Hemocyanins from Helix and Rapana Snails Exhibit in Vitro Antitumor Effects in Human Colorectal Adenocarcinoma

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Isolation of the Hemocyanin and Isoforms from Snail Rapana Venosa
2.3. Isolation of the Native Hemocyanin and Isoforms from Snail H. Lucorum
2.4. Isolation of the Native Hemocyanin and Mucus from Snail H. Aspersa
2.5. Separation of H. Aspersa Hemocyanin Isoforms
2.6. Cell culture and Cell Viability
2.7. Fluorescent Microscopy
2.7.1. Acridine Orange/Ethidium Bromide Double Staining
2.7.2. DAPI Staining
2.8. Transmission Electron Microscopy (TEM)
2.9. Statistical Analysis
3. Results
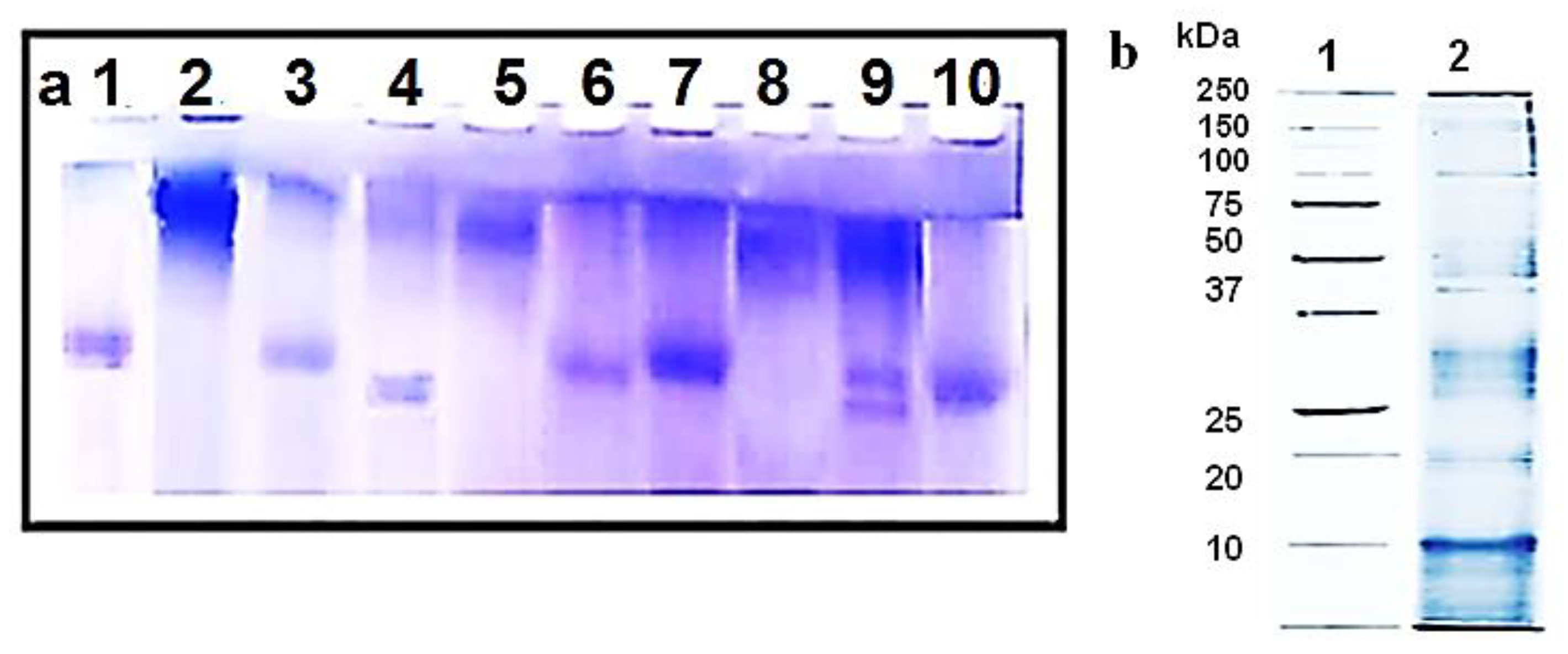
3.1. Isolation of Bioactive Compounds from the Three Mollusk Species
3.2. Effects of the Isolated Bioactive Compounds on the Viability and Proliferative Activity of HT-29 Tumor Cells
3.3. Apotogenic Effects of the Isolated Bioactive Compounds
3.3.1. Vital Double Staining of HT-29 Tumor Cells with Acridine Orange/Ethidium Bromide Acridin
3.3.2. DAPI Staining of HT-29 Tumor Cells
3.4. Transmission Electron Microscopy
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AO | Acridine orange |
| ATCC | 4′,6-Diamidine-2′-phenylindole dihydrochloride |
| DMEM | Dulbecco’s modified Eagle’s medium |
| LD | Linear dichroism |
| DMSO | Dimethyl sulfoxide |
| EB | Ethidium bromide |
| HaH | Helix aspersa hemocyanin |
| HlH | Helix lucorum hemocyanin |
| KLH | Keyhole limpet hemocyanin |
| MTT | Methyl thiazol tetrazolium bromide |
| PBS | Phosphate-buffered saline |
| RvH | Rapana venosa hemocyanin |
| TEM | Transmission electron microscopy |
References
- Falzone, L.; Salomone, S.; Libra, M. Evolution of cancer pharmacological treatments at the turn of the third millennium. Front. Pharmacol. 2018, 9, 1300. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Pezzuto, J.M. Natural products as a vital source for the discovery of cancer chemotherapeutic and chemopreventive agents. Med. Princ. Pract. 2016, 25, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Dong, C.; Li, X.; Han, W.; Su, X. Anticancer potential of bioactive peptides from animal sources. Oncol. Rep. 2017, 38, 637–651. [Google Scholar] [CrossRef] [PubMed]
- Antonova, O.; Toncheva, D.; Rammensee, H.G.; Floetenmeyer, M.; Stevanovic, S.; Dolashka, P. In vitro antiproliferative effect of Helix aspersa hemocyanin on multiple malignant cell lines. Zeitschrift für Naturforschung C 2014, 69, 325–334. [Google Scholar] [CrossRef]
- Becker, M.I.; Arancibia, S.; Salazar, F.; Del Campo, M.; De Ioannes, A. Mollusk Hemocyanins as Natural Immunostimulants in Biomedical Applications, in Immune Response Activation. Duc, G.H.T., Ed.; InTech: Croatia, Rijeka, 2014; pp. 45–72. [Google Scholar]
- Pizarro-Bauerle, J.; Maldonado, I.; Sosoniuk-Roche, E.; Vallejos, G.; López, M.N.; Salazar-Onfray, F.; Aguilar-Guzmán, L.; Valck, C.; Ferreira, A.; Becker, M.I. Molluskan hemocyanins activate the classical pathway of the human complement system through natural antibodies. Front. Immunol. 2017, 8, 188. [Google Scholar] [CrossRef]
- Mora, J.J.; Del Campo, M.; Villar, J.; Paolini, F.; Curzio, G.; Venuti, A.; Jara, L.; Ferreira, J.; Murgas, P.; Lladser, A.; et al. Immunotherapeutic potential of mollusk hemocyanins in combination with human vaccine adjuvants in murine models of oral cancer. J. Immunol. Res. 2019, 2019, 7076942. [Google Scholar]
- Kato, S.; Matsui, T.; Gatsogiannis, C.; Tanaka, Y. Molluscan hemocyanin: Structure, evolution, and physiology. Biophysical Rev. 2018, 10, 191–202. [Google Scholar] [CrossRef]
- Paccagnella, M.; Bologna, L.; Beccaro, M.; Micetic, I.; Di Muro, P.; Salvato, B. Structural subunit organization of molluscan hemocyanins. Micron 2004, 35, 21–22. [Google Scholar] [CrossRef]
- Sandra, K.; Dolashka-Angelova, P.; Devreese, B.; Van Beeumen, J. New insights in Rapana venosa hemocyani N-glycosylation resulting from on-line mass spectrometric analyses. Glycobiology 2007, 17, 141–156. [Google Scholar] [CrossRef]
- Arancibia, S.; Salazar, F.; Becker, M.I. Hemocyanins in the immunotherapy of superficial bladder cancer. In Bladder Cancer-From Basic Science to Robotic Surgery; Canda, A.E., Ed.; InTech: Croatia, Rijeka, 2012; pp. 221–242. [Google Scholar]
- Dolashka, P.; Velkova, L.; Shishkov, S.; Kostova, K.; Dolashki, A.; Dimitrov, I.; Atanasov, B.; Devreese, B.; Voelter, W.; Van Beeumen, J. Glycan structures and antiviral effect of the structural subunit RvH2 of Rapana hemocyanin. Carbohydr. Res. 2010, 345, 2361–2367. [Google Scholar] [CrossRef]
- Zhong, T.-Y.; Arancibia, S.; Born, R.; Tampe, R.; Villar, J.; Del Campo, M.; Manubens, A.; Becker, M.I. Hemocyanins stimulate innate immunity by inducing different temporal patterns of proinflammatory cytokine expression in macrophages. J. Immunol. 2016, 196, 4650–4662. [Google Scholar] [CrossRef]
- Riggs, D.R.; Jackson, B.J.; Vona-Davis, L.; Nigam, A.; McFadden, D.W. In vitro effects of keyhole limpet hemocyanin in breast and pancreatic cancer in regards to cell growth, cytokine production, and apoptosis. Am. J. Surg. 2005, 189, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Dolashka, P.; Velkova, L.; Iliev, I.; Beck, A.; Dolashki, A.; Yossifova, L.; Toshkova, R.; Voelter, W.; Zacharieva, S. Antitumor activity of glycosylated molluscan hemocyanins via Guerin ascites tumor. Immunol. Invest. 2011, 40, 130–149. [Google Scholar] [CrossRef]
- Salazar, M.L.; Jiménez, J.M.; Villar, J.; Rivera, M.; Báez, M.; Manubens, A.; Becker, M. N-Glycosylation of mollusk hemocyanins contributes to their structural stability and immunomodulatory properties in mammals. J. Biol. Chem. 2019, 294, 19546–19564. [Google Scholar] [CrossRef] [PubMed]
- Dolashka-Angelova, P.; Schwarz, H.; Dolashki, A.; Stevanovic, S.; Fecker, M.; Saeed, M.; Voelter, W. Oligomeric stability of Rapana venosa hemocyanin (RvH) and its structural subunits. Biochim. Biophys. Acta. 2003, 1646, 77–85. [Google Scholar] [CrossRef]
- Velkova, L.; Dimitrov, I.; Schwarz, H.; Stevanovic, S.; Voelter, W.; Salvato, B.; Dolashka-Angelova, P. Structure of hemocyanin from garden snail Helix lucorum. Comp. Biochem. Physiol. 2010, 157, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Dolashki, A.; Nissimova, A.; Daskalova, E.; Velkova, L.; Topalova, Y.; Hristova, P.; Traldi, P.; Voelter, W.; Dolashka, P. Structure and antibacterial activity of isolated peptides from the mucus of garden snail Cornu aspersum. Bulg. Chem. Commun. 2018, 50C, 195–200. [Google Scholar]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Schägger, H.; von Jagow, G. Blue Native Electrophoresis for Isolation of Membrane Protein Complexes in Enzymatically Active Form. Anal. Biochem. 1991, 199, 223–231. [Google Scholar] [CrossRef]
- Mossmann, T. Rapid colorimetric assay of cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Wahab, I.; Abdul, A.; Alzubairi, A.; Elhassan, M.; Mohan, S. In vitro ultramorphological assessment of apoptosis induced by zerumbone on (HeLa). J. Biomed. Biotechnol. 2009, 2009, 769568. [Google Scholar] [PubMed]
- Gielens, C.; De Sadeleer, J.; Preaux, G.; Lontie, R. Identification, separation and cheracterization of the hemocyanin components of Helix aspersa. Comp. Biochem. Phys. Part B 1987, 88, 181–186. [Google Scholar] [CrossRef]
- Luo, W.; Yang, G.; Luo, W.; Cao, Z.; Liu, Y.; Qiu, J.; Chen, G.; You, L.; Zhao, F.; Zheng, L.; et al. Novel therapeutic strategies and perspectives for metastatic pancreatic cancer: Vaccine therapy is more than just a theory. Cancer Cell Int. 2020, 20, 66. [Google Scholar] [CrossRef] [PubMed]
- Dolashki, A.; Dolashka, P.; Stenzl, A.; Stevanovic, S.; Aicher, W.K.; Velkova, L.; Velikova, R.; Voelter, W. Antitumour activity of Helix hemocyanin against bladder carcinoma permanent cell lines. Biotech. Biotech. Equip. 2019, 33, 1–13. [Google Scholar] [CrossRef]
- Dolashka-Angelova, P.; Stefanova, T.; Livaniou, E.; Velkova, L.; Klimentzou, P.; Stevanovic, S.; Salvato, B.; Neychev, H.; Voelter, W. Immunological potential of Helix vulgaris and Rapana venosa hemocyanins. Immunol. Invest. 2008, 37, 822–840. [Google Scholar] [CrossRef] [PubMed]
- Miles, D.; Roché, H.; Martin, M.; Perren, T.J.; Cameron, D.A.; Glaspy, J.; Dodwell, D.; Parker, J.; Mayordomo, J.; Tres, A.; et al. Phase III multicenter clinical trial of the sialyl-TN (STn)-keyhole limpet hemocyanin (KLH) vaccine for metastatic breast cancer. Oncologist 2011, 16, 1092–1100. [Google Scholar] [CrossRef]
- Gilewski, T.A.; Ragupathi, G.; Dickler, M.; Powell, S.; Bhuta, S.; Panageas, K.; Koganty, R.R.; Chin-Eng, J.; Hudis, C.; Norton, L.; et al. Immunization of high-risk breast cancer patients with clustered sTn-KLH conjugate plus the immunologic adjuvant QS-21. Clin. Cancer Res. 2007, 13, 2977–2985. [Google Scholar] [CrossRef]
- McFadden, D.W.; Riggs, D.R.; Jackson, B.J.; Vona-Davis, L. Keyhole limpet hemocyanin, a novel immune stimulant with promising anticancer activity in Barrett’s esophageal adenocarcinoma. Am. J. Surg. 2003, 186, 552–555. [Google Scholar] [CrossRef]
- Matusiewicz, M.; Kosieradzka, I.; Niemiec, T.; Grodzik, M.; Antushevich, H.; Strojny, B.; Gołębiewska, M. In Vitro Influence of Extracts from Snail Helix aspersa Müller on the Colon Cancer Cell Line Caco-2. Int. J. Mol. Sci. 2018, 19, 1064. [Google Scholar] [CrossRef]
- Gesheva, V.; Chausheva, S.; Mihaylova, N.; Manoylov, I.; Doumanova, L.; Idakieva, K.; Tchorbanov, A. Anti-cancer properties of gastropodan hemocyanins in murine model of colon carcinoma. BMC Immunol. 2014, 15, 34. [Google Scholar] [CrossRef]
- Stenzl, A.; Dolashki, A.; Stevanovic, S.; Voelter, W.; Aicher, W.; Dolashka, P. Cytotoxic effects of Rapana venosa hemocyanin on bladder cancer permanent cell lines. J. US China Med. Sci. 2016, 13, 79–188. [Google Scholar]
- Antonova, O.; Yossifova, L.; Staneva, R.; Stevanovic, S.; Dolashka, P.; Toncheva, D. Changes in the gene expression profile of the bladder cancer cell lines after treatment with Helix lucorum and Rapana venosa hemocyanin. J. Buon. 2015, 20, 180–187. [Google Scholar] [PubMed]
- Somasundar, P.; Riggs, D.R.; Jackson, B.J.; McFadden, D.W. Inhibition of melanoma growth by hemocyanin occurs via early apoptotic pathways. Am. J. Surg. 2005, 190, 713–716. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 Values (µg/mL) | HT-29 | Balb/c 3T3 |
|---|---|---|
| HaH-total | ›1000 | 934.1 |
| Subunit βc-HaH | 733.8 | ›1000 |
| Subunits α-HaH | 235.3 | 514.6 |
| HlH-total | ›1000 | ›1000 |
| Subunit βc-HlH | ›1000 | ›1000 |
| Subunits α-HlH | ›1000 | ›1000 |
| Subunit RvH I | ›1000 | ›1000 |
| Subunit RvH II | ›1000 | ›1000 |
| Ha-mucus | 415.7 | 825.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Georgieva, A.; Todorova, K.; Iliev, I.; Dilcheva, V.; Vladov, I.; Petkova, S.; Toshkova, R.; Velkova, L.; Dolashki, A.; Dolashka, P. Hemocyanins from Helix and Rapana Snails Exhibit in Vitro Antitumor Effects in Human Colorectal Adenocarcinoma. Biomedicines 2020, 8, 194. https://doi.org/10.3390/biomedicines8070194
Georgieva A, Todorova K, Iliev I, Dilcheva V, Vladov I, Petkova S, Toshkova R, Velkova L, Dolashki A, Dolashka P. Hemocyanins from Helix and Rapana Snails Exhibit in Vitro Antitumor Effects in Human Colorectal Adenocarcinoma. Biomedicines. 2020; 8(7):194. https://doi.org/10.3390/biomedicines8070194
Chicago/Turabian StyleGeorgieva, Ani, Katerina Todorova, Ivan Iliev, Valeriya Dilcheva, Ivelin Vladov, Svetlozara Petkova, Reneta Toshkova, Lyudmila Velkova, Aleksandar Dolashki, and Pavlina Dolashka. 2020. "Hemocyanins from Helix and Rapana Snails Exhibit in Vitro Antitumor Effects in Human Colorectal Adenocarcinoma" Biomedicines 8, no. 7: 194. https://doi.org/10.3390/biomedicines8070194
APA StyleGeorgieva, A., Todorova, K., Iliev, I., Dilcheva, V., Vladov, I., Petkova, S., Toshkova, R., Velkova, L., Dolashki, A., & Dolashka, P. (2020). Hemocyanins from Helix and Rapana Snails Exhibit in Vitro Antitumor Effects in Human Colorectal Adenocarcinoma. Biomedicines, 8(7), 194. https://doi.org/10.3390/biomedicines8070194