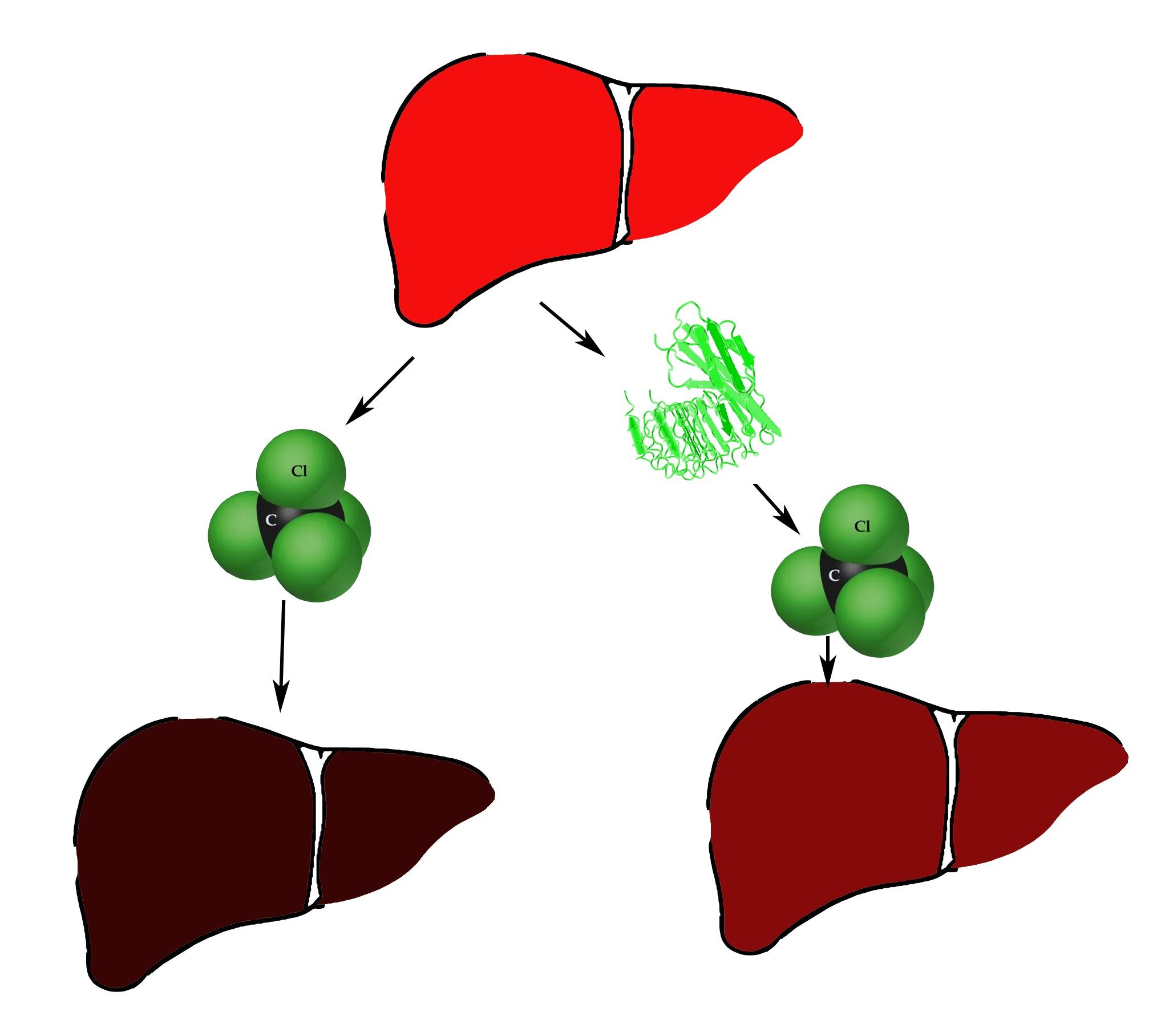
Hepatoprotective Activity of InlB321/15, the HGFR Ligand of Bacterial Origin, in CCI4-Induced Acute Liver Injury Mice

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Experimental Section
2.1. Animals
2.2. Cell Cultures
2.3. Antibodies and Growth Factor
2.4. InlB321/15 Purification
2.5. In Vitro Cell Viability Assay
2.6. Immunofluorescence Microscopy
2.7. SDS-PAGE and Immunoblotting
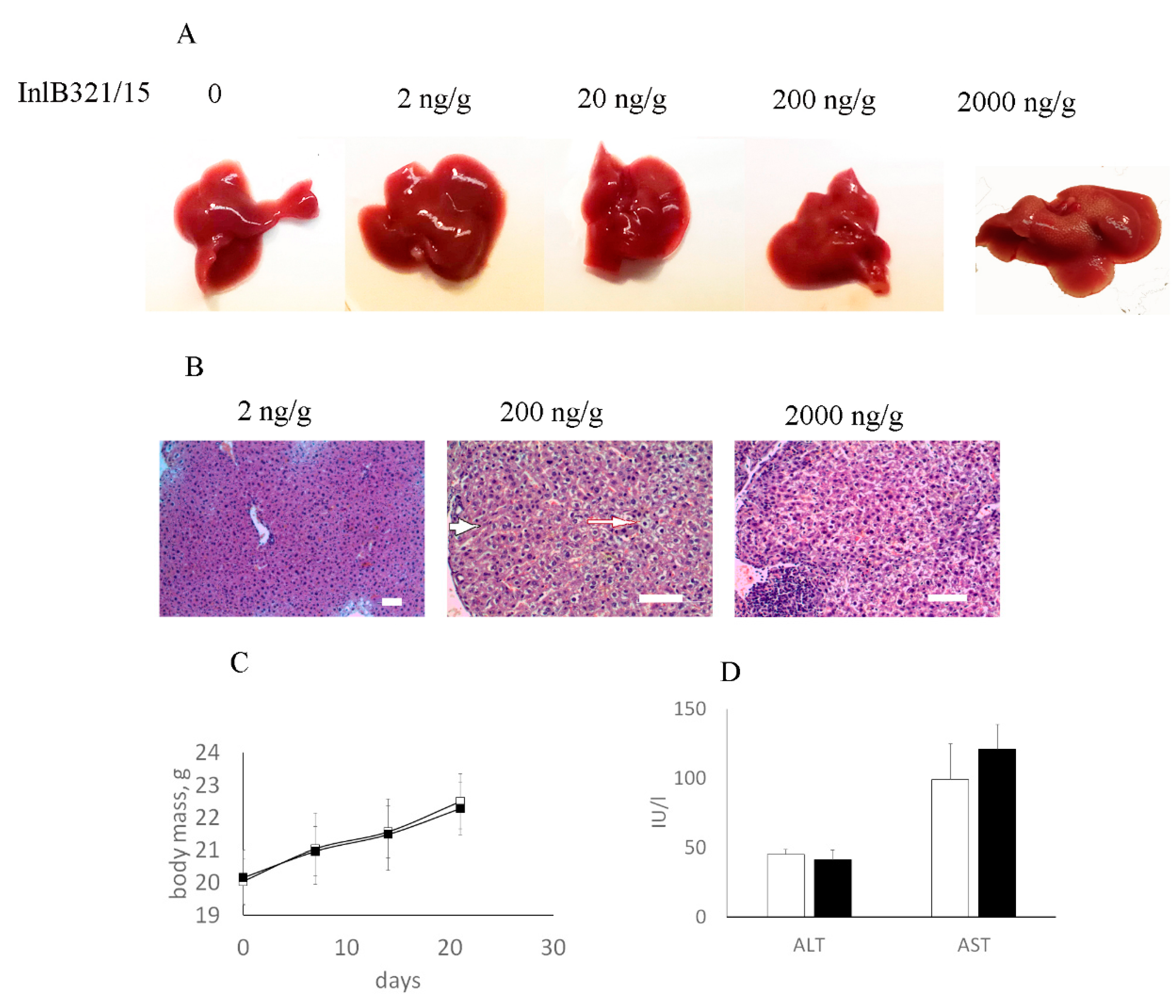
2.8. InlB321/15 Toxicity Assay
2.9. CCl4 Acute Toxicity Testing
2.10. Histological Examination of Liver Sections
2.11. Biochemical Markers
2.12. Statistics
3. Results
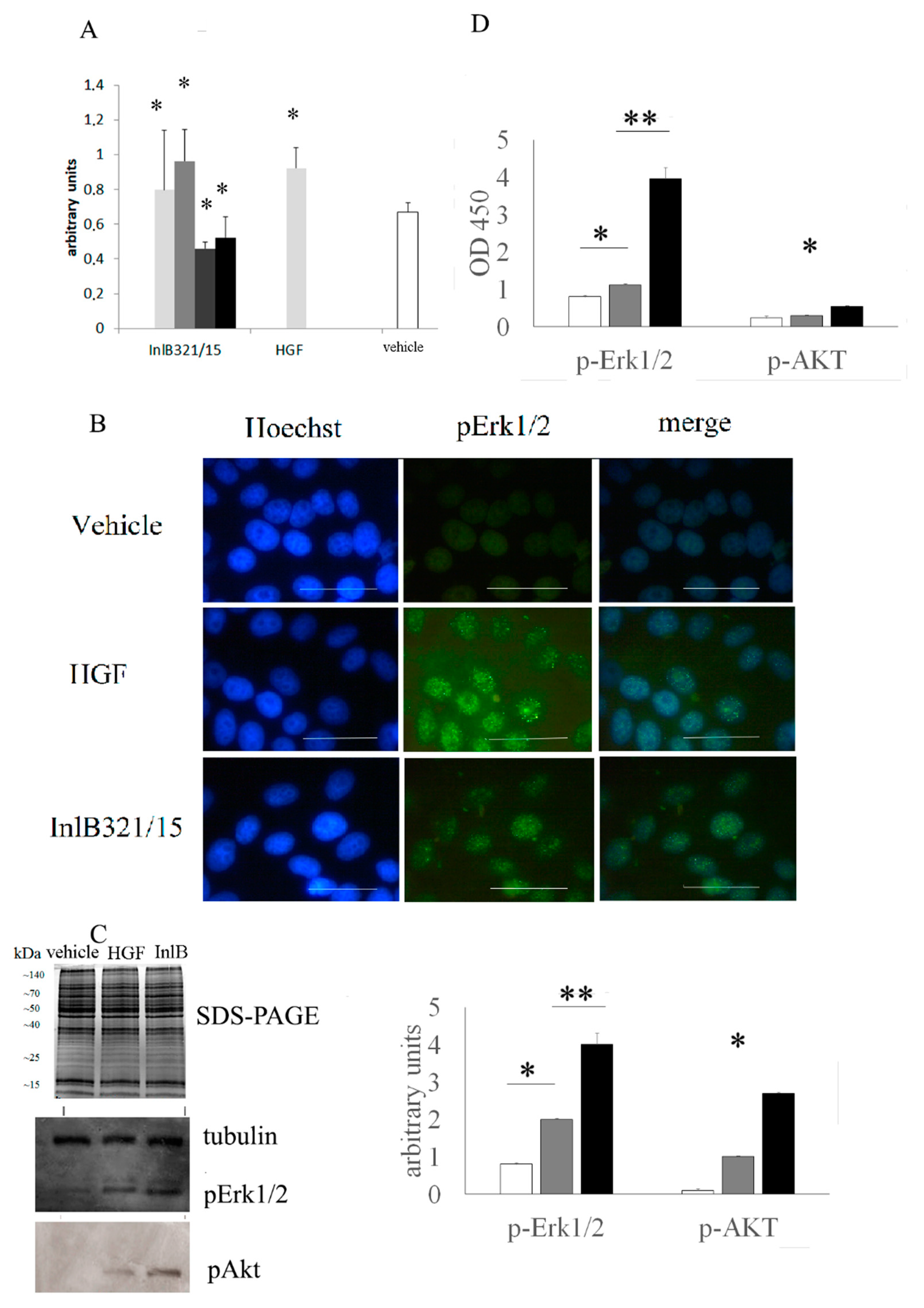
3.1. In Vitro InlB321/15 Mitogenic Activity
3.2. In Vitro inlb321/15-Stimulated MAPK- and PI3K/Akt Signaling Pathway Activation
3.3. InlB321/15 Toxicity in Mice
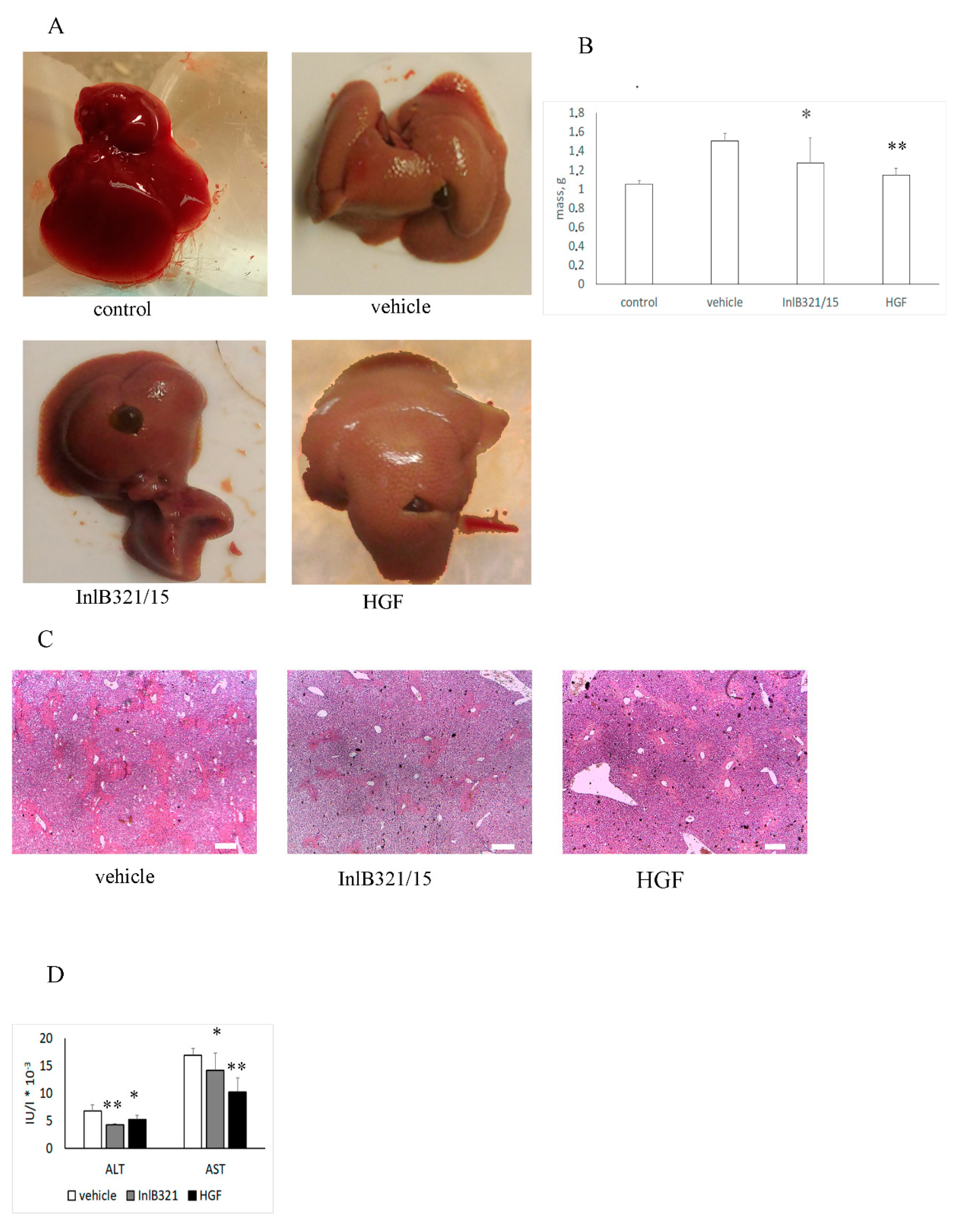
3.4. InlB321/15 Hepatoprotective Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Michalopoulos, G.K. Liver Regeneration. J. Cell Physiol. 2007, 213, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Huh, C.-G.; Factor, V.M.; Sánchez, A.; Uchida, K.; Conner, E.A.; Thorgeirsson, S.S. Hepatocyte growth factor/c-met signaling pathway is required for efficient liver regeneration and repair. Proc. Natl. Acad. Sci. USA 2004, 101, 4477–4482. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, T.; Factor, V.M.; Marquardt, J.U.; Raggi, C.; Seo, D.; Kitade, M.; Conner, E.A.; Thorgeirsson, S.S. Hepatocyte growth factor/c-met signaling is required for stem-cell-mediated liver regeneration in mice. Hepatology 2012, 55, 1215–1226. [Google Scholar] [CrossRef]
- Marquardt, J.U.; Seo, D.; Gómez-Quiroz, L.E.; Uchida, K.; Gillen, M.C.; Kitade, M.; Kaposi-Novak, P.; Conner, E.A.; Factor, V.M.; Thorgeirsson, S.S. Loss of c-Met accelerates development of liver fibrosis in response to CCl 4 exposure through deregulation of multiple molecular pathways. Biochim. Biophys. Acta Mol. Basis Dis. 2012, 1822, 942–951. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Funakoshi, H.; Takahashi, H.; Sakai, K. HGF-Met Pathway in Regeneration and Drug Discovery. Biomedicines 2014, 2, 275–300. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, T.; Takagi, H.; Horiguchi, N.; Toyoda, M.; Sato, K.; Takayama, H.; Mori, M. CCl4-induced acute liver injury in mice is inhibited by hepatocyte growth factor overexpression but stimulated by NK2 overexpression. FEBS Lett. 2002, 532, 391–395. [Google Scholar] [CrossRef]
- Matsumoto, K.; Nakamura, T.; Sakai, K.; Nakamura, T. Hepatocyte growth factor and Met in tumor biology and therapeutic approach with NK4. Proteomics 2008, 8, 3360–3370. [Google Scholar] [CrossRef] [PubMed]
- Imamura, R.; Matsumoto, K. Hepatocyte growth factor in physiology and infectious diseases. Cytokine 2017, 98, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Veiga, E.; Cossart, P. Listeria hijacks the clathrin-dependent endocytic machinery to invade mammalian cells. Nat. Cell Biol. 2005, 7, 894. [Google Scholar] [CrossRef]
- Niemann, H.H.; Jäger, V.; Butler, P.J.G.; van den Heuvel, J.; Schmidt, S.; Ferraris, D.; Gherardi, E.; Heinz, D.W. Structure of the Human Receptor Tyrosine Kinase Met in Complex with the Listeria Invasion Protein InlB. Cell 2007, 130, 235–246. [Google Scholar] [CrossRef]
- Bierne, H.; Cossart, P. InlB, a surface protein of Listeria monocytogenes that behaves as an invasin and a growth factor. J. Cell Sci. 2002, 115, 3357–3367. [Google Scholar]
- Li, N.; Xiang, G.S.; Dokainish, H.; Ireton, K.; Elferink, L.A. The Listeria protein internalin B mimics hepatocyte growth factor-induced receptor trafficking. Traffic 2005, 6, 459–473. [Google Scholar] [CrossRef] [PubMed]
- Chalenko, Y.M.; Sysolyatina, E.V.; Kalinin, E.V.; Sobyanin, K.A.; Ermolaeva, S.A. Natural variants of Listeria monocytogenes internalin B with different ability to stimulate cell proliferation and cytoskeleton rearrangement in HEp-2 cells. Mol. Genet. Microbiol. Virol. 2017, 32, 80–86. [Google Scholar] [CrossRef]
- Braun, L.; Nato, F.; Payrastre, B.; Mazié, J.C.; Cossart, P. The 213-amino-acid leucine-rich repeat region of the Listeria monocytogenes InlB protein is sufficient for entry into mammalian cells, stimulation of PI 3-kinase and membrane ruffling. Mol. Microbiol. 1999, 34, 10–23. [Google Scholar] [CrossRef]
- Ferraris, D.M.; Gherardi, E.; Di, Y.; Heinz, D.W.; Niemann, H.H. Ligand-Mediated Dimerization of the Met Receptor Tyrosine Kinase by the Bacterial Invasion Protein InlB. J. Mol. Biol. 2010, 395, 522–532. [Google Scholar] [CrossRef] [PubMed]
- Mungunsukh, O.; Lee, Y.H.; Marquez, A.P.; Cecchi, F.; Bottaro, D.P.; Day, R.M. A tandem repeat of a fragment of Listeria monocytogenes internalin B protein induces cell survival and proliferation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2010, 299, L905–L914. [Google Scholar] [PubMed]
- Kolditz, F.; Krausze, J.; Heinz, D.W.; Niemann, H.H.; Müller-Goymann, C.C. Wound healing potential of a dimeric InlB variant analyzed by in vitro experiments on re-epithelialization of human skin models. Eur. J. Pharm. Biopharm. 2014, 86, 277–283. [Google Scholar] [CrossRef]
- Usatyuk, P.V.; Fu, P.; Mohan, V.; Epshtein, Y.; Jacobson, J.R.; Gomez-Cambronero, J.; Wary, K.K.; Bindokas, V.; Dudek, S.M.; Salgia, R.; et al. Role of c-Met/phosphatidylinositol 3-kinase (PI3k)/Akt signaling in Hepatocyte growth factor (HGF)-mediated lamellipodia formation, reactive oxygen species (ROS) generation, and motility of lung endothelial cells. J. Biol. Chem. 2014, 289, 13476–13491. [Google Scholar] [CrossRef]
- Bladt, F.; Riethmacher, D.; Isenmann, S.; Aguzzi, A.; Birchmeier, C. Essential role for the c-met receptor in the migration of myogenic precursor cells into the limb bud. Nature 1995, 376, 768–771. [Google Scholar] [CrossRef]
- Borowiak, M.; Garratt, A.N.; Wüstefeld, T.; Strehle, M.; Trautwein, C.; Birchmeier, C. Met provides essential signals for liver regeneration. Proc. Natl. Acad. Sci. USA 2004, 101, 10608–10613. [Google Scholar] [CrossRef]
- Okano, J.I.; Shiota, G.; Matsumoto, K.; Yasui, S.; Kurimasa, A.; Hisatome, I.; Steinberg, P.; Murawaki, Y. Hepatocyte growth factor exerts a proliferative effect on oval cells through the PI3K/AKT signaling pathway. Biochem. Biophys. Res. Commun. 2003, 309, 298–304. [Google Scholar] [CrossRef]
- Pauta, M.; Rotllan, N.; Fernández-Hernando, A.; Langhi, C.; Ribera, J.; Lu, M.; Boix, L.; Bruix, J.; Jimenez, W.; Suárez, Y.; et al. Akt-mediated foxo1 inhibition is required for liver regeneration. Hepatology 2016, 63, 1660–1674. [Google Scholar] [CrossRef] [PubMed]
- Kanayama, M.; Takahara, T.; Yata, Y.; Xue, F.; Shinno, E.; Nonome, K.; Kudo, H.; Kawai, K.; Kudo, T.; Tabuchi, Y.; et al. Hepatocyte growth factor promotes colonic epithelial regeneration via Akt signaling. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 293, G230–G239. [Google Scholar] [CrossRef]
- Xiao, G.-H. Anti-apoptotic signaling by hepatocyte growth factor/Met via the phosphatidylinositol 3-kinase/Akt and mitogen-activated protein kinase pathways. Proc. Natl. Acad. Sci. USA 2001, 98, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Factor, V.M.; Seo, D.; Ishikawa, T.; Kaposi-Novak, P.; Marquardt, J.U.; Andersen, J.B.; Conner, E.A.; Thorgeirsson, S.S. Loss of c-Met disrupts gene expression program required for G2/M progression during liver regeneration in mice. PLoS ONE 2010, 5, e12739. [Google Scholar] [CrossRef] [PubMed]
- Appasamy, R.; Tanabe, M.; Murase, N.; Zarnegar, R.; Venkataramanan, R.; Van Thiel, D.H.; Michalopoulos, G.K. Hepatocyte growth factor, blood clearance, organ uptake, and biliary excretion in normal and partially hepatectomized rats. Lab. Investig. 1993, 68, 270–276. [Google Scholar] [PubMed]
- Nakamura, T.; Nishizawa, T.; Hagiya, M.; Seki, T.; Shimonishi, M.; Sugimura, A.; Tashiro, K.; Shimizu, S. Molecular cloning and expression of human hepatocyte growth factor. Nature 1989, 342, 440–443. [Google Scholar] [CrossRef] [PubMed]
- Morishita, R.; Makino, H.; Aoki, M.; Hashiya, N.; Yamasaki, K.; Azuma, J.; Taniyama, Y.; Sawa, Y.; Kaneda, Y.; Ogihara, T. Phase I/IIa clinical trial of therapeutic angiogenesis using hepatocyte growth factor gene transfer to treat critical limb ischemia. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 713–720. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chalenko, Y.; Sobyanin, K.; Sysolyatina, E.; Midiber, K.; Kalinin, E.; Lavrikova, A.; Mikhaleva, L.; Ermolaeva, S. Hepatoprotective Activity of InlB321/15, the HGFR Ligand of Bacterial Origin, in CCI4-Induced Acute Liver Injury Mice. Biomedicines 2019, 7, 29. https://doi.org/10.3390/biomedicines7020029
Chalenko Y, Sobyanin K, Sysolyatina E, Midiber K, Kalinin E, Lavrikova A, Mikhaleva L, Ermolaeva S. Hepatoprotective Activity of InlB321/15, the HGFR Ligand of Bacterial Origin, in CCI4-Induced Acute Liver Injury Mice. Biomedicines. 2019; 7(2):29. https://doi.org/10.3390/biomedicines7020029
Chicago/Turabian StyleChalenko, Yaroslava, Konstantin Sobyanin, Elena Sysolyatina, Konstantin Midiber, Egor Kalinin, Alexandra Lavrikova, Lyudmila Mikhaleva, and Svetlana Ermolaeva. 2019. "Hepatoprotective Activity of InlB321/15, the HGFR Ligand of Bacterial Origin, in CCI4-Induced Acute Liver Injury Mice" Biomedicines 7, no. 2: 29. https://doi.org/10.3390/biomedicines7020029
APA StyleChalenko, Y., Sobyanin, K., Sysolyatina, E., Midiber, K., Kalinin, E., Lavrikova, A., Mikhaleva, L., & Ermolaeva, S. (2019). Hepatoprotective Activity of InlB321/15, the HGFR Ligand of Bacterial Origin, in CCI4-Induced Acute Liver Injury Mice. Biomedicines, 7(2), 29. https://doi.org/10.3390/biomedicines7020029