Garcinol Exhibits Anti-Neoplastic Effects by Targeting Diverse Oncogenic Factors in Tumor Cells

 ,
,  ,
, Manoj_Kumar.jpg) and
and
Abstract
:1. Introduction
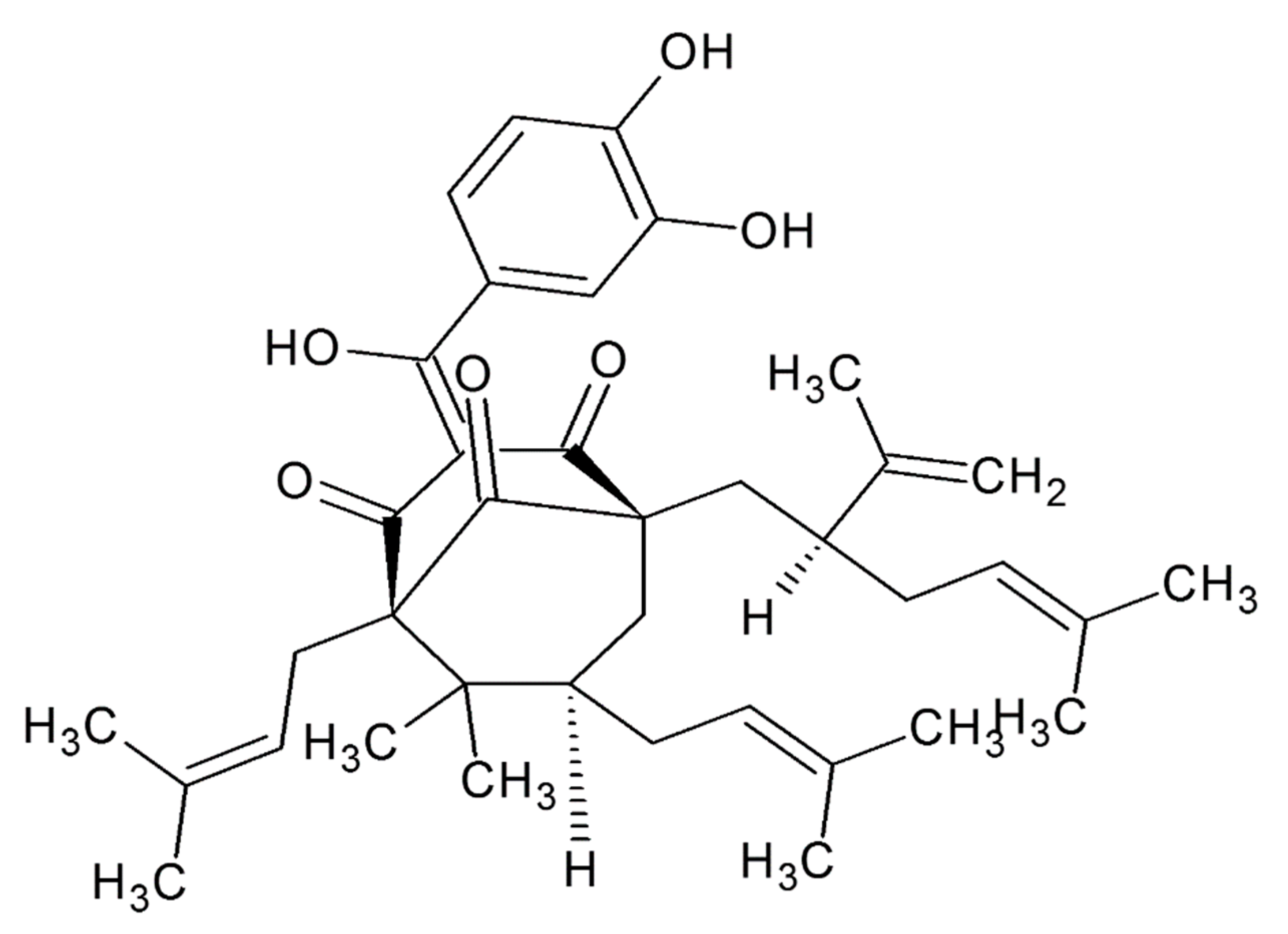
2. Chemistry of Garcinol
Structure Activity Relationship
3. Anti-Neoplastic Regulation of Cellular Processes in Tumor
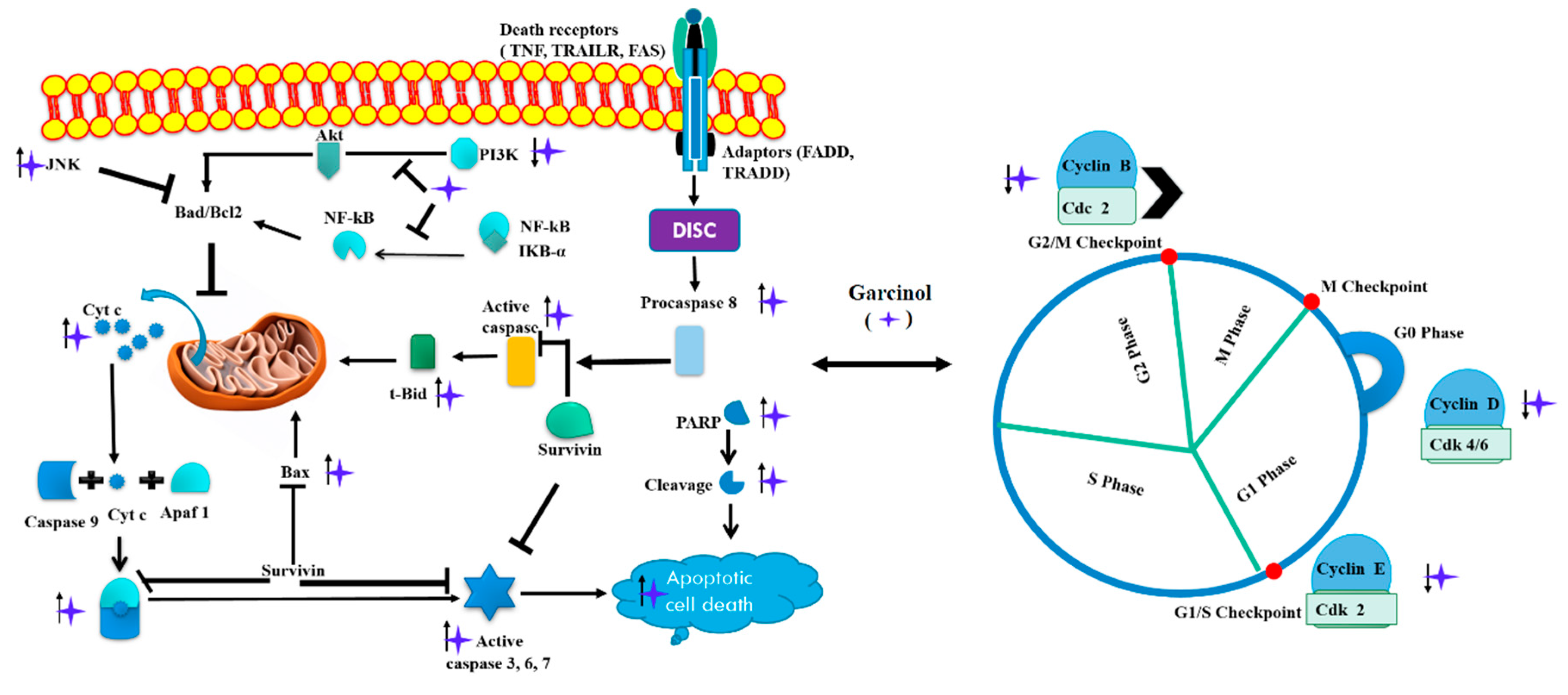
3.1. Apoptosis and Cell Cycle Arrest
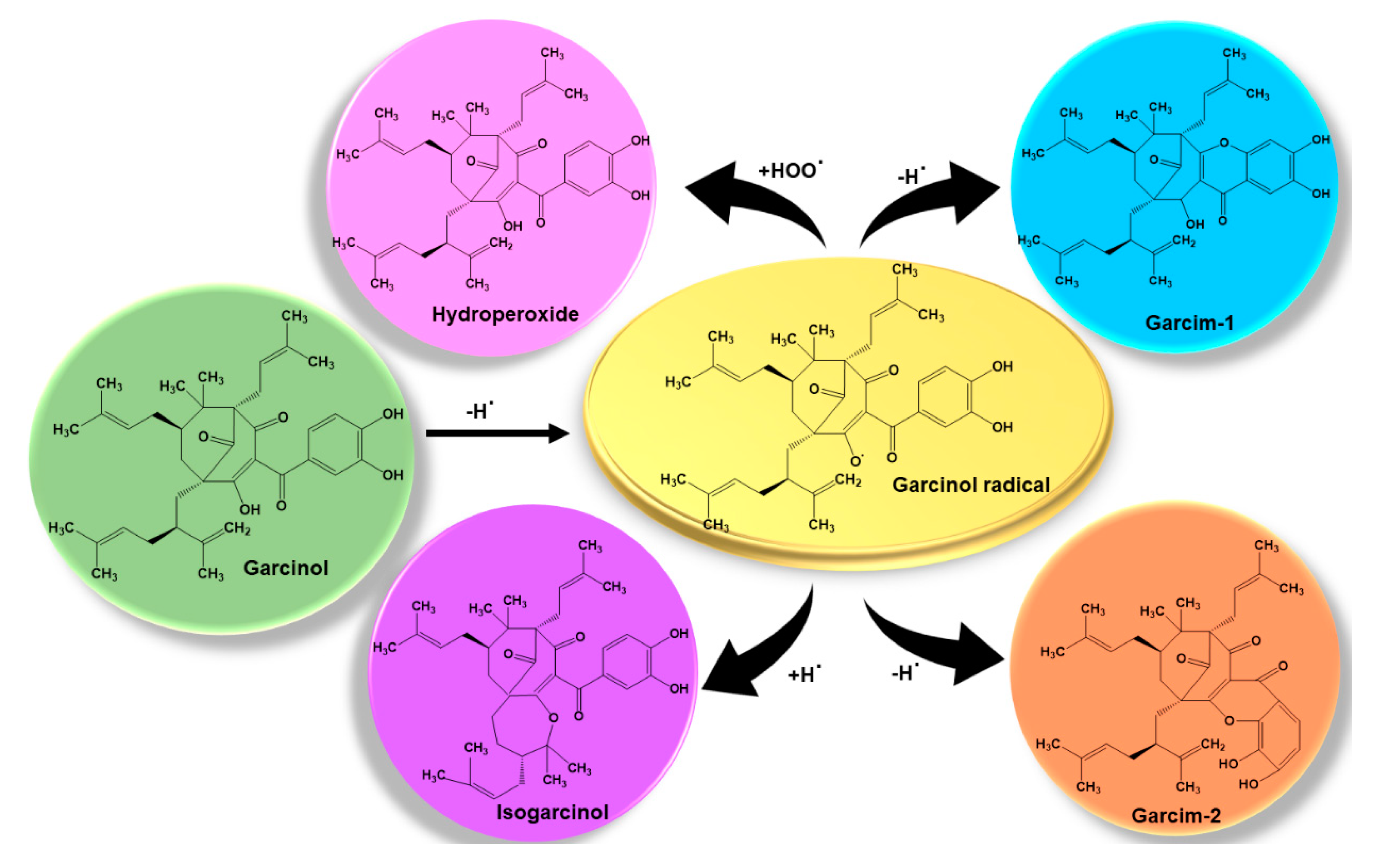
3.2. Antioxidant and Anti-Inflammatory
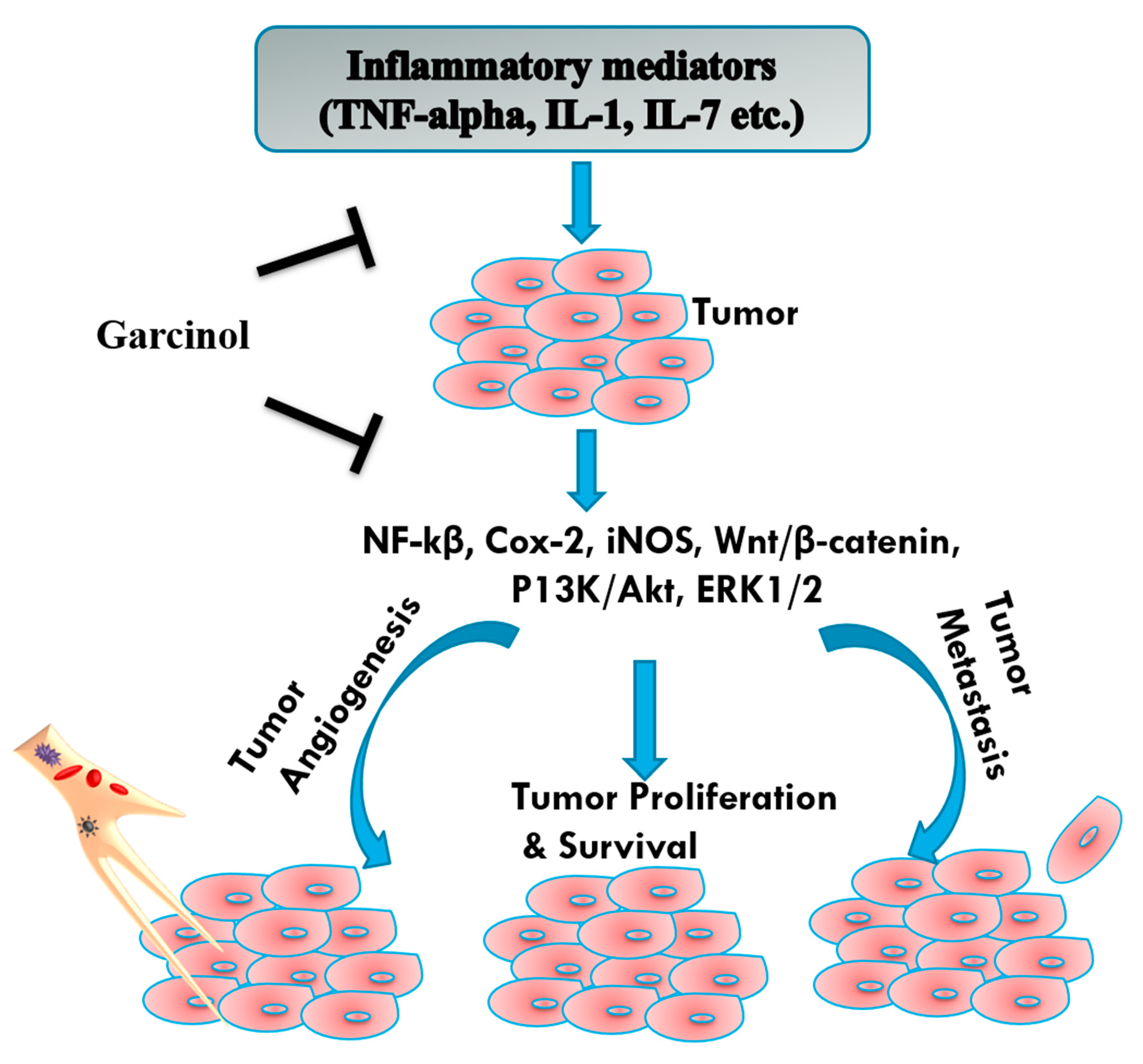
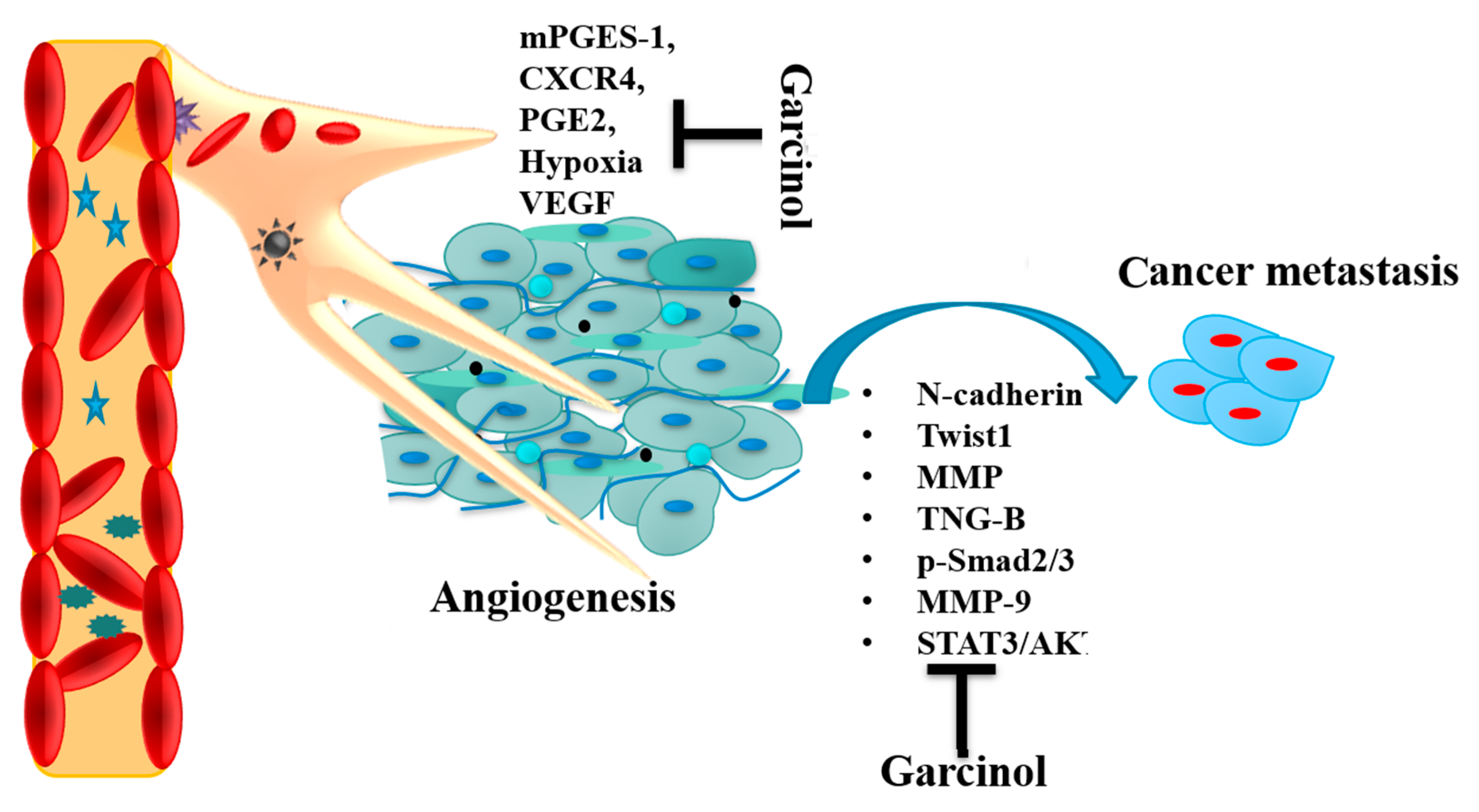
3.3. Angiogenesis and Metastasis
3.4. MicroRNAs (miRNA)
4. Synergistic Effects of Garcinol
5. Bioavailability and Nanotechnology Studies of Garcinol
6. Conclusions and Future Perspective
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Pineros, M.; Znaor, A.; Bray, F. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef] [Green Version]
- Koh, Y.C.; Ho, C.T.; Pan, M.H. Recent advances in cancer chemoprevention with phytochemicals. J. Food Drug Anal. 2020, 28, 14–37. [Google Scholar] [CrossRef]
- Dai, X.; Zhang, J.; Arfuso, F.; Chinnathambi, A.; Zayed, M.E.; Alharbi, S.A.; Kumar, A.P.; Ahn, K.S.; Sethi, G. Targeting TNF-related apoptosis-inducing ligand (TRAIL) receptor by natural products as a potential therapeutic approach for cancer therapy. Exp. Biol. Med. 2015, 240, 760–773. [Google Scholar] [CrossRef] [Green Version]
- Deng, S.; Shanmugam, M.K.; Kumar, A.P.; Yap, C.T.; Sethi, G.; Bishayee, A. Targeting autophagy using natural compounds for cancer prevention and therapy. Cancer 2019, 125, 1228–1246. [Google Scholar] [CrossRef]
- Prasannan, R.; Kalesh, K.A.; Shanmugam, M.K.; Nachiyappan, A.; Ramachandran, L.; Nguyen, A.H.; Kumar, A.P.; Lakshmanan, M.; Ahn, K.S.; Sethi, G. Key cell signaling pathways modulated by zerumbone: Role in the prevention and treatment of cancer. Biochem. Pharmacol. 2012, 84, 1268–1276. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, L.; Manu, K.A.; Shanmugam, M.K.; Li, F.; Siveen, K.S.; Vali, S.; Kapoor, S.; Abbasi, T.; Surana, R.; Smoot, D.T.; et al. Isorhamnetin inhibits proliferation and invasion and induces apoptosis through the modulation of peroxisome proliferator-activated receptor gamma activation pathway in gastric cancer. J. Biol. Chem. 2012, 287, 38028–38040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tewari, D.; Nabavi, S.F.; Nabavi, S.M.; Sureda, A.; Farooqi, A.A.; Atanasov, A.G.; Vacca, R.A.; Sethi, G.; Bishayee, A. Targeting activator protein 1 signaling pathway by bioactive natural agents: Possible therapeutic strategy for cancer prevention and intervention. Pharmacol. Res. 2018, 128, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Aghajanpour, M.; Nazer, M.R.; Obeidavi, Z.; Akbari, M.; Ezati, P.; Kor, N.M. Functional foods and their role in cancer prevention and health promotion: A comprehensive review. Am. J. Cancer Res. 2017, 7, 740–769. [Google Scholar] [PubMed]
- Schobert, R.; Biersack, B. Chemical and Biological Aspects of Garcinol and Isogarcinol: Recent Developments. Chem. Biodivers. 2019, 16, e1900366. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Shanmugam, M.K.; Siveen, K.S.; Wang, F.; Ong, T.H.; Loo, S.Y.; Swamy, M.M.; Mandal, S.; Kumar, A.P.; Goh, B.C.; et al. Garcinol sensitizes human head and neck carcinoma to cisplatin in a xenograft mouse model despite downregulation of proliferative biomarkers. Oncotarget 2015, 6, 5147–5163. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Ho, P.C.; Wong, F.C.; Sethi, G.; Wang, L.Z.; Goh, B.C. Garcinol: Current status of its anti-oxidative, anti-inflammatory and anti-cancer effects. Cancer Lett. 2015, 362, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, V.; Banday, A.Z.; Jindal, A.K.; Das, J.; Rawat, A. Recent advances in elucidating the genetics of common variable immunodeficiency. Genes Dis. 2020, 7, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, V.; Priyanka, K.; Tuli, H.S. Emergence of Circulating MicroRNAs in Breast Cancer as Diagnostic and Therapeutic Efficacy Biomarkers. Mol. Diagn. Ther. 2020, 24, 153–173. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, V.; Tuli, H.S.; Thakral, F.; Singhal, P.; Aggarwal, D.; Srivastava, S.; Pandey, A.; Sak, K.; Varol, M.; Khan, M.A.; et al. Molecular mechanisms of action of hesperidin in cancer: Recent trends and advancements. Exp. Biol. Med. 2020, 245, 486–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aggarwal, V.; Tuli, H.S.; Varol, A.; Thakral, F.; Yerer, M.B.; Sak, K.; Varol, M.; Jain, A.; Khan, M.A.; Sethi, G. Role of Reactive Oxygen Species in Cancer Progression: Molecular Mechanisms and Recent Advancements. Biomolecules 2019, 9, 735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aggarwal, V.; Kashyap, D.; Sak, K.; Tuli, H.S.; Jain, A.; Chaudhary, A.; Garg, V.K.; Sethi, G.; Yerer, M.B. Molecular Mechanisms of Action of Tocotrienols in Cancer: Recent Trends and Advancements. Int. J. Mol. Sci. 2019, 20, 656. [Google Scholar] [CrossRef] [Green Version]
- Das, J.; Sharma, A.; Jindal, A.; Aggarwal, V.; Rawat, A. Leukocyte adhesion defect: Where do we stand circa 2019? Genes. Dis. 2020, 7, 107–114. [Google Scholar] [CrossRef]
- Tuli, H.S.; Tuorkey, M.J.; Thakral, F.; Sak, K.; Kumar, M.; Sharma, A.K.; Sharma, U.; Jain, A.; Aggarwal, V.; Bishayee, A. Molecular Mechanisms of Action of Genistein in Cancer: Recent Advances. Front. Pharmacol. 2019, 10, 1336. [Google Scholar] [CrossRef] [Green Version]
- Behera, A.K.; Swamy, M.M.; Natesh, N.; Kundu, T.K. Garcinol and Its Role in Chronic Diseases. Adv. Exp. Med. Biol. 2016, 928, 435–452. [Google Scholar]
- Farhan, M.; Malik, A.; Ullah, M.F.; Afaq, S.; Faisal, M.; Farooqi, A.A.; Biersack, B.; Schobert, R.; Ahmad, A. Garcinol Sensitizes NSCLC Cells to Standard Therapies by Regulating EMT-Modulating miRNAs. Int. J. Mol. Sci. 2019, 20, 800. [Google Scholar] [CrossRef] [Green Version]
- Tu, S.H.; Chiou, Y.S.; Kalyanam, N.; Ho, C.T.; Chen, L.C.; Pan, M.H. Garcinol sensitizes breast cancer cells to Taxol through the suppression of caspase-3/iPLA2 and NF-kappaB/Twist1 signaling pathways in a mouse 4T1 breast tumor model. Food Funct. 2017, 8, 1067–1079. [Google Scholar] [CrossRef] [PubMed]
- Fernando, H.N.; Kumarasinghe, K.; Gunasekara, T.; Wijekoon, H.; Ekanayaka, E.; Rajapaksha, S.P.; Fernando, S.S.N.; Jayaweera, P.M. Synthesis, Characterization and Antimicrobial Activity of Garcinol Capped Silver Nanoparticles. J. Microbiol. Biotechnol. 2019, 29, 1841–1851. [Google Scholar] [CrossRef] [PubMed]
- Ullah, M.F.; Ahmad, A. Critical Dietary Factors in Cancer Chemoprevention; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Hemshekhar, M.; Sunitha, K.; Santhosh, M.S.; Devaraja, S.; Kemparaju, K.; Vishwanath, B.; Niranjana, S.; Girish, K. An overview on genus Garcinia: Phytochemical and therapeutical aspects. Phytochem. Rev. 2011, 10, 325–351. [Google Scholar] [CrossRef]
- Liu, H.; Gan, F.; Jin, S.; Li, J.; Chen, Y.; Yang, G. Acylphloroglucinol and tocotrienol derivatives from the fruits of Garcinia multiflora. RSC Adv. 2017, 7, 29295–29301. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, A.; Yasmin, T.; Bagchi, D.; Stohs, S.J. The bactericidal effects of Lactobacillus acidophilus, garcinol and Protykin® compared to clarithromycin, on Helicobacter pylori. Mol. Cell Biochem. 2003, 243, 29–35. [Google Scholar] [CrossRef]
- Zheng, D.; Zhang, H.; Zheng, C.-W.; Lao, Y.-Z.; Xu, D.-Q.; Xiao, L.-B.; Xu, H.-X. Garciyunnanimines A–C, novel cytotoxic polycyclic polyprenylated acylphloroglucinol imines from Garcinia yunnanensis. Org. Chem. Front. 2017, 4, 2102–2108. [Google Scholar] [CrossRef]
- Jackson, D.N.; Yang, L.; Wu, S.; Kennelly, E.J.; Lipke, P.N. Garcinia xanthochymus benzophenones promote hyphal apoptosis and potentiate activity of fluconazole against Candida albicans biofilms. Antimicrob. Agents Chemother. 2015, 59, 6032–6038. [Google Scholar] [CrossRef] [Green Version]
- Anu Aravind, A.; Asha, K.; Rameshkumar, K. Phytochemical analysis and antioxidant potential of the leaves of Garcinia travancorica Bedd. Nat. Prod. Res. 2016, 30, 232–236. [Google Scholar] [CrossRef]
- Stark, T.D.; Salger, M.; Frank, O.; Balemba, O.B.; Wakamatsu, J.; Hofmann, T. Antioxidative compounds from Garcinia buchananii stem bark. J. Nat. Prod. 2015, 78, 234–240. [Google Scholar] [CrossRef]
- Wang, L.; Wang, M.; Guo, H.; Zhao, H. Emerging Role of Garcinol in Targeting Cancer Stem Cells of Non-small Cell Lung Cancer. Curr. Pharmacol. Rep. 2019, 5, 14–19. [Google Scholar] [CrossRef]
- Hatcher, H.; Planalp, R.; Cho, J.; Torti, F.M.; Torti, S.V. Curcumin: From ancient medicine to current clinical trials. Cell Mol. Life Sci. 2008, 65, 1631–1652. [Google Scholar] [CrossRef]
- Padhye, S.; Ahmad, A.; Oswal, N.; Sarkar, F.H. Emerging role of Garcinol, the antioxidant chalcone from Garcinia indica Choisy and its synthetic analogs. J. Hematol. Oncol. 2009, 2, 38. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, B.B.; Sung, B. Pharmacological basis for the role of curcumin in chronic diseases: An age-old spice with modern targets. Trends Pharmacol. Sci. 2009, 30, 85–94. [Google Scholar] [CrossRef]
- Zhou, X.Y.; Cao, J.; Han, C.M.; Li, S.W.; Zhang, C.; Du, Y.D.; Zhou, Q.Q.; Zhang, X.Y.; Chen, X. The C8 side chain is one of the key functional group of Garcinol for its anti-cancer effects. Bioorg. Chem. 2017, 71, 74–80. [Google Scholar] [CrossRef]
- Padhye, S.; Ahmad, A.; Oswal, N.; Dandawate, P.; Rub, R.A.; Deshpande, J.; Swamy, K.V.; Sarkar, F.H. Fluorinated 2′-hydroxychalcones as garcinol analogs with enhanced antioxidant and anticancer activities. Bioorg. Med. Chem. Lett. 2010, 20, 5818–5821. [Google Scholar] [CrossRef]
- Praharaj, P.P.; Naik, P.P.; Panigrahi, D.P.; Bhol, C.S.; Mahapatra, K.K.; Patra, S.; Sethi, G.; Bhutia, S.K. Intricate role of mitochondrial lipid in mitophagy and mitochondrial apoptosis: Its implication in cancer therapeutics. Cell Mol. Life. Sci. 2019, 76, 1641–1652. [Google Scholar] [CrossRef]
- Tian, Z.; Shen, J.; Wang, F.; Xiao, P.; Yang, J.; Lei, H.; Kazlauskas, A.; Kohane, I.S.; Wu, E. Cambogin is preferentially cytotoxic to cells expressing PDGFR. PLoS ONE 2011, 6, e21370. [Google Scholar] [CrossRef] [Green Version]
- Shao, S.; Duan, W.; Xu, Q.; Li, X.; Han, L.; Li, W.; Zhang, D.; Wang, Z.; Lei, J. Curcumin Suppresses Hepatic Stellate Cell-Induced Hepatocarcinoma Angiogenesis and Invasion through Downregulating CTGF. Oxid. Med. Cell Longev. 2019, 2019, 8148510. [Google Scholar] [CrossRef]
- Aggarwal, S.; Das, S.N. Garcinol inhibits tumour cell proliferation, angiogenesis, cell cycle progression and induces apoptosis via NF-kappaB inhibition in oral cancer. Tumour Biol. 2016, 37, 7175–7184. [Google Scholar] [CrossRef]
- Dong, H.T.; Cao, J.; Han, C.M.; Su, Y.; Zhang, X.Y.; Chen, X. [Role of 8-allyl Garcinol in the Chemoprevention of Oral Squamous Cell Carcinoma]. Zhongguo Yi Xue Ke Xue Yuan Xue Bao 2019, 41, 1–10. [Google Scholar]
- Zhao, J.; Yang, T.; Ji, J.; Li, C.; Li, Z.; Li, L. Garcinol exerts anti-cancer effect in human cervical cancer cells through upregulation of T-cadherin. Biomed. Pharmacother. 2018, 107, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, B.; Kandimalla, R.; Elancheran, R.; Bharali, R.; Kotoky, J. Garcinia morella fruit, a promising source of antioxidant and anti-inflammatory agents induces breast cancer cell death via triggering apoptotic pathway. Biomed. Pharmacother. 2018, 103, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Mohibi, S.; Mirza, S.; Band, H.; Band, V. Epidermal Growth Factor Receptor activation promotes ADA3 acetylation through the AKT-p300 pathway. Cell Cycle 2017, 16, 1515–1525. [Google Scholar] [CrossRef] [Green Version]
- Shen, K.; Lu, F.; Xie, J.; Wu, M.; Cai, B.; Liu, Y.; Zhang, H.; Tan, H.; Pan, Y.; Xu, H. Cambogin exerts anti-proliferative and pro-apoptotic effects on breast adenocarcinoma through the induction of NADPH oxidase 1 and the alteration of mitochondrial morphology and dynamics. Oncotarget 2016, 7, 50596–50611. [Google Scholar] [CrossRef]
- Shen, K.; Xie, J.; Wang, H.; Zhang, H.; Yu, M.; Lu, F.; Tan, H.; Xu, H. Cambogin Induces Caspase-Independent Apoptosis through the ROS/JNK Pathway and Epigenetic Regulation in Breast Cancer Cells. Mol. Cancer Ther. 2015, 14, 1738–1749. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.; Yuan, L.; Zhang, L.; Zhao, J.; Zhang, C.M.; Deng, H.Y. Garcinol, an acetyltransferase inhibitor, suppresses proliferation of breast cancer cell line MCF-7 promoted by 17beta-estradiol. Asian Pac. J. Cancer Prev. 2014, 15, 5001–5007. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, A.; Sarkar, S.H.; Bitar, B.; Ali, S.; Aboukameel, A.; Sethi, S.; Li, Y.; Bao, B.; Kong, D.; Banerjee, S.; et al. Garcinol regulates EMT and Wnt signaling pathways in vitro and in vivo, leading to anticancer activity against breast cancer cells. Mol. Cancer Ther. 2012, 11, 2193–2201. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, A.; Wang, Z.; Ali, R.; Maitah, M.Y.; Kong, D.; Banerjee, S.; Padhye, S.; Sarkar, F.H. Apoptosis-inducing effect of garcinol is mediated by NF-kappaB signaling in breast cancer cells. J. Cell Biochem. 2010, 109, 1134–1141. [Google Scholar]
- Huang, W.C.; Kuo, K.T.; Adebayo, B.O.; Wang, C.H.; Chen, Y.J.; Jin, K.; Tsai, T.H.; Yeh, C.T. Garcinol inhibits cancer stem cell-like phenotype via suppression of the Wnt/beta-catenin/STAT3 axis signalling pathway in human non-small cell lung carcinomas. J. Nutr. Biochem. 2018, 54, 140–150. [Google Scholar] [CrossRef]
- Yu, S.Y.; Liao, C.H.; Chien, M.H.; Tsai, T.Y.; Lin, J.K.; Weng, M.S. Induction of p21(Waf1/Cip1) by garcinol via downregulation of p38-MAPK signaling in p53-independent H1299 lung cancer. J. Agric. Food Chem. 2014, 62, 2085–2095. [Google Scholar] [CrossRef]
- Ranjbarnejad, T.; Saidijam, M.; Tafakh, M.S.; Pourjafar, M.; Talebzadeh, F.; Najafi, R. Garcinol exhibits anti-proliferative activities by targeting microsomal prostaglandin E synthase-1 in human colon cancer cells. Hum. Exp. Toxicol. 2017, 36, 692–700. [Google Scholar] [CrossRef]
- Hong, J.; Kwon, S.J.; Sang, S.; Ju, J.; Zhou, J.N.; Ho, C.T.; Huang, M.T.; Yang, C.S. Effects of garcinol and its derivatives on intestinal cell growth: Inhibitory effects and autoxidation-dependent growth-stimulatory effects. Free. Radic. Biol. Med. 2007, 42, 1211–1221. [Google Scholar] [CrossRef]
- Liao, C.H.; Sang, S.; Ho, C.T.; Lin, J.K. Garcinol modulates tyrosine phosphorylation of FAK and subsequently induces apoptosis through down-regulation of Src, ERK, and Akt survival signaling in human colon cancer cells. J. Cell Biochem. 2005, 96, 155–169. [Google Scholar] [CrossRef]
- Pieme, C.A.; Ambassa, P.; Yankep, E.; Saxena, A.K. Epigarcinol and isogarcinol isolated from the root of Garcinia ovalifolia induce apoptosis of human promyelocytic leukemia (HL-60 cells). BMC Res. Notes 2015, 8, 700. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, K.; Akao, Y.; Kobayashi, E.; Ito, T.; Ohguchi, K.; Tanaka, T.; Iinuma, M.; Nozawa, Y. Cytotoxic benzophenone derivatives from Garcinia species display a strong apoptosis-inducing effect against human leukemia cell lines. Biol. Pharm. Bull. 2003, 26, 569–571. [Google Scholar] [CrossRef] [Green Version]
- Pan, M.H.; Chang, W.L.; Lin-Shiau, S.Y.; Ho, C.T.; Lin, J.K. Induction of apoptosis by garcinol and curcumin through cytochrome c release and activation of caspases in human leukemia HL-60 cells. J. Agric. Food Chem. 2001, 49, 1464–1474. [Google Scholar] [CrossRef]
- Wang, Y.; Tsai, M.L.; Chiou, L.Y.; Ho, C.T.; Pan, M.H. Antitumor Activity of Garcinol in Human Prostate Cancer Cells and Xenograft Mice. J. Agric. Food Chem. 2015, 63, 9047–9052. [Google Scholar] [CrossRef]
- Ahmad, A.; Wang, Z.; Wojewoda, C.; Ali, R.; Kong, D.; Maitah, M.Y.; Banerjee, S.; Bao, B.; Padhye, S.; Sarkar, F.H. Garcinol-induced apoptosis in prostate and pancreatic cancer cells is mediated by NF- kappaB signaling. Front. Biosci. 2011, 3, 1483–1492. [Google Scholar] [CrossRef]
- Sethi, G.; Chatterjee, S.; Rajendran, P.; Li, F.; Shanmugam, M.K.; Wong, K.F.; Kumar, A.P.; Senapati, P.; Behera, A.K.; Hui, K.M.; et al. Inhibition of STAT3 dimerization and acetylation by garcinol suppresses the growth of human hepatocellular carcinoma in vitro and in vivo. Mol. Cancer 2014, 13, 66. [Google Scholar] [CrossRef] [Green Version]
- Cheng, A.C.; Tsai, M.L.; Liu, C.M.; Lee, M.F.; Nagabhushanam, K.; Ho, C.T.; Pan, M.H. Garcinol inhibits cell growth in hepatocellular carcinoma Hep3B cells through induction of ROS-dependent apoptosis. Food Funct. 2010, 1, 301–307. [Google Scholar] [CrossRef]
- Ahn, K.S.; Sethi, G.; Jain, A.K.; Jaiswal, A.K.; Aggarwal, B.B. Genetic deletion of NAD(P)H:quinone oxidoreductase 1 abrogates activation of nuclear factor-kappaB, IkappaBalpha kinase, c-Jun N-terminal kinase, Akt, p38, and p44/42 mitogen-activated protein kinases and potentiates apoptosis. J. Biol. Chem. 2006, 281, 19798–19808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawhney, M.; Rohatgi, N.; Kaur, J.; Shishodia, S.; Sethi, G.; Gupta, S.D.; Deo, S.V.; Shukla, N.K.; Aggarwal, B.B.; Ralhan, R. Expression of NF-kappaB parallels COX-2 expression in oral precancer and cancer: Association with smokeless tobacco. Int. J. Cancer 2007, 120, 2545–2556. [Google Scholar] [CrossRef] [PubMed]
- Sethi, G.; Ahn, K.S.; Sung, B.; Aggarwal, B.B. Pinitol targets nuclear factor-kappaB activation pathway leading to inhibition of gene products associated with proliferation, apoptosis, invasion, and angiogenesis. Mol. Cancer Ther. 2008, 7, 1604–1614. [Google Scholar] [CrossRef] [Green Version]
- Sethi, G.; Sung, B.; Kunnumakkara, A.B.; Aggarwal, B.B. Targeting TNF for Treatment of Cancer and Autoimmunity. Adv. Exp. Med. Biol. 2009, 647, 37–51. [Google Scholar]
- Shin, E.M.; Hay, H.S.; Lee, M.H.; Goh, J.N.; Tan, T.Z.; Sen, Y.P.; Lim, S.W.; Yousef, E.M.; Ong, H.T.; Thike, A.A.; et al. DEAD-box helicase DP103 defines metastatic potential of human breast cancers. J. Clin. Investig. 2014, 124, 3807–3824. [Google Scholar] [CrossRef]
- Ghate, N.B.; Chaudhuri, D.; Das, A.; Panja, S.; Mandal, N. An Antioxidant Extract of the Insectivorous Plant Drosera burmannii Vahl. Alleviates Iron-Induced Oxidative Stress and Hepatic Injury in Mice. PLoS ONE 2015, 10, e0128221. [Google Scholar] [CrossRef]
- Zhang, J.; Ahn, K.S.; Kim, C.; Shanmugam, M.K.; Siveen, K.S.; Arfuso, F.; Samym, R.P.; Deivasigamanim, A.; Lim, L.H.; Wang, L.; et al. Nimbolide-Induced Oxidative Stress Abrogates STAT3 Signaling Cascade and Inhibits Tumor Growth in Transgenic Adenocarcinoma of Mouse Prostate Model. Antioxid. Redox. Signal. 2016, 24, 575–589. [Google Scholar] [CrossRef]
- Shaikh, R.; Pund, M.; Dawane, A.; Iliyas, S. Evaluation of Anticancer, Antioxidant, and Possible Anti-inflammatory Properties of Selected Medicinal Plants Used in Indian Traditional Medication. J. Tradit. Complement Med. 2014, 4, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Woo, C.C.; Hsu, A.; Kumar, A.P.; Sethi, G.; Tan, K.H. Thymoquinone inhibits tumor growth and induces apoptosis in a breast cancer xenograft mouse model: The role of p38 MAPK and ROS. PLoS ONE 2013, 8, e75356. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Lee, J.H.; Kim, S.H.; Sethi, G.; Ahn, K.S. Artesunate suppresses tumor growth and induces apoptosis through the modulation of multiple oncogenic cascades in a chronic myeloid leukemia xenograft mouse model. Oncotarget 2015, 6, 4020–4035. [Google Scholar] [CrossRef]
- Liu, J.; Wan, J. Rationale for the use of natural anti-inflammatory agents in cancer chemotherapy. N. Am. J. Med. Sci. 2010, 3, 160–166. [Google Scholar] [CrossRef]
- Chai, E.Z.; Siveen, K.S.; Shanmugam, M.K.; Arfuso, F.; Sethi, G. Analysis of the intricate relationship between chronic inflammation and cancer. Biochem. J. 2015, 468, 1–15. [Google Scholar] [CrossRef]
- Rayburn, E.R.; Ezell, S.J.; Zhang, R. Anti-Inflammatory Agents for Cancer Therapy. Mol. Cell Pharmacol. 2009, 1, 29–43. [Google Scholar] [CrossRef]
- Chua, A.W.; Hay, H.S.; Rajendran, P.; Shanmugam, M.K.; Li, F.; Bist, P.; Koay, E.S.; Lim, L.H.; Kumar, A.P.; Sethi, G. Butein downregulates chemokine receptor CXCR4 expression and function through suppression of NF-kappaB activation in breast and pancreatic tumor cells. Biochem. Pharmacol. 2010, 80, 1553–1562. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Lee, J.H.; Chai, E.Z.; Kanchi, M.M.; Kar, S.; Arfuso, F.; Dharmarajan, A.; Kumar, A.P.; Ramar, P.S.; Looi, C.Y.; et al. Cancer prevention and therapy through the modulation of transcription factors by bioactive natural compounds. Semin. Cancer Biol. 2016, 40, 35–47. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Manu, K.A.; Ong, T.H.; Ramachandran, L.; Surana, R.; Bist, P.; Lim, L.H.; Kumar, A.P.; Hui, K.M.; Sethi, G. Inhibition of CXCR4/CXCL12 signaling axis by ursolic acid leads to suppression of metastasis in transgenic adenocarcinoma of mouse prostate model. Int. J. Cancer 2011, 129, 1552–1563. [Google Scholar] [CrossRef]
- Siveen, K.S.; Mustafa, N.; Li, F.; Kannaiyan, R.; Ahn, K.S.; Kumar, A.P.; Chng, W.J.; Sethi, G. Thymoquinone overcomes chemoresistance and enhances the anticancer effects of bortezomib through abrogation of NF-kappaB regulated gene products in multiple myeloma xenograft mouse model. Oncotarget 2014, 5, 634–648. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, R.A.; McLaughlin, R.J. Garcinol: A Magic Bullet of Amnesia for Maladaptive Memories? Neuropsychopharmacology 2017, 42, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Biersack, B. Effects of Garcinol from Kokum (Garcinia indica) on the Prevention and Treatment of Cancer. In Critical Dietary Factors in Cancer Chemoprevention; Springer: Berlin/Heidelberg, Germany, 2016; pp. 253–271. [Google Scholar]
- Sang, S.; Liao, C.-H.; Pan, M.-H.; Rosen, R.T.; Lin-Shiau, S.-Y.; Lin, J.-K.; Ho, C.-T. Chemical studies on antioxidant mechanism of garcinol: Analysis of radical reaction products of garcinol with peroxyl radicals and their antitumor activities. Tetrahedron 2002, 58, 10095–10102. [Google Scholar] [CrossRef]
- Sang, S.; Pan, M.-H.; Cheng, X.; Bai, N.; Stark, R.E.; Rosen, R.T.; Lin-Shiau, S.-Y.; Lin, J.-K.; Ho, C.-T. Chemical studies on antioxidant mechanism of garcinol: Analysis of radical reaction products of garcinol and their antitumor activities. Tetrahedron 2001, 57, 9931–9938. [Google Scholar] [CrossRef]
- Tanaka, T.; Kohno, H.; Shimada, R.; Kagami, S.; Yamaguchi, F.; Kataoka, S.; Ariga, T.; Murakami, A.; Koshimizu, K.; Ohigashi, H. Prevention of colonic aberrant crypt foci by dietary feeding of garcinol in male F344 rats. Carcinogenesis 2000, 21, 1183–1189. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Sang, S.; Park, H.J.; Kwon, S.J.; Suh, N.; Huang, M.T.; Ho, C.T.; Yang, C.S. Modulation of arachidonic acid metabolism and nitric oxide synthesis by garcinol and its derivatives. Carcinogenesis 2006, 27, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Paul, B.; Gaonkar, R.H.; Mukhopadhyay, R.; Ganguly, S.; Debnath, M.C.; Mukherjee, B. Garcinol-loaded novel cationic nanoliposomes: In vitro and in vivo study against B16F10 melanoma tumor model. Nanomedicine 2019, 14, 2045–2065. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, F.; Saito, M.; Ariga, T.; Yoshimura, Y.; Nakazawa, H. Free radical scavenging activity and antiulcer activity of garcinol from Garcinia indica fruit rind. J. Agric. Food Chem. 2000, 48, 2320–2325. [Google Scholar] [CrossRef]
- Yamaguchi, F.; Ariga, T.; Yoshimura, Y.; Nakazawa, H. Antioxidative and anti-glycation activity of garcinol from Garcinia indica fruit rind. J. Agric. Food Chem. 2000, 48, 180–185. [Google Scholar] [CrossRef]
- Kolodziejczyk, J.; Masullo, M.; Olas, B.; Piacente, S.; Wachowicz, B. Effects of garcinol and guttiferone K isolated from Garcinia cambogia on oxidative/nitrative modifications in blood platelets and plasma. Platelets 2009, 20, 487–492. [Google Scholar] [CrossRef]
- Liao, C.H.; Sang, S.; Liang, Y.C.; Ho, C.T.; Lin, J.K. Suppression of inducible nitric oxide synthase and cyclooxygenase-2 in downregulating nuclear factor-kappa B pathway by Garcinol. Mol. Carcinog. 2004, 41, 140–149. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, T.B.; Moon, K.A.; Kim, T.J.; Shin, D.; Cho, Y.S.; Moon, H.B.; Lee, K.Y. Regulation of pro-inflammatory responses by lipoxygenases via intracellular reactive oxygen species in vitro and in vivo. Exp. Mol. Med. 2008, 40, 461–476. [Google Scholar] [CrossRef] [Green Version]
- Koeberle, A.; Northoff, H.; Werz, O. Identification of 5-lipoxygenase and microsomal prostaglandin E2 synthase-1 as functional targets of the anti-inflammatory and anti-carcinogenic garcinol. Biochem. Pharmacol. 2009, 77, 1513–1521. [Google Scholar] [CrossRef] [Green Version]
- Parasramka, M.A.; Gupta, S.V. Garcinol inhibits cell proliferation and promotes apoptosis in pancreatic adenocarcinoma cells. Nutr. Cancer 2011, 63, 456–465. [Google Scholar] [CrossRef]
- Tsai, M.L.; Chiou, Y.S.; Chiou, L.Y.; Ho, C.T.; Pan, M.H. Garcinol suppresses inflammation-associated colon carcinogenesis in mice. Mol. Nutr. Food Res. 2014, 58, 1820–1829. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zhou, H.; Wang, M.; Cen, J.; Wei, Q. Immune regulation and anti-inflammatory effects of isogarcinol extracted from Garcinia mangostana L. against collagen-induced arthritis. J. Agric. Food Chem. 2014, 62, 4127–4134. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.K.; Warrier, S.; Kumar, A.P.; Sethi, G.; Arfuso, F. Potential Role of Natural Compounds as Anti-Angiogenic Agents in Cancer. Curr. Vasc. Pharmacol. 2017, 15, 503–519. [Google Scholar] [CrossRef] [PubMed]
- Siveen, K.S.; Ahn, K.S.; Ong, T.H.; Shanmugam, M.K.; Li, F.; Yap, W.N.; Kumar, A.P.; Fong, C.W.; Tergaonkar, V.; Hui, K.M.; et al. Y-tocotrienol inhibits angiogenesis-dependent growth of human hepatocellular carcinoma through abrogation of AKT/mTOR pathway in an orthotopic mouse model. Oncotarget 2014, 5, 1897–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, S.M.; Li, F.; Rajendran, P.; Kumar, A.P.; Hui, K.M.; Sethi, G. Identification of beta-escin as a novel inhibitor of signal transducer and activator of transcription 3/Janus-activated kinase 2 signaling pathway that suppresses proliferation and induces apoptosis in human hepatocellular carcinoma cells. J. Pharmacol. Exp. Ther. 2010, 334, 285–293. [Google Scholar] [CrossRef]
- Wong, A.L.A.; Hirpara, J.L.; Pervaiz, S.; Eu, J.Q.; Sethi, G.; Goh, B.C. Do STAT3 inhibitors have potential in the future for cancer therapy? Expert Opin. Investig. Drugs 2017, 26, 883–887. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Hirpara, J.L.; Eu, J.Q.; Sethi, G.; Wang, L.; Goh, B.C.; Wong, A.L. Targeting STAT3 and oxidative phosphorylation in oncogene-addicted tumors. Redox Biol. 2019, 25, 101073. [Google Scholar] [CrossRef]
- Mohan, C.D.; Bharathkumar, H.; Bulusu, K.C.; Pandey, V.; Rangappa, S.; Fuchs, J.E.; Shanmugam, M.K.; Dai, X.; Li, F.; Deivasigamani, A.; et al. Development of a novel azaspirane that targets the Janus kinase-signal transducer and activator of transcription (STAT) pathway in hepatocellular carcinoma in vitro and in vivo. J. Biol. Chem. 2014, 289, 34296–34307. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, P.; Li, F.; Shanmugam, M.K.; Vali, S.; Abbasi, T.; Kapoor, S.; Ahn, K.S.; Kumar, A.P.; Sethi, G. Honokiol inhibits signal transducer and activator of transcription-3 signaling, proliferation, and survival of hepatocellular carcinoma cells via the protein tyrosine phosphatase SHP-1. J. Cell. Physiol. 2012, 227, 2184–2195. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, C.; Sethi, G.; Ahn, K.S. Brassinin inhibits STAT3 signaling pathway through modulation of PIAS-3 and SOCS-3 expression and sensitizes human lung cancer xenograft in nude mice to paclitaxel. Oncotarget 2015, 6, 6386–6405. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Shanmugam, M.K.; Chen, L.; Chatterjee, S.; Basha, J.; Kumar, A.P.; Kundu, T.K.; Sethi, G. Garcinol, a polyisoprenylated benzophenone modulates multiple proinflammatory signaling cascades leading to the suppression of growth and survival of head and neck carcinoma. Cancer Prev. Res. 2013, 6, 843–854. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.T.; Yang, X.A.; Fang, L.Y.; Wang, J.H.; Liu, Q. Anti-proliferative and anti-invasive effects of garcinol from Garcinia indica on gallbladder carcinoma cells. Pharmazie. 2018, 73, 413–417. [Google Scholar] [PubMed]
- Wang, J.; Wu, M.; Zheng, D.; Zhang, H.; Lv, Y.; Zhang, L.; Tan, H.S.; Zhou, H.; Lao, Y.Z.; Xu, H.X. Garcinol inhibits esophageal cancer metastasis by suppressing the p300 and TGF-beta1 signaling pathways. Acta Pharmacol. Sin. 2020, 41, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.W.; Lee, P.M.; Bamodu, O.A.; Su, Y.K.; Fong, I.H.; Yeh, C.T.; Chien, M.H.; Kan, I.H.; Lin, C.M. Enhanced Hsa-miR-181d/p-STAT3 and Hsa-miR-181d/p-STAT5A Ratios Mediate the Anticancer Effect of Garcinol in STAT3/5A-Addicted Glioblastoma. Cancers 2019, 11, 1888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.S.; Lee, C.H.; Hsieh, C.D.; Ho, C.T.; Pan, M.H.; Huang, C.S.; Tu, S.H.; Wang, Y.J.; Chen, L.C.; Chang, Y.J.; et al. Nicotine-induced human breast cancer cell proliferation attenuated by garcinol through down-regulation of the nicotinic receptor and cyclin D3 proteins. Breast Cancer Res. Treat. 2011, 125, 73–87. [Google Scholar] [CrossRef]
- Collins, H.M.; Abdelghany, M.K.; Messmer, M.; Yue, B.; Deeves, S.E.; Kindle, K.B.; Mantelingu, K.; Aslam, A.; Winkler, G.S.; Kundu, T.K.; et al. Differential effects of garcinol and curcumin on histone and p53 modifications in tumour cells. BMC Cancer 2013, 13, 37. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, A.; Sarkar, S.H.; Aboukameel, A.; Ali, S.; Biersack, B.; Seibt, S.; Li, Y.; Bao, B.; Kong, D.; Banerjee, S.; et al. Anticancer action of garcinol in vitro and in vivo is in part mediated through inhibition of STAT-3 signaling. Carcinogenesis 2012, 33, 2450–2456. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, L.; Ho, C.T.; Zhang, K.; Liu, Q.; Zhao, H. Garcinol from Garcinia indica Downregulates Cancer Stem-like Cell Biomarker ALDH1A1 in Nonsmall Cell Lung Cancer A549 Cells through DDIT3 Activation. J. Agric. Food Chem. 2017, 65, 3675–3683. [Google Scholar] [CrossRef]
- Ohnishi, H.; Asamoto, M.; Tujimura, K.; Hokaiwado, N.; Takahashi, S.; Ogawa, K.; Kuribayashi, M.; Ogiso, T.; Okuyama, H.; Shirai, T. Inhibition of cell proliferation by nobiletin, a dietary phytochemical, associated with apoptosis and characteristic gene expression, but lack of effect on early rat hepatocarcinogenesis in vivo. Cancer Sci. 2004, 95, 936–942. [Google Scholar] [CrossRef]
- Huang, C.C.; Lin, C.M.; Huang, Y.J.; Wei, L.; Ting, L.L.; Kuo, C.C.; Hsu, C.; Chiou, J.F.; Wu, A.T.H.; Lee, W.H. Garcinol downregulates Notch1 signaling via modulating miR-200c and suppresses oncogenic properties of PANC-1 cancer stem-like cells. Biotechnol. Appl. Biochem. 2017, 64, 165–173. [Google Scholar] [CrossRef]
- Parasramka, M.A.; Ali, S.; Banerjee, S.; Deryavoush, T.; Sarkar, F.H.; Gupta, S. Garcinol sensitizes human pancreatic adenocarcinoma cells to gemcitabine in association with microRNA signatures. Mol. Nutr. Food Res. 2013, 57, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Tanaka, T.; Hirose, Y.; Yamaguchi, F.; Kohno, H.; Toida, M.; Hara, A.; Sugie, S.; Shibata, T.; Mori, H. Dietary garcinol inhibits 4-nitroquinoline 1-oxide-induced tongue carcinogenesis in rats. Cancer Lett. 2005, 221, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Saadat, N.; Akhtar, S.; Goja, A.; Razalli, N.H.; Geamanu, A.; David, D.; Shen, Y.; Gupta, S.V. Dietary Garcinol Arrests Pancreatic Cancer in p53 and K-ras Conditional Mutant Mouse Model. Nutr. Cancer 2018, 70, 1075–1087. [Google Scholar] [CrossRef] [PubMed]
- Tuli, H.S.; Kashyap, D.; Sharma, A.K.; Sandhu, S.S. Molecular aspects of melatonin (MLT)-mediated therapeutic effects. Life Sci. 2015, 135, 147–157. [Google Scholar] [CrossRef]
- Narasimhan, S.; Maheshwaran, S.; Abu-Yousef, I.A.; Majdalawieh, A.F.; Rethavathi, J.; Das, P.E.; Poltronieri, P. Anti-Bacterial and Anti-Fungal Activity of Xanthones Obtained via Semi-Synthetic Modification of alpha-Mangostin from Garcinia mangostana. Molecules 2017, 22, 275. [Google Scholar] [CrossRef] [Green Version]
- Warriar, P.; Barve, K.; Prabhakar, B. Anti-Arthritic Effect of Garcinol Enriched Fraction Against Adjuvant Induced Arthritis. Recent Pat. Inflamm. Allergy Drug Discov. 2019, 13, 49–56. [Google Scholar] [CrossRef]
- Dhamija, I.; Parle, M.; Kumar, S. Antidepressant and anxiolytic effects of Garcinia indica fruit rind via monoaminergic pathway. 3 Biotech. 2017, 7, 131. [Google Scholar] [CrossRef] [Green Version]
- Parasramka, M.A.; Gupta, S.V. Synergistic effect of garcinol and curcumin on antiproliferative and apoptotic activity in pancreatic cancer cells. J. Oncol. 2012, 2012, 709739. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Seo, S.U.; Min, K.J.; Woo, S.M.; Nam, J.O.; Kubatka, P.; Kim, S.; Park, J.W.; Kwon, T.K. Garcinol Enhances TRAIL-Induced Apoptotic Cell Death through Up-Regulation of DR5 and Down-Regulation of c-FLIP Expression. Molecules 2018, 23, 1614. [Google Scholar] [CrossRef] [Green Version]
- Boisgard, A.S.; Lamrayah, M.; Dzikowski, M.; Salmon, D.; Kirilov, P.; Primard, C.; Pirot, F.; Fromy, B.; Verrier, B. Innovative drug vehicle for local treatment of inflammatory skin diseases: Ex vivo and in vivo screening of five topical formulations containing poly(lactic acid) (PLA) nanoparticles. Eur. J. Pharm. Biopharm. 2017, 116, 51–60. [Google Scholar] [CrossRef]
- Zhang, M.; Viennois, E.; Prasad, M.; Zhang, Y.; Wang, L.; Zhang, Z.; Han, M.K.; Xiao, B.; Xu, C.; Srinivasan, S.; et al. Edible ginger-derived nanoparticles: A novel therapeutic approach for the prevention and treatment of inflammatory bowel disease and colitis-associated cancer. Biomaterials 2016, 101, 321–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaonkar, R.H.; Ganguly, S.; Dewanjee, S.; Sinha, S.; Gupta, A.; Ganguly, S.; Chattopadhyay, D.; Chatterjee Debnath, M. Garcinol loaded vitamin E TPGS emulsified PLGA nanoparticles: Preparation, physicochemical characterization, in vitro and in vivo studies. Sci. Rep. 2017, 7, 530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamaly, N.; Yameen, B.; Wu, J.; Farokhzad, O.C. Degradable Controlled-Release Polymers and Polymeric Nanoparticles: Mechanisms of Controlling Drug Release. Chem. Rev. 2016, 116, 2602–2663. [Google Scholar] [CrossRef] [Green Version]
- Collignon, P.; Powers, J.H.; Chiller, T.M.; Aidara-Kane, A.; Aarestrup, F.M. World Health Organization ranking of antimicrobials according to their importance in human medicine: A critical step for developing risk management strategies for the use of antimicrobials in food production animals. Clin. Infect. Dis. 2009, 49, 132–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senarathna, U.L.; Fernando, S.S.; Gunasekara, T.D.; Weerasekera, M.M.; Hewageegana, H.G.; Arachchi, N.D.; Siriwardena, H.D.; Jayaweera, P.M. Enhanced antibacterial activity of TiO2 nanoparticle surface modified with Garcinia zeylanica extract. Chem. Cent. J. 2017, 11, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danhier, F.; Ansorena, E.; Silva, J.M.; Coco, R.; Le Breton, A.; Preat, V. PLGA-based nanoparticles: An overview of biomedical applications. J. Control Release 2012, 161, 505–522. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; He, B.; Liu, L.; Qu, G.; Shi, J.; Hu, L.; Jiang, G. Antibacterial mechanism of silver nanoparticles in Pseudomonas aeruginosa: Proteomics approach. Metallomics 2018, 10, 557–564. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Cancer | Cell Lines | Effects | Mechanisms | Concentration | Ref. |
|---|---|---|---|---|---|
| Skin (Melanoma) | B16F10 cells | Inhibited cell proliferation | ↑ Apoptosis, ↑ Caspase-3, ↑ Caspase-9, ↑ Bax, ↓ Bcl-2, | 10–50 μg/mL | [85] |
| Head | CAL27 | Reduction of cell viability | ↑ Apoptosis, ↓ STAT3, ↓ c-Src, JAK1/2, ↓ NF-κB, ↓ TAK1 | 0–50 µM | [103] |
| Brain (Glioblastoma) | U-87 MG and GBM8401 GBM | Inhibited cell viability | ↑ Apoptosis, ↑ STAT3 and ↑ STAT5A, ↑ hsa-miR-181d/STAT3 and hsa-miR-181d/5A ratio, ↑ Bax; ↓ Bcl-2 | 2.5–40 µM | [106] |
| Cervical | Hela and SiHa | Suppressed cell viability, invasion, and migration | ↑ Apoptosis ↑ T-cadherin knockdown of T-cadherin, ↑ P13K/AKT signaling pathway | 0, 5, 10, 25 μM | [42] |
| Oral squamous cell carcinoma | SCC-4, SCC-9 and SCC-25 | Inhibits tumor cell proliferation, cell cycle progression, and angiogenesis | ↓ NF-κB ↓ COX-2 ↓ VEGF | 0 to 25 µM | [40] |
| Cal 27 | Inhibit cell proliferation | ↑ Apoptosis | 0–20 µM | [41] | |
| Esophageal | KYSE150 and KYSE450 | Inhibits cancer metastasis | ↓ p300 and TGF-β1, ↓ p300/CBP, ↓ p-Smad2/3 | 0, 5, 10, 20 μM | [105] |
| Breast | MCF-7, MDA-MB-231, AU-565 | Inhibition of cell proliferation | ↓ cyclin D3 expression ↓ a9-nAChR expression | 0 to 20 μM | [107] |
| MCF7 | Suppressed cell proliferation | Inhibit histone acetyltransferase activities, ↓ acetylation of p53, ↑ DNA damage signaling and the induction of chromatin regulators such as TIP60 and SUV420H2 | 0, 10, 20 μM | [108] | |
| MDA-MB-231 and BT-549 | Inhibit cell proliferation | ↑ E-cadherin, ↓ vimentin, ↓ ZEB-1, ↓ ZEB-2, ↑ miR-200, ↑ let-7 family microRNAs | 0 to 25 µM | [48] | |
| MDA-MB-231 | Inhibit cell proliferation | ↑ Apoptosis, ↑ STAT3, ↓ total and p-STAT-3, ↓ IL-6-induced STAT-3 phosphorylation, ↓ u-Pa, ↓ VEGF and MMP-9 | 0, 10, 25 µM | [109] | |
| MCF-7 | Inhibited cell proliferation, inhibited cell cycle progression | ↑ Apoptosis, Cell cycle arrest at G0/G1 phase, ↑ ac-H4, ↑ ac-H3, ↑ NF-κB/ac-p65, ↓ ac-p65 in NF-κB pathway, ↓ cyclin D1, ↓ Bcl-xl, ↓ Bcl-2 | 10–50 µM | [47] | |
| MCF7, MDAMB231 and SKBR3 | Anti- proliferative responses | ↑ Apoptosis, ↑ P53 ↑ Bax ↓ Bcl-XL, ↓ Nitrite and TNF-α level | 1.56–25 μg/mL | [43] | |
| Leukemia | HL-60 cells | Cancer chemo preventive effect | ↑ Apoptosis, ↑ caspase-3/CPP32 activity, ↑ degradation of PARP, ↓ Bcl-2, ↑ Bad, ↑ Bax | IC50 values of 9.42 µM and 19.5 µM | [57] |
| NB4, HL60, U937, and K562 | Growth inhibitory effects | ↑ Apoptosis, ↑caspase 3, ↓ mitochondrial transmembrane potential | 0–20 µM | [56] | |
| Lung | H441 and A549 NSCLC cell lines | Inhibits cancer cell | ↓ Wnt/β-catenin /STAT3, Impaired phosphorylation of LRP6, ↓Axin2, ↓ β-catenin, Dvl2, ↓ cyclin D1, | 0~20 μM | [50] |
| A549 and H1299 | Anti- proliferative responses | ↑ Apoptosis, ↑ miRNAs (miR-200b, miR-205, miR-218, and let-7c) | 0–20 µM | [20] | |
| A549, H460, H1299, H1650, H358, and HCC827 | Inhibited cellproliferation/suppressed cellviability | ↓ C/EBPβ, ↓ ALDH1A1, ↑ DDIT3 | 0.1% (v/v) | [110] | |
| Hepatocellular | A431, Hep3B cells | Decreasescell viability | ↑ Apoptosis, ↓ CEBP, ↑ GADD153, ↑ ROS, ↓ mitochondrial membrane potential, ↑ Bax/Bcl-2 ratio, ↑ Caspase-8, ↑ tBid, ↑ caspase-3, ↑caspase-9 | 0–50 µM | [61] |
| MH1C1 and HepG2 | Inhibition of cell proliferation | ↑ Apoptosis, ↓ cyclin E, ↓ cyclin B, ↓ bcl-2 | 500 ppm | [111] | |
| C3A, HepG2, PLC/PRF5, and HUH-7 | Inhibition of cell proliferation | ↑ Apoptosis, ↓ IL-6, ↓ STAT3 acetylation, ↓ cyclin D1, ↓ Bcl-2, ↓ Bcl-xL, ↓ survivin, ↓ Mcl-1, ↓ VEGF | 10 µM | [60] | |
| Gastric | RAW264.7 macrophage cell lines | Chemoprevention and anti-proliferative responses | ↓ (iNOS) ↓ COX-2, lowers the LPS-induced increase of intracellular ROS | 0–5 µM | [89] |
| Pancreatic | BxPC-3 | Inhibit cell proliferation | ↑ Apoptosis, ↓ STAT-3 signaling pathway, ↓ IL-6, ↓ uPA, ↓ VEGF, ↓ MMP-9 | 0, 10, 25 µM | [109] |
| BxPC-3 | Inhibited cell growth | ↓ NF- κB signaling pathway | 0–25 µM | [59] | |
| PANC-1 | Suppresses oncogenic properties of cells | ↑ ABCG2, ↑ Oct4, ↑ CD44, ↓ Mcl-1, ↓ EZH2, ↓ Gli-1, ↓ Notch1, ↑ tumor suppressor miRNAs, ↑ miR-200c | 0–10 µM | [112] | |
| BxPC-3 and Panc-1 | Inhibited cell proliferation | ↑ Apoptosis, ↑ PARP, VEGF, MMPs, ILs, caspases, and NF-B, ↓ VEGF, ↓ MMP-9, ↓ IL-8 angiogenic factors | 0–20 μM | [113] | |
| Gallbladder | GBC-SD and NOZ | Anti-proliferative | ↓ MMP2, ↓ MMP9, ↓ Stat3 and Akt activation, ↓ mRNA levels of MMP2 and MMP9 | 0–30 µM | [104] |
| Colon | HT-29 | Inhibited cell invasion | ↑ Apoptosis, ↓ Src, ↓ ERK, ↓ Akt, ↓ Bcl-2, ↑ Bax | 10 μM | [54] |
| HT-29 and HCT-116 | Inhibiting growth of cancer cells | ↑ Apoptosis, ↑ p-ERK1/2 | IC50 of 3.2–21.4 μM, | [53] | |
| HT-29 cells | Anti-proliferative activities | ↑ Apoptosis, ↓ HIF-1α, ↓ mPGES-1, ↓ CXCR4, ↓ VEGF, ↑ caspase 3, ↓ MMP-2, ↑ MMP-9, | 0–25 µM | [52] | |
| Prostate | DU145 | Inhibit cell proliferation | ↑ Apoptosis, inhibition of STAT-3 signaling pathway, ↓ p-STAT-3, ↓ IL-6, ↓ uPA, ↓ MMP-9, ↓ VEGF, | 0, 10, 25 µM | [109] |
| LNCaP, C4-2B and PC3 | Inhibited cell growth | ↓ NF-κB signaling pathway | 0–25 µM | [59] |
| Type of Cancer | Animal Models | Effects | Mechanisms | Dose | Duration | Ref. |
|---|---|---|---|---|---|---|
| Skin (Melanoma) | Male Balb/c mice injected subcutaneously with B16F10 tumor | Tumor inhibition | ↑ Metastasis, ↑ Apoptosis, ↑ Bax and ↓ Bcl-2. ↑ Caspase-3, ↑ Caspase-9 | 25 mg/kg | 20 days | [85] |
| Head | Male athymic nu/nu mice inoculated subcutaneously with CAL27 cells | Inhibited tumor growth | ↓ constitutively activated STAT3, ↓ c-Src, JAK1/2, ↓ NF-κB, ↓ TAK1 | 1–2 mg/kg | 4 weeks | [103] |
| Brain (Glioblastoma) | NOD/SCID mice inoculated subcutaneously with U87MG cells | Inhibited tumor growth | ↑ STAT3 and ↑ STAT5A, ↑ hsa-miR-181d/STAT3, ↑ hsa-miR-181d/5A ratio, ↑ Bax, ↓ Bcl-2 | 1 mg/Kg | 4 weeks | [106] |
| Cervical | Male BALB/c nu/nu mice5 inoculated subcutaneously with Hela cells | Inhibited tumor growth | ↑ Apoptosis, ↑ T-cadherin knockdown of T-cahderin, ↑ P13K/AKT signaling pathway | 1 mg/kg and 2 mg/kg | 5 weeks | [42] |
| Oral (tongue) | Male F344 rats | Reduction in cell proliferation | ↓ 4-NQO-induced tongue neoplasms, ↓ BrdU-labeling index, cyclin D1-positive cell ratio | 100 ppm or 500 ppm | 32 weeks | [114] |
| Esophageal | Male BALC/c nude mice Intravenously injected with KYSE150 cells via the tail vein | Reduced tumorincidence | ↓ p300 and TGF-β1 signaling pathways, ↓ protein levels of p300/CBP (transcriptional cofactors and HATs), ↓ p-Smad2/3 expression in the nucleus | 20 mg/kg | 5 weeks | [105] |
| Breast | Female homozygous ICR SCID mice inoculated subcutaneously with MDA-MB-231 | Inhibited tumor growth | ↓ STAT-3 signaling pathway, ↓ p-STAT-3, ↓ IL-6-induced STAT-3 signaling, ↓ VEGF ↓ MMP-9 | 5 mg/day | 4 weeks | [109] |
| Female homozygous ICR SCID mice inoculated subcutaneously with MDA-MB-231 | Inhibited tumor growth | ↓ NF-kB, ↓ miRNAs, vimentin, ↓ β-catenin, miR-200s, ↓ let-7s, ↓ NF-κB, ↓ Wnt signaling pathways. | 5mg/d/animal | 4 weeks | [48] | |
| Male Balb/c mice inoculated with mammary carcinoma 4T1 cells | Antitumor anti-metastasis effects | ↓ caspase-3, ↓ cytosolic Ca2+-iPLA2) ↓ NF-κB, ↓ Twist1 | 1 mg/ kg | 5 weeks | [21] | |
| Lung | Female NMRI (nu/nu) mice inoculated subcutaneously with A549 cells | Inhibited tumor growth | ↓ALDH1A1, ↑DDIT3 | 15mg/kg | 40 day | [110] |
| NOD/SCID mouse bearing H441 tumor sphere | Inhibited tumor growth | ↓ Wnt/β-catenin/STAT3 axis, ↓ p-LRP6, ↓ Axin2, ↓ β-catenin, ↓ cyclin D1 | 5 mg/kg | 9 weeks | [50] | |
| Hepatocellular | Female athymic nu/nu mice inoculated subcutaneously PLC/PRF5 cells | Inhibited tumor growth | ↑ Apoptosis, ↓ IL-6 induced STAT3 activation, ↓ STAT3 acetylation, ↓ cyclin D1, ↓ Bcl-2, ↓Bcl-xL, ↓ survivin, ↓ Mcl-1, ↓VEGF | 1mg/kg and 2 kg/kg | 3 weeks | [60] |
| Pancreatic | Male KPC mice (K-rasLSL.G12D/+; p53R172H/+; PdxCretg/+) | Reduction in tumor volumes | ↓ COX2, ↓ cyclin D1, ↓ VEGF, ↓ Wnt/b-catenin | 0.5 g/kg | 5 weeks | [115] |
| Colon | Male F344 rats | Inhibition of aberrant crypt foci | ↓ PCNA index, ↓ NO and 02, ↓ iNOS, ↓ COX-2 | 15 mg/kg | 5 weeks | [83] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aggarwal, V.; Tuli, H.S.; Kaur, J.; Aggarwal, D.; Parashar, G.; Chaturvedi Parashar, N.; Kulkarni, S.; Kaur, G.; Sak, K.; Kumar, M.; et al. Garcinol Exhibits Anti-Neoplastic Effects by Targeting Diverse Oncogenic Factors in Tumor Cells. Biomedicines 2020, 8, 103. https://doi.org/10.3390/biomedicines8050103
Aggarwal V, Tuli HS, Kaur J, Aggarwal D, Parashar G, Chaturvedi Parashar N, Kulkarni S, Kaur G, Sak K, Kumar M, et al. Garcinol Exhibits Anti-Neoplastic Effects by Targeting Diverse Oncogenic Factors in Tumor Cells. Biomedicines. 2020; 8(5):103. https://doi.org/10.3390/biomedicines8050103
Chicago/Turabian StyleAggarwal, Vaishali, Hardeep Singh Tuli, Jagjit Kaur, Diwakar Aggarwal, Gaurav Parashar, Nidarshana Chaturvedi Parashar, Samruddhi Kulkarni, Ginpreet Kaur, Katrin Sak, Manoj Kumar, and et al. 2020. "Garcinol Exhibits Anti-Neoplastic Effects by Targeting Diverse Oncogenic Factors in Tumor Cells" Biomedicines 8, no. 5: 103. https://doi.org/10.3390/biomedicines8050103
APA StyleAggarwal, V., Tuli, H. S., Kaur, J., Aggarwal, D., Parashar, G., Chaturvedi Parashar, N., Kulkarni, S., Kaur, G., Sak, K., Kumar, M., & Ahn, K. S. (2020). Garcinol Exhibits Anti-Neoplastic Effects by Targeting Diverse Oncogenic Factors in Tumor Cells. Biomedicines, 8(5), 103. https://doi.org/10.3390/biomedicines8050103