Sex and Gender Influences on Cancer Immunotherapy Response

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Cancer Susceptibility and Development
2.1. Sex–Gender Differences in Cancer
2.2. Viruses and Cancer
3. Sex, Gender and the Immune System
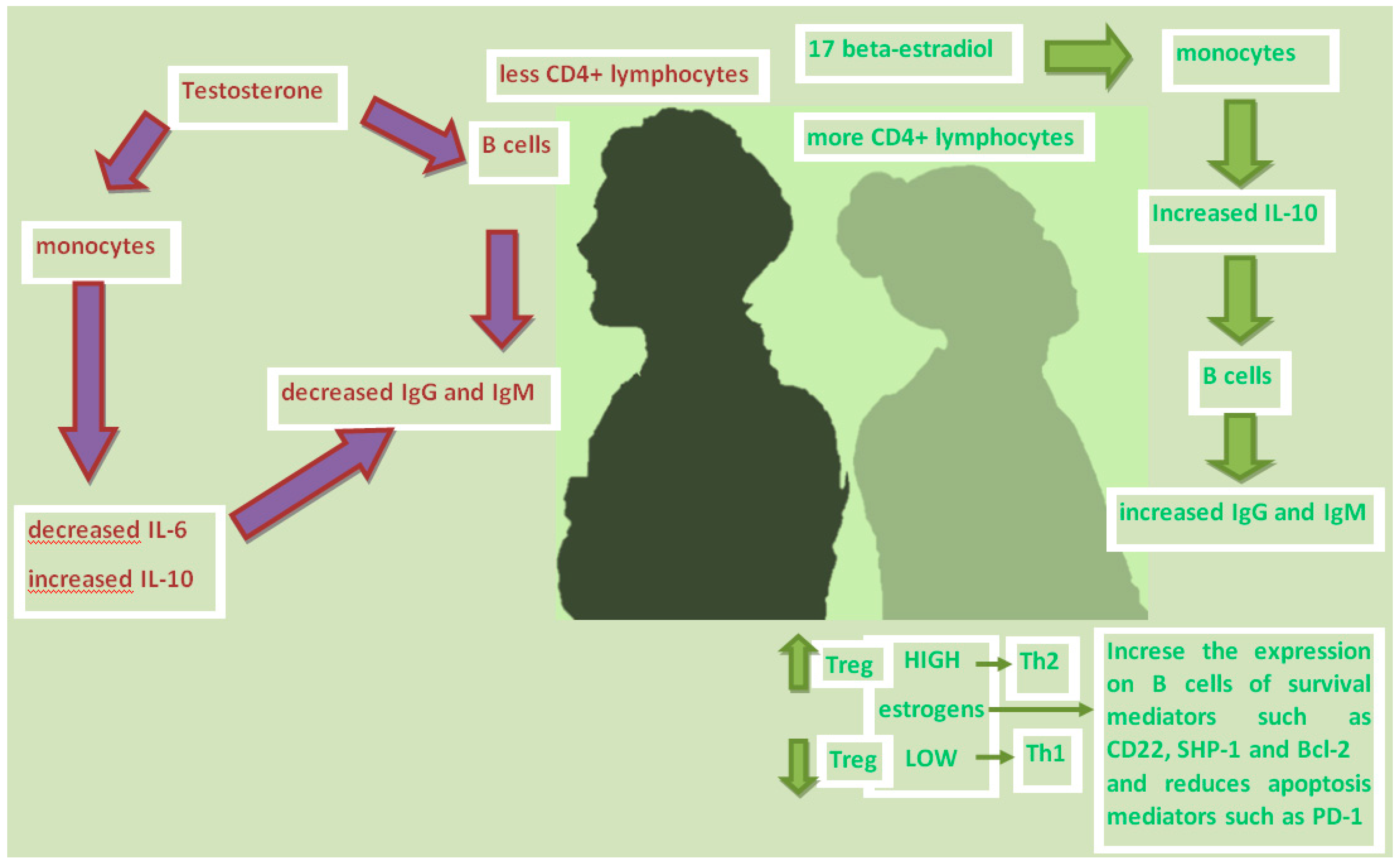
3.1. Sex Hormones and Immunity
3.2. Immunity, Microbiome and Aging
3.3. Sex, Genes, Immunity and Cancer
3.4. Other Factors Influencing the Immune System and Response to Immunotherapy
4. mTOR as a Link between the Immune System and Sex Hormones
5. Sex and Effectiveness of Cancer Immunotherapy
5.1. Immune Checkpoint Inhibitors

{kind=link}
| Meta-Analysis | Sex | OS-HR (anti-CTLA4) | OS-HR (anti-PD1) | PFS-HR (anti-PD1) | OS-HR (anti-CTLA4 + anti-PD1) | ||||
|---|---|---|---|---|---|---|---|---|---|
| p | p | p | p | ||||||
| Botticelli et al. [70] | M | 0.65 | 0.078 | 0.72 | 0.285 | 0.66 | 0.158 | ||
| F | 0.79 | 0.81 | 0.85 | ||||||
| Pinto et al. [79] | M | 0.76 | 0.69 | ||||||
| F | 1.03 | ||||||||
| Conforti et al. [28] | M | 0.72 | 0.0019 | ||||||
| F | 0.86 | ||||||||
| Wallis et al. [81] | M | 0.75 | 0.60 | ||||||
| F | 0.77 | ||||||||
5.2. Vaccines and Other Therapies
6. The Role of Gender in Cancer Immunotherapy
7. Patients’ Sex and Adverse Events of Cancer Immunotherapy
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- De Martinis, M.; Sirufo, M.M.; Suppa, M.; Di Silvestre, D.; Ginaldi, L. Sex and gender aspects for patient stratification in allergy prevention and treatment. Int. J. Molsci. 2020, 24, 1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, A.D.; Oertelt-Prigione, S.; Adjei, A.; Buclin, T.; Cristina, V.; Csajka, C.; Coukos, G.; Dafni, U.; Dotto, G.P.; Ducreux, M.; et al. Gender medicine and oncology: Report and consensus of an ESMO workshop. Ann. Oncol. 2019, 1, 1914–1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Springer, K.W.; Hankivsky, O.; Bates, L.M. Gender and health: Relational, intersectional, and biosocial approaches. Soc. Sci. Med. 2012, 74, 1661–1666. [Google Scholar] [CrossRef] [PubMed]
- Marino, M.; Masella, R.; Bulzomi, P.; Campesi, I.; Malorni, W.; Franconi, F. Nutrition and human health froma sex-gender perspective. Mol. Asp. Med. 2011, 32, 1–70. [Google Scholar] [CrossRef] [PubMed]
- Franconi, F.; Campesi, I.; Colombo, D.; Antonini, P. Sex-Gender Variable: Methodological Recommendations for Increasing Scientific Value of Clinical Studies. Cells 2019, 8, 476. [Google Scholar] [CrossRef] [Green Version]
- Ribas, A.; Wolchok, J.D. Cancer immunotherapy using checkpoint blockade. Science 2018, 359, 1350–1355. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.F.; Song, H.F.; Zhang, Q.; Yan, C.X. Pan-cancer immunogenomic analyses reveal sex disparity in the efficacy of cancer immunotherapy. Eur. J. Cancer 2020, 126, 136–138. [Google Scholar] [CrossRef]
- Prakash, V.S.; Mansukhani, N.A.; Helenowski, I.B.; Woodruff, T.K.; Kibbe, M.R. Sex Bias in Interventional Clinical Trials. J. Womens Health 2018, 27, 1342–1348. [Google Scholar] [CrossRef]
- Gemmati, D.; Varani, K.; Bramanti, B.; Piva, R.; Bonaccorsi, G.; Trentini, A.; Manfrinato, M.C.; Tisato, V.; Carè, A.; Bellini, T. “Bridging the Gap” Everything that Could Have Been Avoided If We Had Applied Gender Medicine, Pharmacogenetics and Personalized Medicine in the Gender-Omics and Sex-Omics Era. Int. J. Mol. Sci. 2019, 21, 296. [Google Scholar] [CrossRef] [Green Version]
- Legato, M.J. The “biological sex or gender?” Debate: “Everything flows, nothing stands still. Nothing endures but change”. Gend. Med. 2011, 8, 161–163. [Google Scholar] [CrossRef]
- Klinge, I. Gender perspectives in European research. Pharmacol. Res. 2008, 58, 183–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Straface, E.; Gambardella, L.; Brandani, M.; Malorni, W. Sex differences at cellular level: “cells have a sex”. Handb. Exp. Pharmacol. 2012, 214, 49–65. [Google Scholar]
- Clocchiatti, A.; Cora, E.; Zhang, Y.; Dotto, G.P. Sexual dimorphism in cancer. Nat. Rev. Cancer 2016, 16, 330–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massimini, M.; Palmieri, C.; De Maria, R.; Romanucci, M.; Malatesta, D.; De Martinis, M.; Maniscalco, L.; Ciccarelli, A.; Ginaldi, L.; Buracco, P.; et al. 17-AAG and Apoptosis, Autophagy, and Mitophagy in Canine Osteosarcoma Cell Lines. Vet. Pathol. 2017, 54, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Lista, P.; Straface, E.; Brunelleschi, S.; Franconi, F.; Malorni, W. On the Role of Autophagy in Human Diseases: A Gender Perspective. J. Cell Mol. Med. 2011, 15, 1443–1457. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Shao, X.; Wang, X.; Liu, L.; Liang, H. Sex disparities in cancer. Cancer Lett. 2019, 466, 35–38. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Lee, S.; Lee, E.; Lim, H.; Shin, J.Y.; Jung, J.; Kim, S.G.; Moon, A. Sex-biased differences in the correlation between epithelial-to-mesenchymal transition-associated genes in cancer cell lines. Oncol. Lett. 2019, 18, 6852–6868. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.B.; Dawsey, S.M.; Freedman, N.D.; Inskip, P.D.; Wichner, S.M.; Quraishi, S.M.; Devesa, S.S.; McGlynn, K.A. Sex disparities in cancer incidence by period and age. Cancer Epidemiol. Biomark. Prev. 2009, 18, 1174–1182. [Google Scholar] [CrossRef] [Green Version]
- Cook, M.B.; McGlynn, K.A.; Devesa, S.S.; Freedman, N.D.; Anderson, W.F. Sex Disparities in Cancer Mortality and Survival. Cancer Epidemiol. Biomark. Prev. 2011, 20, 1629–1637. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; He, Z.; Wang, X.; Li, H.; Wu, T.; Sun, X.; Wu, K.; Liu, X.-S. Can tumor mutational burden determine the most effective treatment for lung cancer patients? Lung Cancer Manag. 2020, 8, LMT21, Published 14 January 2020. [Google Scholar] [CrossRef]
- Ortona, E.; Pierdominici, M.; Rider, V. Editorial: Sex Hormones and Gender Differences in Immune Responses. Front. Immunol. 2019, 10, 1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegrini, P.; Contasta, I.; DelBeato, T.; Ciccone, F.; Berghella, A.M. Gender-specific cytokine pathways, targets, and biomarkers for the switch from health to adenoma and colorectal cancer. Clin. Dev. Immunol. 2011, 2011, 819724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Konishi, N.; Miki, C.; Yoshida, T.; Tanaka, K.; Toiyama, Y.; Kusunoki, M. Interleukin-1 receptor antagonist inhibits the expression of vascular endothelial growth factor in colorectal carcinoma. Oncology 2005, 68, 138–145. [Google Scholar] [CrossRef]
- Wetzler, M.; Kurzrock, R.; Estrov, Z.; Kantarjian, H.; Gisslinger, H.; Underbrink, M.P.; Talpaz, M. Altered levels of interleukin-1 beta and interleukin-1 receptor antagonist in chronic myelogenous leukemia: Clinical and prognostic correlates. Blood 1994, 84, 3142–3147. [Google Scholar] [CrossRef] [Green Version]
- Chraa, D.; Naim, A.; Olive, D.; Badou, A. T lymphocyte subsets in cancer immunity: Friends or foes. J. Leukoc. Boil. 2018, 105, 243–255. [Google Scholar] [CrossRef]
- Furman, D.; Hejblum, B.P.; Simon, N.; Jojic, V.; Dekker, C.L.; Thiébaut, R.; Tibshirani, R.J.; Davis, M.M. Systems Analysis of Sex Differences Reveals an Immunosuppressive Role for Testosterone in the Response to Influenza Vaccination. Proc. Natl. Acad. Sci. USA 2014, 111, 869–874. [Google Scholar] [CrossRef] [Green Version]
- Conforti, F.; Pala, L.; Bagnardi, V.; De Pas, T.; Martinetti, M.; Viale, G.; Gelber, R.D.; Goldhirsch, A. Cancer immunotherapy efficacy and patients’ sex: A systematic review and meta-analysis. Lancet Oncol. 2018, 19, 737–746. [Google Scholar] [CrossRef]
- Ruggieri, A.; Malorni, W.; Ricciardi, W. Gender disparity in response to anti-viral vaccines: New clues toward personalized vaccinology. Ital. J. Gend.-Specif. Med. 2016, 2, 93–98. [Google Scholar]
- Garutti, P.; Montori, S.; Bazzan, E.; Tarabbia, C. Gender differences in the epidemiology and prevention of human papillomavirus (HPV) and HPV-related diseases. Ital. J. Gend.-Specif. Med. 2018, 4, 152–156. [Google Scholar]
- Carrel, L.; Willard, H.F. X-inactivation profile reveal sex tensive variability in X-linked gene expression in females. Nature 2005, 434, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, I.; Dejager, L.; Libert, C. X-chromosome-located microRNAs in immunity: Might they explain male/female differences? The X chromosome-genomic context may affect X-located miRNAs and downstream signaling, thereby contributing to the enhanced immune response of females. Bioessays 2011, 33, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Kanda, N.; Tamaki, K. Estrogen enhances immunoglobulin production by human PBMCs. J. Allergy Clin. Immunol. 1999, 103, 282–288. [Google Scholar] [CrossRef]
- Mirandola, L.; Wade, R.; Verma, R.; Pena, C.; Hosiriluck, N.; Figueroa, J.A.; Cobos, E.; Jenkins, M.R.; Chiriva-Internati, M. Sex-driven differences in immunological responses: Challenges and opportunities for the immunotherapies of the third millennium. Int. Rev. Immunol. 2015, 34, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Verthelyi, D. Sex hormones as immunomodulators in health and disease. Int. Immunopharmacol. 2001, 1, 983–993. [Google Scholar] [CrossRef]
- Canellada, A.; Alvarez, I.; Berod, L.; Gentile, T. Estrogen and Progesterone Regulate the IL-6 Signal Transduction Pathway in Antibody Secreting Cells. J. Steroid Biochem. Mol. Biol. 2008, 111, 255–261. [Google Scholar] [CrossRef]
- Salem, M.L. Estrogen, a Double-Edged Sword: Modulation of TH1-And TH2-mediated Inflammations by Differential Regulation of TH1/TH2 Cytokine Production. Curr. Drug Targets Inflamm. Allergy. 2004, 3, 97–104. [Google Scholar] [CrossRef]
- Thompson, M.G.; Peiffer, D.S.; Larson, M.; Navarro, F.; Watkins, S.K. FOXO3, estrogen receptor alpha, and androgen receptor impact tumor growth rate and infiltration of dendritic cell subsets differentially between male and female mice. Cancer Immunol. Immunother. 2017, 66, 615–625. [Google Scholar] [CrossRef]
- Kanda, N.; Tsuchida, T.; Tamaki, K. Testosterone inhibits immunoglobulin production by human peripheral blood mononuclear cells. Clin. Exp. Immunol. 1996, 106, 410–415. [Google Scholar] [CrossRef]
- Mohamad, N.; Wong, S.; Hasan, W.N.W.; Jolly, J.J.; Nur-Farhana, M.F.; Ima-Nirwana, S.; Chin, K.Y. The relationship between circulating testosterone and inflammatory cytokines in men. Aging. Male 2019, 22, 129–140. [Google Scholar] [CrossRef]
- Sthoeger, Z.M.; Chiorazzi, N.; Lahita, R.G. Regulation of the immune response by sex hormones. I. Invitro effects of estradiol and testosterone on pokeweed mitogen-induced human B cell differentiation. J. Immunol. 1988, 141, 91–98. [Google Scholar] [PubMed]
- Fish, E.N. The X-files in immunity: Sex-based differences predispose immune responses. Nat. Rev. Immunol. 2008, 8, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Kyurkchiev, D.; Ivanova-Todorova, E.; Hayrabedyan, S.; Altankova, I.; Kyurkchiev, S. Female Sex Steroid Hormones Modify Some Regulatory Properties of Monocyte-Derived Dendritic Cells. Am. J. Reprod. Immunol. 2007, 58, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Miyagi, M.; Aoyama, H.; Morishita, M.; Iwamoto, Y. Effects of Sex Hormones on Chemotaxis of Human Peripheral Polymorphonuclear Leukocytes and Monocytes. J. Periodontol. 1992, 63, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Seillet, C.; Rouquié, N.; Foulon, E.; Douin-Echinard, V.; Krust, A.; Chambon, P.; Arnal, J.F.; Guéry, J.C.; Laffont, S. Estradiol promotes functional responses in inflammatory and steady-state dendritic cells through differential requirement for activation function-1 of estrogen receptor α. J. Immunol. 2013, 190, 5459–5470. [Google Scholar] [CrossRef] [Green Version]
- Nowak, K.; Jablonska, E.; Ratajczak-Wrona, W. Neutrophils life under estrogenic and xenoestrogenic control. J. Steroid Biochem. Mol. Boil. 2019, 186, 203–211. [Google Scholar] [CrossRef]
- Klein, S.L.; Marriott, I.; Fish, E.N. Sex-based differences in immune function and responses to vaccination. Trans. R. Soc. Trop. Med. Hyg. 2015, 109, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Polanczyk, M.J.; Hopke, C.; Vandenbark, A.A.; Offner, H. Estrogen-mediated immunomodulation involves reduced activation of effector T cells, potentiation of Treg cells, and enhanced expression of the PD-1 costimulatory pathway. J. Neurosci. Res. 2006, 84, 370–378. [Google Scholar] [CrossRef]
- Pala, L.; Nezi, L.; de Pas, T.; Pennacchioli, E.; Cocorocchio, E.; Ferrucci, P.; Conforti, F.; Goldhirsch, A. Sex Differences in Efficacy and Toxicity of Systemic Cancer Treatments: Role of the Microbiome. J. Clin. Oncol. 2019, 37, 439. [Google Scholar] [CrossRef]
- Conforti, F.; Pala, L.; Goldhirsch, A. Different effectiveness of anticancer immunotherapy in men and women relies on sex-dimorphism of the immune system. Oncotarget 2018, 9, 31167–31168. [Google Scholar] [CrossRef]
- Conforti, F.; Pala, L.; Bagnardi, V.; Viale, G.; De Pas, T.; Pagan, E.; Pennacchioli, E.; Cocorocchio, E.; Ferrucci, P.F.; DeMarinis, F.; et al. Sex-based heterogeneity in response to lung cancer immunotherapy: A systematic review and meta-analysis. J. Natl. Cancer Inst. 2019, 111, 772–781. [Google Scholar] [CrossRef] [PubMed]
- Carè, A.; Bellenghi, M.; Matarrese, P.; Gabriele, L.; Salvioli, S.; Malorni, W. Sex disparity in cancer: Roles of microRNAs and related functional players. Cell Death Differ. 2018, 25, 477–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaunay, F.; Pettersson, K.; Tujague, M.; Gustafsson, J.A. Functional Differences between the Amino-Terminal Domains of Estrogen Receptors Alpha and Beta. Mol. Pharmacol. 2000, 58, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Frasor, J.; Danes, J.M.; Komm, B.; Chang, K.C.N.; Lyttle, C.R.; Katzenellenbogen, B.S. Profiling of Estrogen Up-And Down-Regulated Gene Expression in Human Breast Cancer Cells: Insights Into Gene Networks and Pathways Underlying Estrogenic Control of Proliferation and Cell Phenotype. Endocrinology 2003, 144, 4562–4574. [Google Scholar] [CrossRef]
- Trzeciak, A.R.; Barnes, J.; Ejiogu, N.; Foster, K.; Brant, L.J.; Zonderman, A.B.; Evans, M.K. Age, Sex, and Race Influence Single-Strand Break Repair Capacity in a Human Population. Free Radic. Biol. Med. 2008, 45, 1631–1641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ide, T.; Tsutsui, H.; Ohashi, N.; Hayashidani, S.; Suematsu, N.; Tsuchihashi, M.; Tamai, H.; Takeshita, A. Greater Oxidative Stress in Healthy Young Men Compared with Premenopausal Women. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 438–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matarrese, P.; Colasanti, T.; Ascione, B.; Margutti, P.; Franconi, F.; Alessandri, C.; Conti, F.; Riccieri, V.; Rosano, G.; Ortona, E.; et al. Gender Disparity inS usceptibility to Oxidative Stress and Apoptosis Induced by Autoantibodies Specific to RLIP76 in Vascular Cells. Antioxid. Redox Signal. 2011, 15, 2825–2836. [Google Scholar] [CrossRef]
- Hewagama, A.; Patel, D.; Yarlagadda, S.; Strickland, F.M.; Richardson, B.C. Stronger Inflammatory/Cytotoxic T-cell Response in Women Identified by Microarray Analysis. Genes Immun. 2009, 10, 509–516. [Google Scholar] [CrossRef]
- Rothenberger, N.J.; Somasundaram, A.; Stabile, L.P. The Role of the Estrogen Pathway in the Tumor Microenvironment. Int. J. Mol. Sci. 2018, 19, 611. [Google Scholar] [CrossRef] [Green Version]
- De Martinis, M.; Sirufo, M.M.; Suppa, M.; Ginaldi, L. New perspectives in food allergy. Int. J. Mol. Sci. 2020, 21, 1474. [Google Scholar] [CrossRef] [Green Version]
- Irelli, A.; Sirufo, M.M.; Scipioni, T.; DePietro, F.; Pancotti, A.; Ginaldi, L.; De Martinis, M. mTOR Links Tumor Immunity and Bone Metabolism: What are the Clinical Implications? Int. J. Mol. Sci. 2019, 21, 5841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martelli, A.M.; Evangelisti, C.; Chiarini, F.; McCubrey, J.A. The phosphatidyl inositol 3-kinase/Akt/mTOR signaling network as a therapeutic target in acute myelogenous leukemia patients. Oncotarget 2010, 1, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H. mTOR signaling in immune cells and its implications for cancer immunotherapy. Cancer Lett. 2017, 408, 182–189. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, J.S.; Massi, D.; Teng, M.W.L.; Mandala, M. PI3K-AKT-mTOR inhibition in cancer immunotherapy, redux. Semin. Cancer Biol. 2018, 48, 91–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, G.; Zippelius, A.; Wicki, A.; Mandalà, M.; Tang, F.; Massi, D.; Hemmings, B.A. Integrated Akt/PKB signaling in immunomodulation and its potential role in cancer immunotherapy. J. Natl. Cancer Inst. 2015, 107. [Google Scholar] [CrossRef] [Green Version]
- Conciatori, F.; Bazzichetto, C.; Falcone, I.; Pilotto, S.; Bria, E.; Cognetti, F.; Milella, M.; Ciuffreda, L. Role of mTOR Signaling in Tumor Microenvironment: An Overview. Int. J. Mol. Sci. 2018, 19, 2453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aaltonen, K.E.; Rosendahl, A.H.; Olsson, H.; Malmström, P.; Hartman, L.; Fernö, M. Association between insulin-like growth factor-1 receptor (IGF1R) negativity and poor prognosis in a cohort of women with primary breast cancer. BMC Cancer 2014, 14, 794. [Google Scholar] [CrossRef] [Green Version]
- Sobhani, N.; Generali, D.; Zanconati, F.; Bortul, M.; Scaggiante, B. Current status of PI3K-mTOR inhibition in hormone-receptor positive, HER2-negative breast cancer. World J. Clin. Oncol. 2018, 9, 172–179. [Google Scholar] [CrossRef]
- Alayev, A.; Salamon, R.S.; Berger, S.M.; Schwartz, N.S.; Cuesta, R.; Snyder, R.B.; Holz, M.K. mTORC1 directly phosphorylates and activates ERα upon estrogen stimulation. Oncogene 2016, 35, 3535–3543. [Google Scholar] [CrossRef]
- Botticelli, A.; Onesti, C.E.; Zizzari, I.; Cerbelli, B.; Sciattella, P.; Occhipinti, M.; Roberto, M.; DiPietro, F.; Bonifacino, A.; Ghidini, M.; et al. The sexist behaviour of immunecheckpoint inhibitors in cancer therapy? Oncotarget 2017, 8, 99336–99346. [Google Scholar] [CrossRef]
- Baar, E.L.; Carbajal, K.A.; Ong, I.M.; Lamming, D.W. Sex-and tissue-specific changes in mTOR signaling with age in C57BL/6Jmice. Aging Cell 2016, 15, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.Y.; Rodriguez, A.A.; Dastidar, S.G.; DelGreco, E.; Carr, K.L.; Sitzmann, J.M.; Academia, E.C.; Viray, C.M.; Martinez, L.L.; Kaplowitz, B.S.; et al. Increased 4E-BP1 Expression Protects against Diet-Induced Obesity and Insulin Resistance in Male Mice. Cell Rep. 2016, 16, 1903–1914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaturantabut, S.; Shwartz, A.; Evason, K.J.; Cox, A.G.; Labella, K.; Schepers, A.G.; Yang, S.; Acuña, M.; Houvras, Y.; Mancio-Silva, L.; et al. Estrogen Activation of G-Protein-Coupled Estrogen Receptor1 Regulates Phosphoinositide3-Kinase and mTOR Signaling to Promote Liver Growth in Zebrafish and Proliferation of Human Hepatocytes. Gastroenterology 2019, 156, 1788–1804. [Google Scholar] [CrossRef] [Green Version]
- Kono, M.; Fujii, T.; Lim, B.; Karuturi, M.S.; Tripathy, D.; Ueno, N.T. Androgen Receptor Function and Androgen Receptor-Targeted Therapies in Breast Cancer: A Review. JAMA Oncol. 2017, 1, 1266–1273. [Google Scholar] [CrossRef]
- Chen, H.; Zhou, L.; Wu, X.; Li, R.; Wen, J.; Sha, J.; Wen, X. The PI3K/AKT pathway in the pathogenesis of prostate cancer. Front. Biosci. (LandmarkEd) 2016, 21, 1084–1091. [Google Scholar]
- Audet-Walsh, É.; Dufour, C.R.; Yee, T.; Zouanat, F.Z.; Yan, M.; Kalloghlian, G.; Vernier, M.; Caron, M.; Bourque, G.; Scarlata, E.; et al. Nuclear mTOR acts as a transcriptional integrator of the androgen signaling pathway in prostate cancer. Genes Dev. 2017, 31, 1228–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jäger, U.; Fridrik, M.A.; Zeitlinger, M.; Heintel, D.; Hopfinger, G.; Burgstaller, S.; Mannhalter, C.; Oberaigner, W.; Porpaczy, E.; Skrabs, C.; et al. Rituximab serum concentrations during immuno-chemotherapy of follicular lymphoma correlate with patient gender, bone marrow infiltration and clinical response. Haematologica 2012, 97, 1431–1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, X.; Lu, X.; Guo, J.; Teng, G.J. Interventional therapy combined with immunecheckpoint inhibitors: Emerging opportunities for cancer treatment in the era of immunotherapy. Cancer Treat. Rev. 2019, 74, 49–60. [Google Scholar] [CrossRef]
- Pinto, J.A.; Vallejos, C.S.; Raez, L.E.; Mas, L.A.; Ruiz, R.; Torres-Roman, J.S.; Morante, Z.; Araujo, J.M.; Gomez, H.L.; Aguilar, A.; et al. Gender and outcomes in non-small cell lung cancer: An old prognostic variable comes back for targeted therapy and immunotherapy? ESMO Open 2018, 3, e000344. [Google Scholar] [CrossRef] [Green Version]
- Lin, P.Y.; Sun, L.; Thibodeaux, S.R.; Ludwig, S.M.; Vadlamudi, R.K.; Hurez, V.J.; Bahar, R.; Kious, M.J.; Livi, C.B.; Wall, S.R.; et al. B7-H1-dependent sex-related differences in tumor immunity and immunotherapy responses. J. Immunol. 2010, 185, 2747–2753. [Google Scholar] [CrossRef]
- Wallis, C.J.D.; Butaney, M.; Satkunasivam, R.; Freedland, S.J.; Patel, S.P.; Hamid, O.; Pal, S.K.; Klaassen, Z. Association of Patient Sex With Efficacy of ImmuneCheckpoint Inhibitors and Overall Survival in Advanced Cancers: A Systematic Review and Meta-analysis. JAMA Oncol. 2019, 5, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Hassler, M.R.; Abufaraj, M.; Kimura, S.; Stangl-Kremser, J.; Gust, K.; Glybochko, P.V.; Schmidinger, M.; Karakiewicz, P.I.; Shariat, S.F. Impact of Patients’ Gender on Efficacy of Immunotherapy in Patients with Metastatic Kidney Cancer: A Systematic Review and Meta-analysis. Clin. Genitourin. Cancer 2020, 18, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Rahman, O. Does a patient’s sex predict the efficacy of cancer immunotherapy? Lancet Oncol. 2018, 19, 716–717. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, J.; He, Z.; Wu, K.; Liu, X.S. The predictive power of tumor mutational burden in lung cancer immunotherapy response is influenced by patients’ sex. Int. J. Cancer 2019, 145, 2840–2849. [Google Scholar] [CrossRef]
- Xu, H.; Cao, D.; He, A.; Ge, W. The prognostic role of obesity is independent of sex in cancer patients treated with immunecheckpoint inhibitors: A pooled analysis of 4090 cancer patients. Int. Immunopharmacol. 2019, 74, 105745. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, B.C.; Guibert, N. Immunotherapy efficacy and gender: Discovery in precision medicine. Transl. Lung Cancer Res. 2018, 7, S211–S213. [Google Scholar] [CrossRef]
- Abdel-Rahman, O. Smoking and EGFR status may predict outcomes of advanced NSCLC treated with PD-(L)1 inhibitors beyond firstline: A meta-analysis. Clin. Respir. J. 2018, 12, 1809–1819. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, H.S.; Kim, B.J. Prognostic value of smoking status in non-small-cell lung cancer patients treated with immunecheckpoint inhibitors: A meta-analysis. Oncotarget 2017, 8, 93149–93155. [Google Scholar] [CrossRef] [Green Version]
- Bellmunt, J.; de Wit, R.; Vaughn, D.J.; Fradet, Y.; Lee, J.L.; Fong, L.; Vogelzang, N.J.; Climent, M.A.; Petrylak, D.P.; Choueiri, T.K.; et al. Pembrolizumab as Second-Line Therapy for Advanced Urothelial Carcinoma. N. Engl. J. Med. 2017, 376, 1015–1026. [Google Scholar] [CrossRef] [Green Version]
- Schmid, P.; Adams, S.; Rugo, H.S.; Schneeweiss, A.; Barrios, C.H.; Iwata, H.; Diéras, V.; Hegg, R.; Im, S.A.; Shaw Wright, G.; et al. Atezolizumab and Nab-Paclitaxel in Advanced Triple-Negative Breast Cancer. N. Engl. J. Med. 2018, 379, 2108–2121. [Google Scholar] [CrossRef]
- Calles, A.; Riess, J.W.; Brahmer, J.R. Checkpoint Blockade in Lung Cancer with Driver Mutation: Choose the Road Wisely. Am. Soc. Clin. Oncol. Educ. Book 2020, 40, 372–384. [Google Scholar] [CrossRef] [PubMed]
- Arab, A.; Yazdian-Robati, R.; Behravan, J. HER2-Positive Breast Cancer Immunotherapy: A Focus on Vaccine Development. Arch. Immunol. Ther. Exp. (Warsz) 2020, 68, 2. [Google Scholar] [CrossRef] [PubMed]
- Aldous, A.R.; Dong, J.Z. Personalized neoantigen vaccines: A new approach to cancer immunotherapy. Bioorg. Med. Chem. 2018, 26, 2842–2849. [Google Scholar] [CrossRef] [PubMed]
- Mandal, R.; Chan, T.A. Personalized Oncology Meets Immunology: The Path toward Precision Immunotherapy. Cancer Discov. 2016, 6, 703–713. [Google Scholar] [CrossRef] [Green Version]
- Berghella, A.M.; Contasta, I.; Lattanzio, R.; DiGregorio, G.; Campitelli, I.; Silvino, M.; Liberatore, L.L.; Navarra, L.; Caterino, G.; Mongelli, A.; et al. The Role of Gender-specific Cytokine Pathways as Drug Targets and Gender-specific Biomarkers in Personalized Cancer Therapy. Curr. Drug Targets 2017, 18, 485–495. [Google Scholar] [CrossRef]
- Lotter, H.; Altfeld, M. Sex differences in immunity. Semin. Immunopathol. 2019, 41, 133–135. [Google Scholar] [CrossRef] [Green Version]
- Fischinger, S.; Boudreau, C.M.; Butler, A.L.; Streeck, H.; Alter, G. Sex differences in vaccine-induced humoral immunity. Semin. Immunopathol. 2019, 41, 239–249. [Google Scholar] [CrossRef] [Green Version]
- GubbelsBupp, M.R.; Potluri, T.; Fink, A.L.; Klein, S.L. The Confluence of Sex Hormones and Aging on Immunity. Front. Immunol. 2018, 9, 1269. [Google Scholar] [CrossRef]
- Özdemir, B.C.; Dotto, G.P. Sex Hormones and Anticancer Immunity. Clin. Cancer Res. 2019, 25, 4603–4610. [Google Scholar] [CrossRef] [Green Version]
- Irelli, A.; Sirufo, M.M.; Morelli, L.; D’Ugo, C.; Ginaldi, L.; De Martinis, M. Neuroendocrine Cancer of the Breast: A Rare Entity. J. Clin. Med. 2020, 9, 1452. [Google Scholar] [CrossRef]
- Jaillon, S.; Berthenet, K.; Garlanda, C. Sexual Dimorphism in Innate Immunity. Clin. Rev. Allergy Immunol. 2019, 56, 308–321. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Cowley, L.A.; Liu, X.S. Sex Differences in Cancer Immunotherapy Efficacy, Biomarkers, and Therapeutic Strategy. Molecule 2019, 24, 3214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, S.L. The effects of hormones on sex differences in infection: From genes to behavior. Neurosci. Biobehav. Rev. 2000, 24, 627–638. [Google Scholar] [CrossRef]
- Sánchez-Rodríguez, C.; Cruces, K.P.; RiestraAyora, J.; Martín-Sanz, E.; Sanz-Fernández, R. BCG immune activation reduces growth and angiogenesis in an in vitro model of head and neck squamous cell carcinoma. Vaccine 2017, 35, 6395–6403. [Google Scholar] [CrossRef] [PubMed]
- de Bree, L.C.J.; Janssen, R.; Aaby, P.; van Crevel, R.; Joosten, L.A.B.; Benn, C.S.; Netea, M.G. The impact of sex hormones on BCG-induced trained immunity. J. Leukoc. Biol. 2018, 104, 573–578. [Google Scholar] [CrossRef]
- Pampena, M.B.; Cartar, H.C.; Cueto, G.R.; Levy, E.M.; Blanco, P.A.; Barrio, M.M.; Mordoh, J. Dissecting the Immune Stimulation Promoted by CSF-470 Vaccine Plus Adjuvants in Cutaneous Melanoma Patients: Long Term Antitumor Immunity and Short Term Release of Acute Inflammatory Reactants. Front. Immunol. 2018, 2. [Google Scholar] [CrossRef]
- Uhlig, A.; Strauss, A.; SeifAmirHosseini, A.; Lotz, J.; Trojan, L.; Schmid, M.; Uhlig, J. Gender-specific Differences in Recurrence of Non-muscle-invasive Bladder Cancer: A Systematic Review and Meta-analysis. Eur. Urol. Focus 2018, 4, 924–936. [Google Scholar] [CrossRef]
- Sirufo, M.M.; De Martinis, M.; Ginaldi, L. Omalizumab an effective and safe alternative therapy in severe refractory atopicdermatitis. Acase report. Medicine 2018, 97, e10897. [Google Scholar] [CrossRef]
- De Martinis, M.; Sirufo, M.M.; Ginaldi, L. A “Stadium” Urticaria, Cold Urticaria is Still a Mostly Unknown Disease, with a Wide Spectrum of Severity Degrees and Few Therapeutic Certainties: Is Omalizumab one of These? Reflections from a Clinical Case Report. Iran. Red Crescent Med. J. 2019, 21, 84250. [Google Scholar] [CrossRef]
- De Martinis, M.; Sirufo, M.M.; Ginaldi, L. Solar urticaria, a disease with many dark sides: Is omalizumab the right therapeutic response? Reflections from a clinical case report. Open Med. 2019, 14, 403–406. [Google Scholar] [CrossRef] [Green Version]
- Sirufo, M.M.; Ginaldi, L.; De Martinis, M. Asthma, urticaria and omalizumab in children: Reflections from a clinical case report. Front. Pediatr. 2019, 7, 213. [Google Scholar] [CrossRef] [PubMed]
- De Martinis, M.; Sirufo, M.M.; Ginaldi, L. Food allergy and ageing. Int. J. Mol. Sci. 2019, 20, 5580. [Google Scholar] [CrossRef] [Green Version]
- De Martinis, M.; Sirufo, M.M.; Ginaldi, L. Food allergy insights: A changing landscape. Arch. Immunol. Ther. Exp. 2020, 68, 8. [Google Scholar] [CrossRef] [PubMed]
- De Martinis, M.; Sirufo, M.M.; De Pietro, F.; Bassino, E.M.; Ginaldi, L.; De Martinis, M. Osteoporosis in Skin Diseases. Int. J. Mol. Sci. 2020, 21, 4749. [Google Scholar] [CrossRef]
- Jendoubi, F.; Gaudenzio, N.; Gallini, A.; Negretto, M.; Paul, C.; BulaiLivideanu, C. Omalizumab in the treatment of adult patients with mastocytosis: A systematic review. Clin. Exp. Allergy 2020, 50, 654–661. [Google Scholar] [CrossRef] [PubMed]
- Gouder, C.; Asciak, R.; Montefort, S. Sex differences in the efficacy, safety, and tolerability of omalizumab after 1 year in Maltese patients with asthma. Ann. Allergy Asthma Immunol. 2017, 118, 513–514. [Google Scholar] [CrossRef] [PubMed]
- Sirufo, M.M.; De Pietro, F.; Bassino, E.M.; Ginaldi, L.; De Martinis, M. Translational allergy and omalizumab: The pioneer. Indian J. Pharm. Ed. Res. 2020. submitted. [Google Scholar]
- Iwai, K.; Soejima, K.; Kudoh, S.; Umezato, Y.; Kaneko, T.; Yoshimori, K.; Tokuda, H.; Yamaguchi, T.; Mizoo, A.; Setoguchi, Y.; et al. Extended survival observed in adoptive activated T lymphocyte immunotherapy for advanced lung cancer: Results of a multicenter historical cohort study. Cancer Immunol. Immunother. 2012, 61, 1781–1790. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.W.; Asakura, Y.; Tajima, K.; Iwai, T.; Taji, H.; Chou, T.; Morishima, Y.; Suzumiya, J.; Sakamaki, H.; Suzuki, R.; et al. High-dose therapy and autologous stemcell transplantation for relapsed or high-risk diffuse large B-cell lymphoma: A nationwide survey. Int. J. Hematol. 2020, 111, 256–266. [Google Scholar] [CrossRef]
- Kim, H.I.; Lim, H.; Moon, A. Sex Differences in Cancer: Epidemiology, Genetics and Therapy. Biomol. Ther. 2018, 26, 335–342. [Google Scholar] [CrossRef]
- Gratwohl, A.; RuizdeElvira, C.; Gratwohl, M.; Greinix, H.T.; Duarte, R. Graft-versus-Host Disease Subcommittee of the Complications and Quality of Life Working Party of the European Society for Blood and Marrow Transplantation. Gender and Graft-versus-Host Disease after Hematopoietic Stem Cell Transplantation. Biol. Blood Marrow Transplant. 2016, 22, 1145–1146. [Google Scholar] [CrossRef] [Green Version]
- Nakasone, H.; Kawamura, K.; Yakushijin, K.; Shinohara, A.; Tanaka, M.; Ohashi, K.; Ota, S.; Uchida, N.; Fukuda, T.; Nakamae, H.; et al. B Mispreferred over PBSCs in transplantation from an HLA-matched related female donor to a male recipient. Blood Adv. 2019, 3, 1750–1760. [Google Scholar] [CrossRef] [PubMed]
- Ciurea, S.O.; AlMalki, M.M.; Kongtim, P.; Fuchs, E.J.; Luznik, L.; Huang, X.J.; Ciceri, F.; Locatelli, F.; Aversa, F.; Castagna, L. The European Society for Blood and Marrow Transplantation (EBMT) consensus recommendations for donor selection in haploidentical hematopoietic cell transplantation. Bone Marrow Transplant. 2020, 55, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Takekiyo, T.; Dozono, K.; Nara, S.; Murayama, Y.; Minamihama, N.; Nakano, N.; Kubota, A.; Tokunaga, M.; Miyazono, T.; Takeuchi, S.; et al. Gender differences in physical function and muscle mass change in patients undergoing allogeneic hematopoietic stemcell transplantation. Bone Marrow Transplant. 2017, 52, 1460–1462. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, N.S.; Brazauskas, R.; Tecca, H.R.; Vogel, J.; Mattila, D.; Lee, S.J.; Horowitz, M.M.; Rizzo, J.D.; Shaw, B.E. Female Sex is Associated With Poor Health-related Quality of Life in Children at 12 Months Post-Hematopoietic Cell Transplantation. J. Pediatr. Hematol. Oncol. 2019, 41, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Volk, V.; Schneider, A.; Spineli, L.M.; Grosshennig, A.; Stripecke, R. The gender gap: Discrepant humanT-cell reconstitution after cordblood stemcell transplantation in humanized female and male mice. Bone Marrow Transplant. 2016, 51, 596–597. [Google Scholar] [CrossRef] [Green Version]
- Lafta, I.J.; Bryant, H.E.; Goldman, A.S. ‘Sex’ in the cancer cell. Oncotarget 2014, 5, 7984–7985. [Google Scholar] [CrossRef]
- Shin, J.Y.; Jung, H.J.; Moon, A. Molecular Markers in Sex Differences in Cancer. Toxicol. Res. 2019, 35, 331–341. [Google Scholar] [CrossRef] [Green Version]
- Colli, L.M.; Morton, L.M.; Chanock, S.J. Sex-Related Effect on Immunotherapy Response: Implications and Opportunities. J. Natl. Cancer Inst. 2019, 111, 749–750. [Google Scholar] [CrossRef]
- Díaz, A.; Beleña, Á.; Zueco, J. The Role of Age and Gender in Perceived Vulnerability to Infectious Diseases. Int. J. Environ. Res. Public Health 2020, 11, 485. [Google Scholar] [CrossRef] [Green Version]
- Klein, S.L.; Morgan, R. The impact of sex and gender on immunotherapy outcomes. Biol. Sex. Differ. 2020, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, B.C.; Rowe, A.K.; Crawford-Williams, F.; Baade, P.; Chambers, S.K.; Ralph, N.; Aitken, J.F. Geographical Disparities in Screening and Cancer-Related Health Behaviour. Int. J. Environ. Res. Public Health 2020, 17, 1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Short, S.E.; Yang, Y.C.; Jenkins, T.M. Sex, gender, genetics, and health. Am. J. Public Health 2013, 103, S93–S101. [Google Scholar] [CrossRef]
- Campostrini, S.; Carrozzi, G.; Severoni, S.; Masocco, M.; Salmaso, S.; WHO Migration Health Programme; Office of the Regional Director; WHO Regional Office for Europe; PASSI National Coordinating Group. Migrant Health in Italy: A Better Health Status Difficult to Maintain-Country of Origin and Assimilation Effects Studied From the Italian Risk Factor Surveillance Data. Popul. Health Metr. 2019, 17, 14. [Google Scholar] [CrossRef]
- Norum, J.; Nieder, C. Tobacco Smoking and Cessation and PD-L1 Inhibitorsin Non-Small Cell Lung Cancer (NSCLC): A Review of the Literature. ESMO Open 2018, 3, e000406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wessels, H.; de Graeff, A.; Wynia, K.; de Heus, M.; Kruitwagen, C.L.; Woltjer, G.T.; Teunissen, S.C.; Voest, E.E. Gender-related needs and preferences in cancer care indicate the need for an individualized approach to cancer patients. Oncologist 2010, 15, 648–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinninella, E.; Cintoni, M.; Raoul, P.; Lopetuso, L.R.; Scaldaferri, F.; Pulcini, G.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. Food Components and Dietary Habits: Keys for a Healthy Gut Microbiota Composition. Nutrients 2019, 11, 2393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cani, P.D.; Jordan, B.F. Gut microbiota-mediated inflammation in obesity: A link with gastrointestinal cancer. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 671–682. [Google Scholar] [CrossRef]
- Ajaj, R.; Cáceres, J.O.H.; Berlin, A.; Wallis, C.J.D.; Chandrasekar, T.; Klaassen, Z.; Ahmad, A.E.; Leao, R.; Finelli, A.; Fleshner, N.; et al. Gender-based psychological and physical distress differences in patients diagnosed with non-metastatic renal cell carcinoma. World J. Urol. 2020, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bergerot, C.D.; Battle, D.; Staehler, M.D.; Pal, S.K. Distress in patients with renal cell carcinoma: A curious gap in knowledge. Bju Int. 2018, 123, 208–209. [Google Scholar] [CrossRef] [Green Version]
- Mancini, M.; Righetto, M.; Baggio, G. Gender-Related Approach to Kidney Cancer Management: Moving Forward. Int. J. Mol. Sci. 2020, 21, 3378. [Google Scholar] [CrossRef]
- Shin, H.; Bartlett, R.; DeGagne, J.C. Health-Related Quality of Life among Survivors of Cancer in Adolescence: An Integrative Literature Review. J. Pediatr. Nurs. 2019, 44, 97–106. [Google Scholar] [CrossRef]
- Westby, R.P.; Berg, C.J.; Leach, C. Gender, race, BMI, and social support in relation to the health-related quality of life of cancer survivors: Are port from the American Cancer Society’s Study of Cancer Survivors II (SCS-II). Qual. Life Res. 2016, 25, 409–421. [Google Scholar] [CrossRef]
- Inderberg, E.M.; Wälchli, S. Sympathetic improvement of cancer vaccine efficacy. Hum. Vaccines Immunother. 2020, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFarland, D.C. New lung cancer treatments (immunotherapy and targeted therapies) and their associations with depression and other psychological side effects as compared to chemotherapy. Gen. Hosp. Psychiatr. 2019, 60, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.; Kim, S.Y.; Lee, E.; Lee, S.; Oh, S.; Jung, J.; Kim, K.S.; Moon, A. Sex-Dependent Adverse Drug Reactions to 5-Fluorouracil in Colorectal Cancer. Biol. Pharm. Bull. 2019, 42, 594–600. [Google Scholar] [CrossRef] [Green Version]
- Duma, N.; Abdel-Ghani, A.; Yadav, S.; Hoversten, K.P.; Reed, C.T.; Sitek, A.N.; Enninga, E.A.L.; Paludo, J.; Aguilera, J.V.; Leventakos, K.; et al. Sex Differences in Tolerability to Anti-Programmed Cell Death Protein1 Therapy in Patients with Metastatic Melanoma and Non-Small Cell Lung Cancer: Are We All Equal? Oncologist 2019, 24, e1148–e1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrera, C.; Potrony, M.; Puig, S. Sex as a predictor of response to cancer immunotherapy. Lancet Oncol. 2018, 19, e375. [Google Scholar] [CrossRef]
- Clarke, M.; Lewin, J.; Lazarakis, S.; Thompson, K. Overlooked Minorities: The Intersection of Cancer in Lesbian, Gay, Bisexual, Transgender, and/or Intersex Adolescents and Young Adults. J. Adolesc. Young Adult Oncol. 2019, 8, 525–528. [Google Scholar] [CrossRef]
- Tabaac, A.R.; Sutter, M.E.; Wall, C.S.J.; Baker, K.E. Gender Identity Disparities in Cancer Screening Behaviors. Am. J. Prev. Med. 2018, 54, 385–393. [Google Scholar] [CrossRef]

| ESTROGENS | |
|---|---|
| HIGH | LOW |
| Increased Treg | Decreased Treg |
| Th2 differentiation | Th1 differentiation |
| Reduced monocytes secretion of IL-1β, IL-6 and TNF | Increased monocytes secretion of IL-1β, IL-6 and TNF |
| Increased chemotactic capacity of neutrophils | Reduced chemotactic capacity of neutrophils |
| Mitigated cytotoxicity of natural killer (NK) cells | Stimulated cytotoxicity of natural killer (NK) cells |
| Increased differentiation and activation of dendritic cells | Reduced action on dendritic cells |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Irelli, A.; Sirufo, M.M.; D’Ugo, C.; Ginaldi, L.; De Martinis, M. Sex and Gender Influences on Cancer Immunotherapy Response. Biomedicines 2020, 8, 232. https://doi.org/10.3390/biomedicines8070232
Irelli A, Sirufo MM, D’Ugo C, Ginaldi L, De Martinis M. Sex and Gender Influences on Cancer Immunotherapy Response. Biomedicines. 2020; 8(7):232. https://doi.org/10.3390/biomedicines8070232
Chicago/Turabian StyleIrelli, Azzurra, Maria Maddalena Sirufo, Carlo D’Ugo, Lia Ginaldi, and Massimo De Martinis. 2020. "Sex and Gender Influences on Cancer Immunotherapy Response" Biomedicines 8, no. 7: 232. https://doi.org/10.3390/biomedicines8070232