017, 7, 2915. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Zhang, J.; Chen, X.; Zhang, J.; Ding, X.; You, X.; Fan, L.; Chen, C.; Zhou, Y. MicroRNA-155 Mediates Obesity-Induced Renal Inflammation and Dysfunction. Inflammation 2019, 42, 994–1003. [Google Scholar] [CrossRef] [PubMed]
- Ren, Q.; Yu, S.; Zeng, H.; Xia, H. The role of PTEN in puromycin aminonucleoside-induced podocyte injury. Int. J. Med. Sci. 2022, 19, 1451–1459. [Google Scholar] [CrossRef]
- Duan, P.; Tan, J.; Miao, Y.; Zhang, Q. PINK1/Parkin-Mediated Mitophagy Plays a Protective Role in Albumin Overload-Induced Renal Tubular Cell Injury. Front. Biosci. 2022, 27, 184. [Google Scholar] [CrossRef]
- Hu, Z.; Wang, H.; Lee, I.H.; Modi, S.; Wang, X.; Du, J.; Mitch, W.E. PTEN inhibition improves muscle regeneration in mice fed a high-fat diet. Diabetes 2010, 59, 1312–1320. [Google Scholar] [CrossRef]
- Ji, J.; Qin, Y.; Ren, J.; Lu, C.; Wang, R.; Dai, X.; Zhou, R.; Huang, Z.; Xu, M.; Chen, M.; et al. Mitochondria-related miR-141-3p contributes to mitochondrial dysfunction in HFD-induced obesity by inhibiting PTEN. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef]
- Garner, K.L.; Betin, V.M.S.; Pinto, V.; Graham, M.; Abgueguen, E.; Barnes, M.; Bedford, D.C.; McArdle, C.A.; Coward, R.J.M. Enhanced insulin receptor, but not PI3K, signalling protects podocytes from ER stress. Sci. Rep. 2018, 8, 3902. [Google Scholar] [CrossRef]
- An, C.; Jiao, B.; Du, H.; Tran, M.; Zhou, D.; Wang, Y. Myeloid PTEN deficiency aggravates renal inflammation and fibrosis in angiotensin II-induced hypertension. J. Cell Physiol. 2022, 237, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Axelrod, C.L.; Fealy, C.E.; Erickson, M.L.; Davuluri, G.; Fujioka, H.; Dantas, W.S.; Huangc, E.; Pergolaa, K.; Meya, J.T.; King, W.T.; et al. Lipids activate skeletal muscle mitochondrial fission and quality control networks to induce insulin resistance in humans. Metabolism 2021, 121, 154803. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Wang, C.; Zhou, X.; Li, Y.; Ma, Y.; Zhang, R.; Li, R. Downregulation of PTEN promotes podocyte endocytosis of lipids aggravating obesity-related glomerulopathy. Am. J. Physiol. Renal Physiol. 2020, 318, 589–599. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Target Gene | Regulation Status | Involvement with ORG Development | Model Used | Source of Isolation | REFERENCE | |
|---|---|---|---|---|---|---|
| miR-802 | Inhibition of NF-κB-repressing factor (NRF) | Upregulated | Promotes kidney inflammation and fibrosis | C57BL/6J mice and humans | Kidney tissue (mice); serum (humans) | Sun et al. [52] (PMID 30729676) |
| miR-130b | Not identified, correlated with decreased adiponectin | Upregulated | Promotes renal lipid accumulation and albuminuria | adipoKO mice on a C57BL/6 background | Kidney tissue | Pereira et al. [53] (PMID 32652137) |
| miR-21 | Not identified, highly correlated with expression of KIM-1 | Upregulated | Promotes kidney inflammation and fibrosis | HuCRPtg mice on a C57BL/6 background | Kidney tissue | Morrison et al. [54] (PMID 28588299) |
| miR-155 | Inhibition of SHIP1/INNP5D (NF-κB signaling pathway) | Upregulated | Promotes kidney inflammation and oxidative stress | C57BL/6J mice and humans | Kidney tissue | Zheng et al. [55] (PMID 30715692) |
| miR-205 | Inhibition of PTEN | Upregulated | Promotes increased lipid uptake in podocytes | Wistar Han rats | Kidney tissue and urine | López-Martínez et al. [8] (PMID 38928144) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarro-Díaz, M.; López-Martínez, M. The Role of miRNAs as Early Biomarkers in Obesity-Related Glomerulopathy: Implications for Early Detection and Treatment. Biomedicines 2025, 13, 1030. https://doi.org/10.3390/biomedicines13051030
Navarro-Díaz M, López-Martínez M. The Role of miRNAs as Early Biomarkers in Obesity-Related Glomerulopathy: Implications for Early Detection and Treatment. Biomedicines. 2025; 13(5):1030. https://doi.org/10.3390/biomedicines13051030
Chicago/Turabian StyleNavarro-Díaz, Maruja, and Marina López-Martínez. 2025. "The Role of miRNAs as Early Biomarkers in Obesity-Related Glomerulopathy: Implications for Early Detection and Treatment" Biomedicines 13, no. 5: 1030. https://doi.org/10.3390/biomedicines13051030
APA StyleNavarro-Díaz, M., & López-Martínez, M. (2025). The Role of miRNAs as Early Biomarkers in Obesity-Related Glomerulopathy: Implications for Early Detection and Treatment. Biomedicines, 13(5), 1030. https://doi.org/10.3390/biomedicines13051030