1. Introduction
Medicinal plants and their extracts have been used by humans for centuries to feed and treat themselves. The antioxidant and anti-inflammatory properties of botanical extracts are among those with very high potential for the development of new drugs. Plant extracts can have anti-inflammatory and immunoregulatory properties through different active components, in particular antioxidants like flavonoids and carotenoids [
1,
2,
3,
4]. In a recent study, the antioxidant capacity of a spinach extract, constituted of enriched thylakoid membrane extracts and defined as Thykamine has been demonstrated [
5]. Thylakoid membranes composing this extract are found in photosynthetic organisms. They form a network of aggregated membranes attached to unstacked membranes in which galactolipids represent the main lipid constituents. The particular organization of these membranes promotes optimal activity of the two photosystems I and II (PSI and PSII), protein complexes in which photochemical reactions take place and responsible of photosynthesis activity. The antennae complexes of these two photosystems contain carotenoid pigments conferring to thylakoid extracts such as Thykamine a high antioxidant capacity. Moreover, PSI, being rich in Fe-S clusters which are sensitive to reactive oxygen species (ROS)s, are protected by superoxide dismutase (SOD) enzymes located at the external surface of the membrane. SOD reacts with superoxide to form hydrogen peroxide, neutralizing ROS. These antioxidant properties represent powerful weapons against oxidative and nitrosative stresses occurring when there is an imbalance in the production of reactive oxygen species and/or reactive nitrogen species (RNS). Oxidative stress contributes to the development of the inflammatory and immune response of some chronic diseases leading to the cellular dysfunctions and tissue destructions. As a corollary, the different biochemical structure and enzyme activities of Thykamine could be involved in the anti-inflammatory and immunoregulatory functions of thylakoid membranes found in spinach leaves.
The therapeutic effects of Thykamine were previously reported. For example, immuno-modulatory properties of Thykamine on alveolar macrophages were demonstrated through regulation of pro- and anti-inflammatory cytokine production [
6]. Moreover, it was shown that Thykamine contained various antioxidant molecules and enzymes involved in protecting and restoring the harmful effects of UV exposure [
7]. Since neutrophils are the most abundant cellular subset and the first migrating cells accumulating at sites of inflammation with possible deleterious effects, we proposed that Thykamine could affect neutrophils function at these sites.
The inflammatory response to injury implicates an obligatory infiltration of leukocytes in tissue, and among leukocytes, neutrophils are the first migrating cells. Recruitment of neutrophils to sites of inflammation must involve chemotaxis, adhesion of neutrophils to endothelium, trans-endothelial diapedesis, and extravascular migration of neutrophils. From the signals of attraction, neutrophils enter an activation status including the generation of reactive oxygen intermediates and arachidonic acid metabolites, phagocytosis, and the release of proteolytic enzymes. Major chemotactic factors for neutrophils are non-specific products such as formyl-methionyl-peptides, split products of the fifth component of complement, leukotriene B4, and specific attractants such as chemokines (i.e., interleukin 8 (IL-8). The effector functions of neutrophils are fully manifested after their directional influx and resultant local accumulation at sites of tissue reactions. These functions are activated by phagocytosis and soluble stimuli, and depend on both the release of granular contents such as lysosomal hydrolases and the expression of membrane associated enzyme systems such as the superoxide-generating NADPH oxidase. However, besides microbicidal activities and degradation of particles, hydrolytic and oxidative enzymes from neutrophils granules may contribute to deleterious inflammation and damage of host tissues.
Thykamine, being a mixture of various components (thylakoids and galactolipids), corresponds to the “multi-target/multi-component” approach. In this study, the anti-inflammatory activities of Thykamine were investigated in two inflammatory animal models. The antioxidant properties of Thykamine, including superoxide anions (O2−) scavenging and protection against lipid peroxidation and nitric oxide (NO) production were also examined in cell models. We hypothesized that these antioxidant properties combined to the anti-inflammatory activities observed in the animal models could interfere with the inflammatory functions of human blood neutrophils. We studied whether Thykamine could modulate primary functions of freshly isolated human blood neutrophils in vitro, such as chemotaxis, phagocytosis, production of superoxide anions, and degranulation, and how Thykamine could enter the cell.
2. Materials and Methods
2.1. In Vivo Rodent Models of Inflammation
TNBS-induced colitis in rats. Colonic inflammation was induced by using the technique of Morris et al. with slight modifications [
8]. Male Wistar rats (Charles River laboratories, Sherbrooke, QC, Canada) were kept in individuel cages, at 20 °C and 55% relative humidity with 12-h light/12-h dark cycle. The rat experiment was approved by the Animal Experiment committee of the Charles River Laboratories (Montreal, QC Canada) on 15 April 2002 and conducted in accordance with the Canadian Council on Animal Care and Use. The rats were subjected to an 18-h starving period prior to 5 treatments. Briefly, male Wistar rats (180–200 g) after an overnight food deprivation, were anesthetized with isoflurane before insertion of a polyethylene catheter of 8 cm into the colon via the anus. The hapten TNBS at 25 mg/mL dissolved in 50% aqueous/ethanol (vol/vol) was injected into rat colon (total volume injected: 1 mL/rat). The control rats received 1 mL of vehicle (aqueous/ethanol, vol/vol) intracolonically. Rats were injected intraperitoneally with Thykamine (50, 5, or 0.5 mg/kg) in sterile physiologic saline (1 mL) immediately prior to anesthesia. These doses were selected based on previous studies [
6,
7].
Carrageenan-induced paw edema in rats. Male Wistar rats (180–200 g) which had been fasted overnight (18 h) received Thykamine (0.5, 5 or 50 mg/kg) in sterile physiologic saline by intraperitoneal injection (1 mL) immediately prior to subplantar injection of carrageenan (0.1 mL of 1% suspension in 0.9% saline) in the right hind paw [
9]. Paw thickness was measured from ventral to dorsal surfaces, with a dial caliper. Paw circumference was measured immediately prior to carrageenan injection and 5 h afterwards. Edema was expressed in mm as the increase of paw circumference measured after carrageenan injection and compared to the pre-injection value for individual animals.
2.2. Reagents
Dextran T-500 and Ficoll-Paque were obtained from Pharmacia Biotech (Montréal, QC, Canada). fMLP, human LF, anti-human LF antibody and O-dianiside hydrochloride were obtained from Sigma Chemical Co (St-Louis, MO, USA). Chemotaxis plates (96 wells) were purchased from Neuroprobes, Gaithersburg, MD, USA. LTB4 was from Cascade Biochem Ltd. (Reading, Berkshire, England). IL-8 was from Peprotech Canada Inc. (Ottawa, ON, Canada). Fluorescein-labeled Escherichia coli K-12 BioParticles, calcein-AM and cytochrome c (125 mg/mL) were from Molecular Probes, Invitrogen Canada Inc. (Burlington, ON, Canada). Anti-CD63 Fluorescein isothiocyanate (FITC)-labeled and anti-CD66b PE-labeled antibodies were from Beckman Coulter, Inc. (Mississauga, ON, Canada). Thylakoid extracts (Thykamine) were provided by Devonian Health Group (Montmagny, QC, Canada). Thykamine extract were solubilized at 100 mg/mL (5%) in phosphate buffered saline (PBS).
2.3. Neutrophil Preparations
Neutrophils were obtained from venous blood of healthy as previously described [
10]. Contaminating erythrocytes were eliminated by a hypotonic lysis (15 s, RT). After 2 washes, neutrophils were resuspended in Hank’s Balanced Salt solution (HBSS) containing 10 mM HEPES pH 7.4, 1.6 mM Ca
++, and no magnesium. Differential cell counts of leukocytes were performed by flowcytometry (EPICS-XL, Beckman Coulter), Wright’s and non-specific esterase stains. Final neutrophil suspensions were more than 98% pure with no CD3 positive cells and non-specific esterase positive cells represented less than 0.1% of the cell population. Viability was greater than 98% as routinely assessed by trypan blue dye exclusion. When appropriate, neutrophils were preincubated with Thykamine at 37 °C for 30 min before experiments.
2.4. J77A4.1 Cell Culture
J774A.1 monocyte cells (
Mus musculus) (American Type Culture Collection, Manassas, VA, USA) were grown in Dulbecco’s Modified Eagle Medium (DMEM) 5% serum supplemented with gentamicin 0.05 mg/mL in a moisture-saturated atmosphere containing CO
2 5% at 37 °C (VWR Basic CO
2 Incubator Air jacket 5.3CF). After a first pass, cells were divided (2000 cells/wells) on a 24 well microplates. Cells were pretreated with Thykamine extracts for 24 h in the same culture environment and conditions as growth conditions. After pretreatment, cells were washed twice with DMEM 5% serum and then activated with liposaccharide (LPS, Sigma #L4391-1MG) 100 ng/mL to produce NO for a period of 24 h. NO production was measured in the supernatants using the Griess reagent method and kit (Promega #G2930) and NO concentration were calculated with a standard curve of NO
2− [
11].
2.5. Protection of Lipid Peroxidation by Thykamine
Thykamine were tested for its capacity to protect against lipid peroxidation. The assay used was an adaptation of a spectrometric cuvette method [
12] to a microplate (Greiner Bio-One UV-Star) test. Lipidic emulsions were prepared with linoleic acid, reactive oxygen species were generated by 2,2-azobis (2-methylpropionamidine) dihydrochloride (AAPH) at 37 °C in continuous stirring at 234 nm for 45 min (Microplate Reader xMark, Biorad). Areas under the curves at 234 nm were calculated and the differences between Thykamine samples.
2.6. Detection of SOD in Thykamine
SOD zymography was performed in non-denaturating conditions on a polyacrylamide gel (acrylami-de/bis-acrylamide 30%/10%) [
13,
14]. Thykamine extracts were added to obtain a final concentration of 0.2 mg proteins. Electrophoresis was run at 80 V for 3 h. Activity was revealed with nitroblue tetrazolium solution (0.1%). Superoxide ions were generated by adding riboflavin. The presence of SOD-like enzyme was revealed by white spots on the blue colored gel.
For the determination of SOD activity, SOD was isolated from Thykamine extract using 0.1 mg/mL in 50 mM potassium phosphate buffer (pH 7.8), 1 mM EDTA, and 2% (
w/
v) polyvinylpolypyrrolidone. After centrifugation at 10,000×
g for 30 min at 4 °C, the supernatant was collected and SOD activity was assayed using the photo-oxidation of riboflavin generating ROS, including O
2−. These anions reduced nitro blue tetrazolium (NBT) at 560 nm [
7].
2.7. Evaluation of O2− Production
The production of O2− was determined using the reduction of cytochrome c. Neutrophils (107 cells/mL) were preincubated 30 min at 37 °C with diluent or Thykamine and then diluted to 106 cells/mL in the presence of 130 µM cytochrome c, followed by a stimulation with 0.1 µM fMLP for 5 min at 37 °C. Absorbance of supernatants was measured at 540 and 550 nm. Amounts of O2− present in supernatants were determined by the difference between the two readings and using an extinction coefficient of 21.1.
2.8. Flow Cytometric Analysis of Neutrophils Pretreated with Thykamine
Flow cytometric analyses were performed on neutrophils resuspended in HBSS. Due to the structure of Thykamine, neutrophils pretreated with these thylakoid membranes were directly visible through the spontaneous fluorescence of the compound monitored at an excitation wavelength of 488 nm and an emission wavelength of 675 nm. Samples were analyzed with a flowcytometer EPICS-XL (Beckman Coulter, Miami, FL, USA). FSC was set on linear scales, SSC and fluorescence channels (FL4) were set on logarithmic scales.
2.9. Confocal Microscopy of Neutrophils under Thykamine
Neutrophils were preincubated with different concentrations of Thykamine for 30 min before evaluation by confocal microscopy. Neutrophils were resuspended in 50 mL phosphate-buffered saline (PBS), placed on coverslips, air-dryed, and fixed in 4% paraformaldehyde (pH 7.4) for 15 min. They were further washed with PBS before analysis. Images were collected on an Olympus BX-61 confocal laser microscope and analyzed with the application program Fluoview 500.
2.10. Monitoring of Neutrophil Chemotaxis by Fluorescence
Chemotaxis of neutrophils was studied by the following two fluorescence-based neutrophil migration methods: Transpolycarbonate migration (TPM) assay and transendothelial migration (TEM) assay [
15,
16]. TPM assay was performed using a disposable 96-well chemotaxis chamber with an 8 µm pore size polycarbonate filter. Freshly isolated neutrophils (1 × 10
7 cells/mL) were incubated with 5 µg/calcein-AM for 30 min at 37 °C in the dark. Total fluorescence was obtained from a known number of neutrophils. The test was performed by placing neutrophils on the filter followed by a migration for 1 h (37 °C and 5% CO
2) in the dark. Non-migrating cells were removed by gently wiping the filter with a tissue. Cell migration was measured with a microplate fluorescence reader (FL600; Bio-Tek Instruments, Winooski, VT, USA) with bottom-read configuration (excitation 485 nm; emission 530 nm) [
15].
For TEM assay, human endothelial cells (HUVEC) were seeded out on PVP-free collagen-coated polycarbonate filters (pores of 8 µm) in Transwell culture plate inserts at 1.5 × 105 cells/0.4 mL/culture plate insert. After the obtainment of a complete monolayer of endothelial cells, neutrophils, preincubated with calcein-AM as above, were added to the upper chamber (4 × 105 cells), while the lower chamber was filled with or without 10−7 M LTB4, IL-8 or fMLP. Transmigration of neutrophils through the endothelial monolayer was allowed to proceed during half an hour. Non-migrating neutrophils were removed and migration of neutrophils was evaluated by the fluorescence measured in the lower chamber with a microplate fluorescence reader (excitation 485 nm; emission 530 nm). In both assays, results were expressed in percentage of cells that migrated toward the chemotactic agent.
2.11. Flow Cytometric Analysis of Phagocytosis
Phagocytosis by neutrophils was studied by monitoring the process of internalization of fluorescent labeled bacterial particles as indicated in the Vybrant Phagocytosis assay kit V-6694 (Molecular Probes Inc., Eugene, OR, USA) with slight modifications. Samples were analyzed with a flowcytometer EPICS-XL as above, and the fluorescence intensity was determined at 485 nm excitation and 530 nm emission wavelengths. Results were expressed as arbitrary units of fluorescence of neutrophils that have phagocytized fluorescent particles among 10,000 cells analyzed.
2.12. Analysis of Neutrophil Degranulation
Flow cytometry and ELISA measurements were used to analyze degranulation by neutrophils. Exocytosis of granules was assessing by evaluating the appearance of the granular markers CD63 (primary granules) and CD66b (secondary granules) at the cell surface, using anti-CD63or CD67 antibodies and fluorescent secondary antibody as previously reported [
17]. Results were expressed in arbitrary units of fluorescence intensity.
Exocytosis of neutrophils was also studied by assessing the release of MPO and LF in supernatants. Neutrophils (10 million/mL) were stimulated 5 min at 37 °C with vehicle or 10
−7 M fMLP in the presence or absence of 1 µg/mL cytochalasin B, an actin depolymerizing agent known to amplify granule exocytosis. One hundred (100) µL of supernatant or cells were added to 2.4 mL of a mixture of potassium phosphate buffer, 0.2 mg/mL of O-dianiside dihydrochloride and 500 µL of 0.003% H
2O
2. MPO contents were obtained by measuring absorbance at 460 nm and calculated from human MPO calibration. Results were expressed in percentages (ratio extra-cellular/intra- + extra-cellular materials). Release of LF was measured by ELISA [
18]. Absorbance was read at 450 nm with a microplate reader, and the LF concentration was calculated by comparison to the human LF calibration curve. Results are expressed in ng/mL.
2.13. Analysis of LDH Release
LDH release assay was used to determine the effects of Thykamine on neutrophils viability. Neutrophils and Thykamine were incubated 1 h at 37 °C before centrifugation. Both supernatants and pelleted neutrophils were collected separately for testing. Triton X-100 was used to lyse neutrophils cells. Reaction solution was composed of 1.25 mL substrate (0.14 mg/mL NADH in 0.1 M sodium phosphate buffer, pH 7.35) and 50 μL of pyruvate solution (Sigma, St-Louis, MO, USA) and was added to 50 µL of lysed cell or supernatant samples. Colorimetric readings were measured at 340 nm. Results were expressed as percentages of the ratio of absorbance read in supernatants by the total absorbance (cells plus supernatant).
2.14. Statistics
Results are presented as means and ± SEM. Statistical analyses were performed using GraphPad Instat 3.0 (GraphPad Software, Inc., San Diego, CA, USA). Sample groups were analyzed by using paired or unpaired Student’s t test. Significance was set at two-tailed p value < 0.05.
4. Discussion
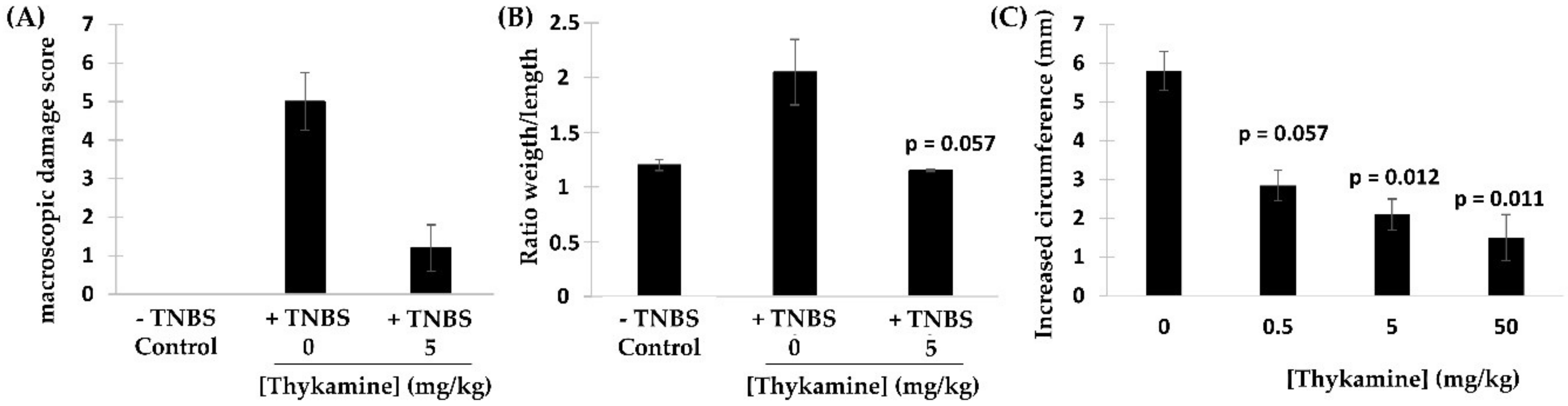
Results presented in this article demonstrated the high properties of Thykamine extracts to regulate and inhibit inflammation through their modulating activity on physical and biochemical parameters of inflammation. Thykamine injected intraperitoneally to rats reduced the inflammatory process of TNBS-induced colitis and carrageenan-induced paw edema (
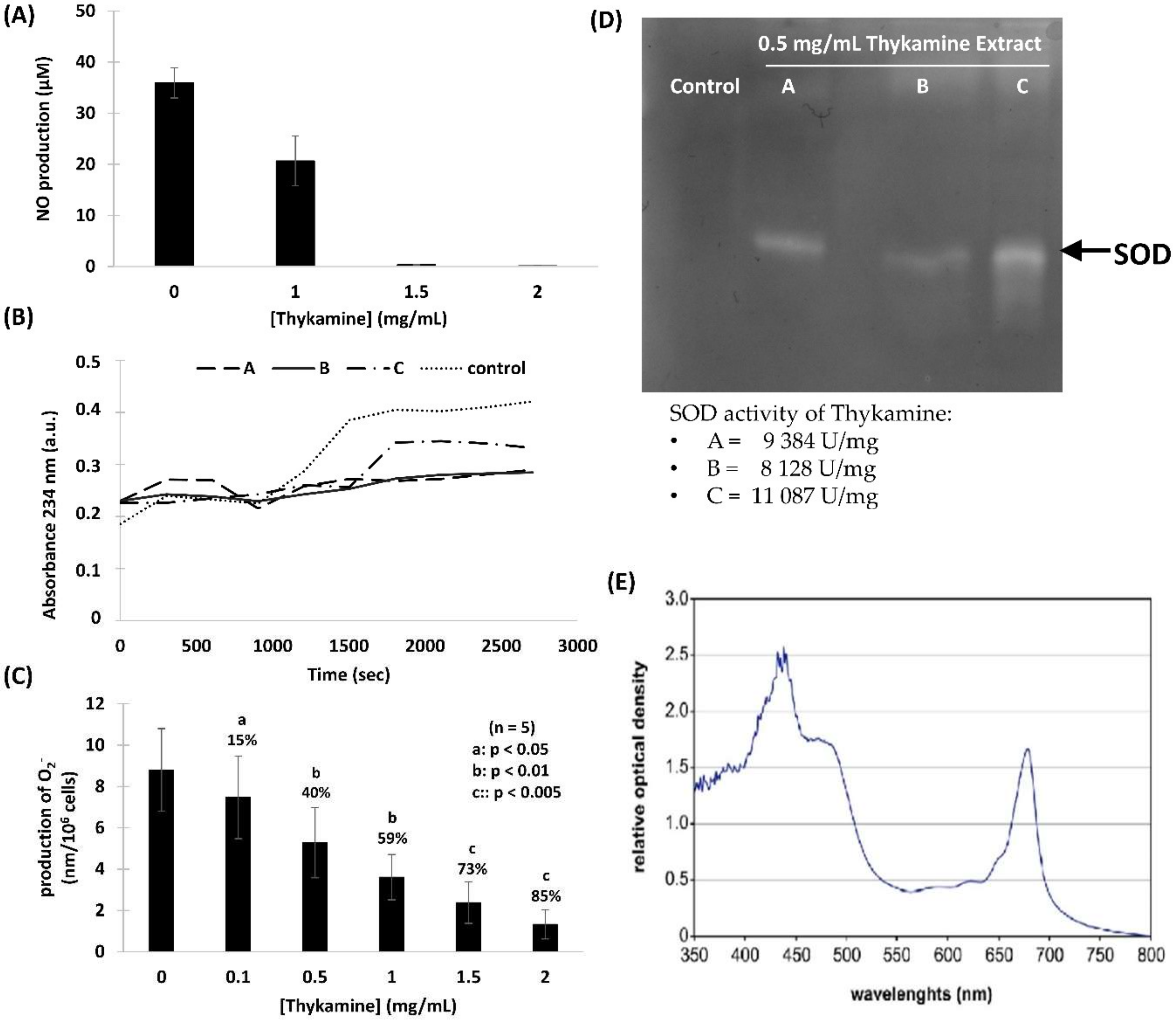
Figure 1). Thykamine extracts also protected against lipid peroxidation and inhibit NO production in J774A.1 cells (
Figure 2). The presence of SOD enzyme having a relevant superoxide inhibition activity as well as carotenoids in Thykamine extracts could account for the prevention against lipid peroxidation (
Figure 2). Moreover, SOD inhibits the formation of ONOO
− by decreasing superoxide levels that prevent the peroxidant effects produced by ONOO
−. Asada (2006) has demonstrated that thylakoid membranes, forming Thykamine extracts, contained Cu/Zn-SOD enzymes [
26]. Glycolipids composition of thylakoid membranes of Thykamine could also be involved in the inhibition of NO production since a treatment with these lipids blocked LPS-induced Inducible nitrite oxide synthase (iNOS) expression in HUVECs [
27]. The bilayer lipid membranes of Thykamine extracts are constituted of glycolipids including monogalactosyl diacylglycerol (MGDG), digalactosyl diacylglycerol (DGDG) and sulfoquinovosyl diacylglycerol (SQDG) [
27].
The involvement and role of redox molecules, such as NO and ROS, as key mediators of immunity have recently gained improved attention and appreciation. Physiological mechanisms conducting to an imbalance in these species are known to be implicated in diseases such as diabetes, cancer, heart and lung disease, autoimmune diseases, ageing and various infectious diseases. Beside SOD and other antioxidants found on Thykamine membranes, a possible mechanism of action of this anti-inflammatory botanical agent could be the binding between [2Fe-2S] clusters, located on Thykamine, and the cysteine of the enzyme glutaredoxin (Grx1). It is now reported that Grx1 accumulated in various inflammatory disease [
28] having a pro-inflammatory activity [
29]. Thus, development of Grx1 inhibitors could be efficient agents for anti-inflammatory activity. The binding of Grx1 on Thykamine could adduct the nucleophilic active site of Grx1 and explain its anti-inflammatory activity.
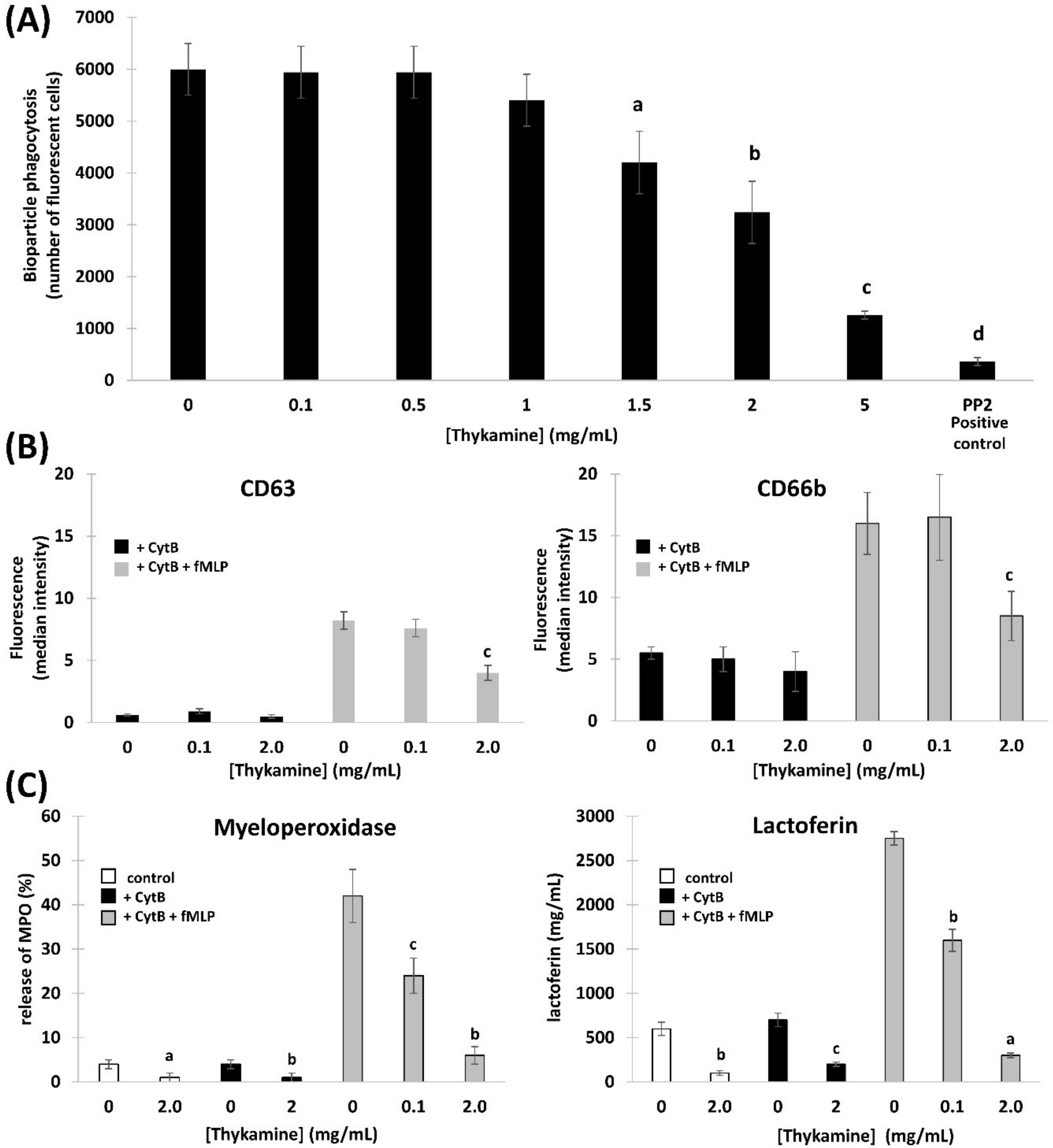
Results presented in this study show that Thykamine extract isolated from spinach have antioxidant properties that could explained their ability to modulate phlogogenic functions of human blood neutrophils. Moreover, the resolution of inflammation in two rat models could be explained by the effects of Thykamine extracts on human neutrophils.
Primary functions of neutrophils, like ROS production and degranulation following chemotaxis toward and phagocytosis of foreign particles abnormally present in tissues, are associated with an obligatory inflammatory response. However, this inflammatory reaction can pathologically persist with deleterious impact on tissues and subject health. Normal human blood neutrophils were shown to be modulated dose-dependently when preincubated with Thykamine that decreased the production of O
2−, and degranulation of primary and secondary granules stimulated by fMLP, as well as chemotaxis in response to fMLP, IL-8 or LTB4, and phagocytosis of bioparticles. Note that 2 mg/mL Thykamine inhibited 85% of fMLP-induced production of O
2− (
Figure 2C). As the Thykamine extracts were found to contain relevant SOD activities and carotenoid concentration, Thykamine could interfere with neutrophil functions through these natural antioxidants. SOD activities scavenging increase glutathione enzymes as well as carotenoids from spinach and have an antioxidant activity on human erythrocytes by increasing gluthatione reductase and decreasing catalase, an effect mainly related to lutein [
30]. Interestingly, β-carotene has been reported to abrogate the generation of ROS induced by
para-nonylphenol in human blood neutrophils and in whole blood in vitro [
31]. These data suggest that Thykamine extracts, at least in part through SOD and carotenoids, could impact on neutrophil machinery by modulating signals associated with the regulation of NADPH-oxidase. It is also useful to stress that 0.1 to 2 mg/mL Thykamine linearly inhibited (from 15 to 85%) neutrophil production of O
2− (
Figure 2C). These data suggest that carotenoids in the thylakoid complex Thykamine did not show any pro-oxidant actions as reported with high concentrations of pure carotenoids [
4,
32]. Similarly, carotenoids can have, depending on concentrations, anti- or pro-apoptotic effects on tumor cells [
33]. Beta-carotene cleavage products have been recently reported to have effects on spontaneous apoptosis of human neutrophils [
34]. Results on the absence of effects of Thykamine on neutrophil viability suggest that carotenoids contained in Thykamine do not impact neutrophil apoptosis. Moreover, our results indicate that β-carotene in Thykamine was not associated with pro-apoptotic cleavage products, or if so Thykamine was capable of their neutralization. This is particularly important since β-carotene elevated concentrations and cleavage products have been related to neutrophil toxic effects, especially in smokers [
35,
36].
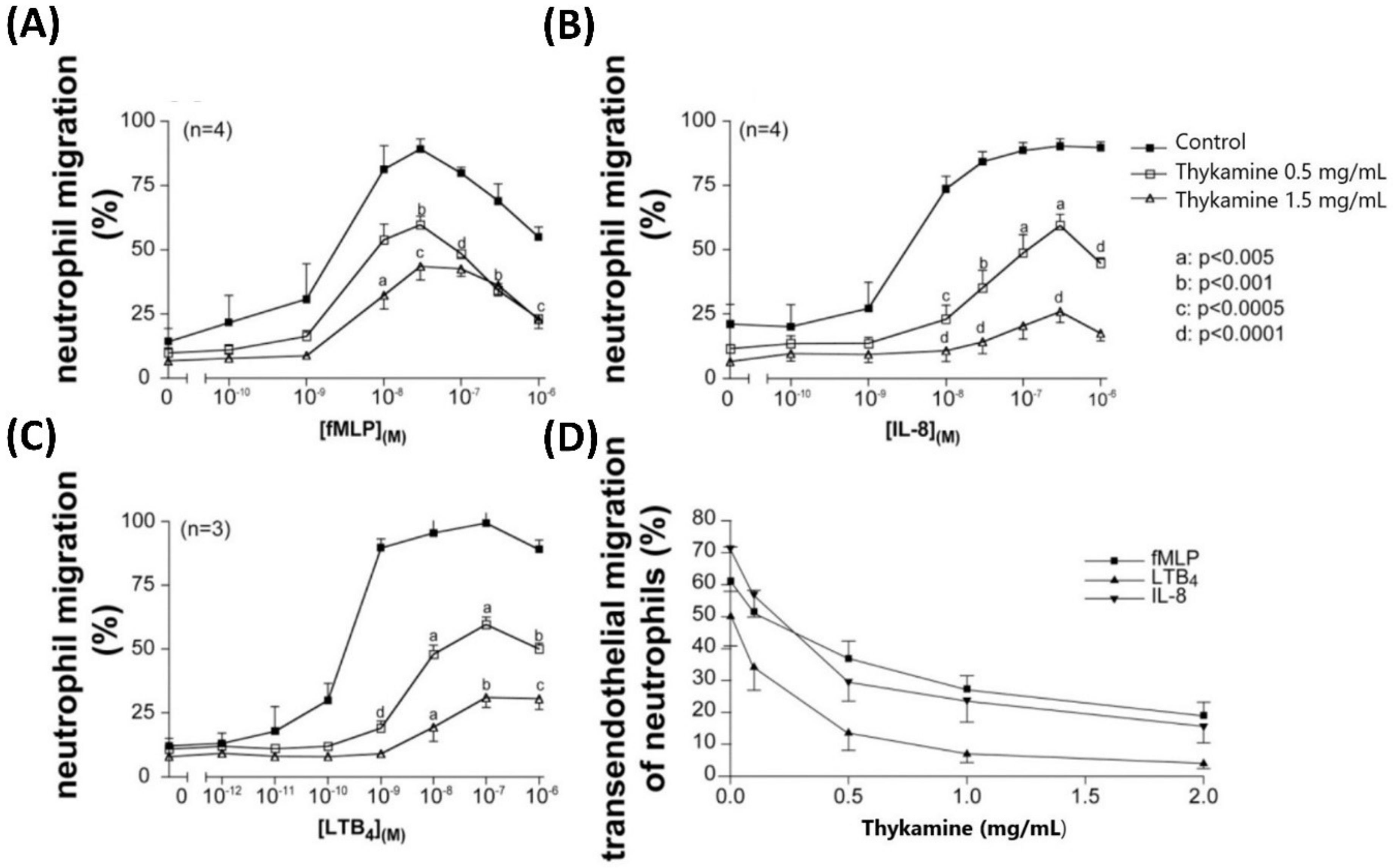
Considering the other primary functions of human blood neutrophils that are involved in inflammation, we showed significant inhibition by Thykamine extracts of several processes including; (1) chemotaxis of human blood neutrophil induced by three different agonists (
Figure 3), (2) phagocytosis of bioparticles by neutrophils, and (3) degranulation of neutrophil via the release of primary and secondary granules induced by fMLP (
Figure 4). Interestingly, β-carotene intake alone was shown to have no effect or increasing effect on neutrophil chemotaxis in cows suggesting that this anti-oxidant molecule alone was not related to the inhibitory effects of Thykamine extracts on human neutrophil chemotaxis [
37,
38]. It is also noteworthy to stress that Thykamine extracts were able to down-regulate the spontaneous degranulation of human blood neutrophils in vitro, an effect that could be associated with a reduction of their spontaneous apoptosis. Finally, the inhibitory effect of Thykamine on neutrophil phagocytosis, that was only partial (46% inhibition at 2 mg/mL Thykamine (
Figure 4), could be a beneficial anti-inflammatory effect while this major neutrophil function of phagocytosis was maintained to a certain extent. This reduction of neutrophil phagocytosis by Thykamine could be explained by SOD enzyme since it has been shown that exogenously added SOD induced neutrophil apoptosis by the dismutation of superoxide radicals [
39]. It is noteworthy that, for the first time, a natural product has been shown to reduce phagocytosis by normal human neutrophils.
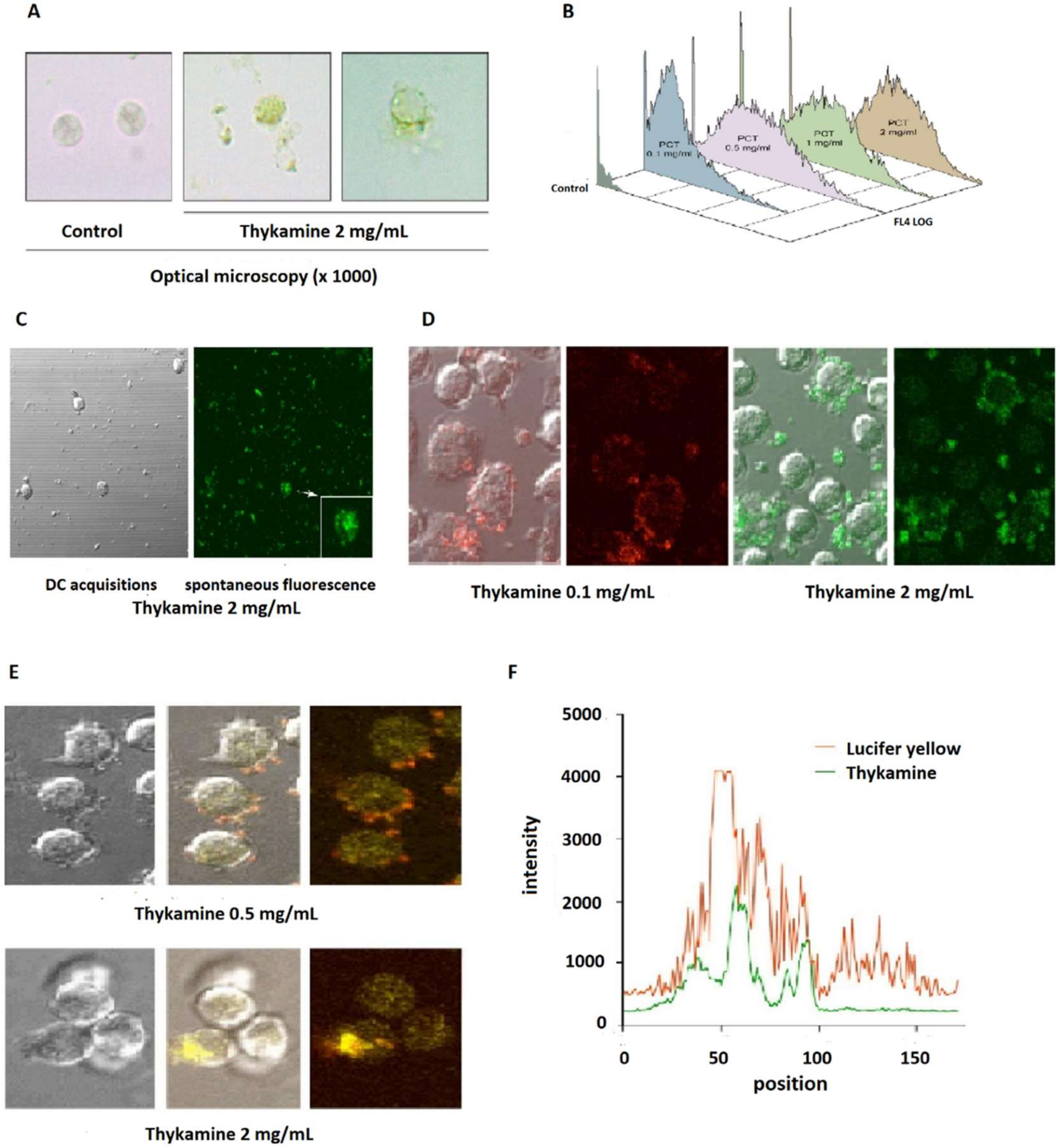
Different techniques of microscopy allowed us to localize easily Thykamine particles, that are spontaneously fluorescent, in the extra-cellular milieu, as well as Thykamine particles wrapping and inside the cells with dose-dependent modifications of granularity (
Figure 5A). Short-term incubations (<1 h) that showed various inhibitory effects of Thykamine on neutrophil functions indicate a rapid cellular penetration of this complex. A major function of defense supported by neutrophils is rapid phagocytosis of foreign particles [
40]. This could be the case for Thykamine particles. However, the data presented in
Figure 5E,F using Lucifer Yellow dye indicate that Thykamine particles also entered the cell by pinocytosis. This mode of cell entry could be supported by a report on the effects of carotenoids on gap junctions. Carotenoids were showed to significantly upregulate expression of connexin 43 protein and gap junctional intercellular communication, as demonstrated by Lucifer Yellow dye transfer [
41]. Thus, it is possible that Thykamine, through their carotenoids, is capable of a rapid entry in neutrophils by pinocytosis. This rapid penetration of Thykamine in neutrophils could be also associated with effects on the complex neutrophil enzyme machinery involved in the inflammatory process.
Further investigations are needed to decipher the exact mechanisms of anti-inflammatory action of this complex natural product that may require the integrity of its lipido-protidic composition to remain active. It is now recognized that phosphoinositide 3-kinases (PI3Ks) are important signaling lipid kinases that regulate multiple biological functions [
42]. For example, increased PI3K activity has been observed in tumor cells [
43]. Different isoform of PI3K plays specific roles in different cell types. The PI3K isoform p110∂, mainly expressed in white blood cells, plays a key role in the immune system. For example, p110δ PI3K plays a main role in promoting neointimal thickening and inflammatory processes during vascular stenosis, with its inhibition providing significant reduction in restenosis following carotid injury [
44]. The p110δ also is involved in B-cell antigen and IL-4 receptor signaling [
45]. Since PI3K isoform p110∂ has a key role in immune system, it would be interested to verify how it is influenced or inhibited by Thykamine.
In conclusion, the present report adds a new dimension to the anti-inflammatory effect of the natural product Thykamine from spinach thylakoids previously suspected in human macrophages [
6]. Thykamine has in vivo anti-inflammatory effects in two animal models of acute inflammation directly related to the presence of neutrophils. As a corollary, Thykamine extracts significantly impact human neutrophil primary functions by reducing their possible toxic effects during the inflammatory process.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




