
Age-Dependent Dysregulation of Muscle Vasculature and Blood Flow Recovery after Hindlimb Ischemia in the mdx Model of Duchenne Muscular Dystrophy

 and
and
Abstract
:1. Introduction
2. Materials and Method
2.1. Animal Models
2.2. Blood Cell Count
2.3. In Vitro Angiogenesis Assay
2.4. Forelimb Grip Strength Assessment
2.5. HLI Induction
2.6. Generation and Differentiation of Human-Induced Pluripotent Stem Cells (hiPSCs)-Derived Skeletal Muscle Cells
2.7. RNA Isolation, Reverse Transcription (RT), and Quantitative Real-Time PCR (qRT-PCR)
2.8. Histological and Immunofluorescent Analysis of Muscles
2.9. Determination of Serum Creatine Kinase (CK) and Lactate Dehydrogenase (LDH) Activity
2.10. Enzyme-Linked Immunosorbent Assay (ELISA)
2.11. Flow Cytometry Analysis
2.12. Statistical Analyses
3. Results
3.1. Alterations in Angiogenesis Markers Are Present in Skeletal Muscles of 6- and 12-Week-Old Dystrophic Mice
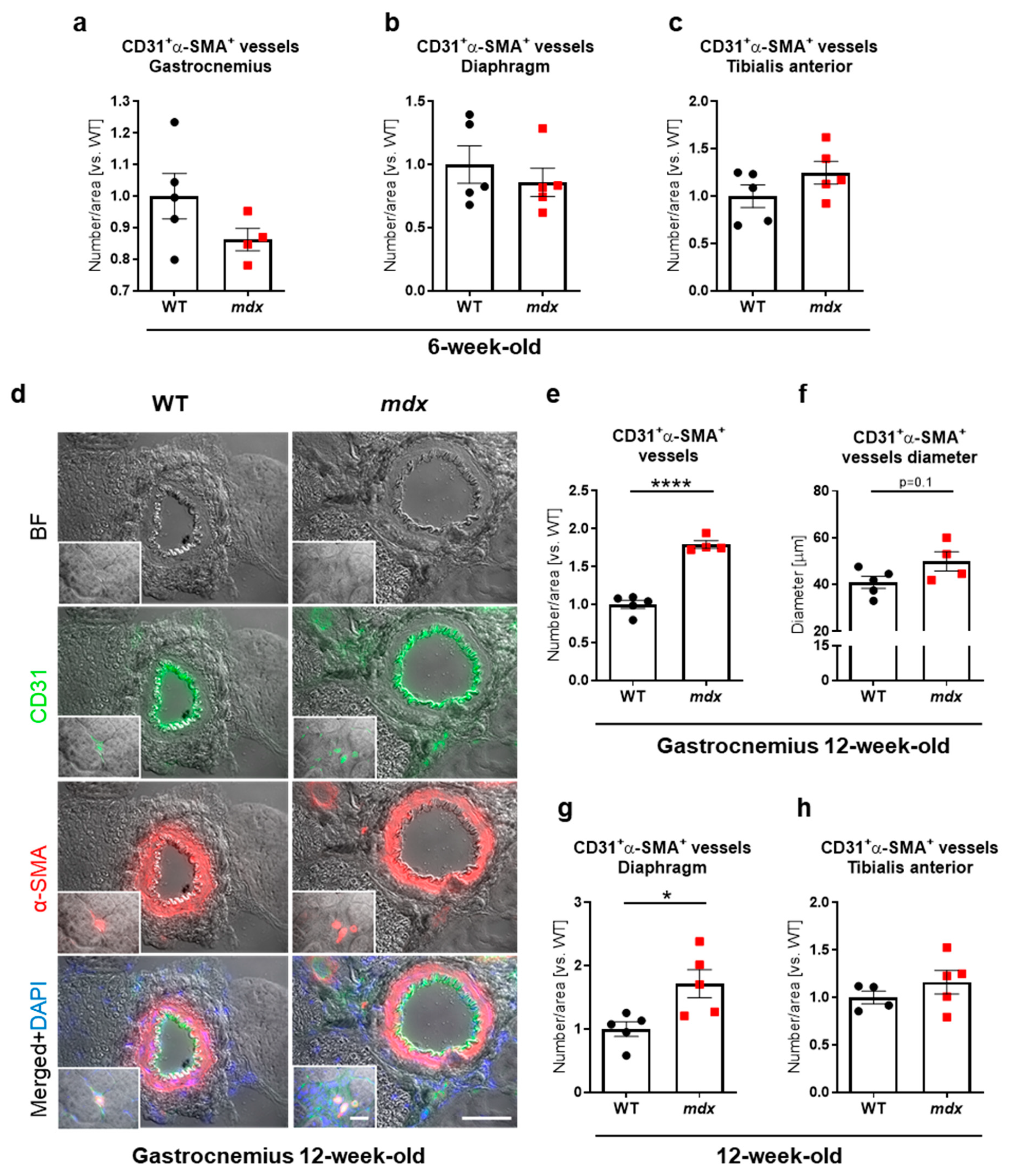
3.2. Increased Number of Blood Vessels Is Predominantly Observed in Skeletal Muscles of 12-Week-Old, but Not 6-Week-Old Dystrophic Animals
3.3. Basal Blood Perfusion and Revascularisation in Response to HLI Are Decreased in 12-Week-Old, but Not 6-Week-Old mdx Mice
3.4. HLI-Induced Inflammatory Response Is Impaired in 12-Week-Old Dystrophic Mice
3.5. 12-Week-Old Dystrophic Mice Failed to Upregulate a Regenerative and Tissue Remodeling Response to HLI
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carter, J.C.; Sheehan, D.W.; Prochoroff, A.; Birnkrant, D.J. Muscular Dystrophies. Clin. Chest Med. 2018, 39, 377–389. [Google Scholar] [CrossRef]
- Bladen, C.L.; Salgado, D.; Monges, S.; Foncuberta, M.E.; Kekou, K.; Kosma, K.; Dawkins, H.; Lamont, L.; Roy, A.J.; Chamova, T.; et al. The TREAT-NMD DMD Global Database: Analysis of More than 7,000 Duchenne Muscular Dystrophy Mutations. Hum. Mutat. 2015, 36, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Campbell, K.P.; Kahl, S.D. Association of Dystrophin and an Integral Membrane Glycoprotein. Nature 1989, 338, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Landfeldt, E.; Thompson, R.; Sejersen, T.; McMillan, H.J.; Kirschner, J.; Lochmüller, H. Life Expectancy at Birth in Duchenne Muscular Dystrophy: A Systematic Review and Meta-Analysis. Eur. J. Epidemiol 2020, 35, 643–653. [Google Scholar] [CrossRef] [Green Version]
- Łoboda, A.; Dulak, J. Muscle and Cardiac Therapeutic Strategies for Duchenne Muscular Dystrophy: Past, Present, and Future. Pharmacol. Rep. 2020, 72, 1227–1263. [Google Scholar] [CrossRef]
- Nichols, B.; Takeda, S.; Yokota, T. Nonmechanical Roles of Dystrophin and Associated Proteins in Exercise, Neuromuscular Junctions, and Brains. Brain Sci. 2015, 5, 275–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumont, N.A.; Wang, Y.X.; von Maltzahn, J.; Pasut, A.; Bentzinger, C.F.; Brun, C.E.; Rudnicki, M.A. Dystrophin Expression in Muscle Stem Cells Regulates Their Polarity and Asymmetric Division. Nat. Med. 2015, 21, 1455–1463. [Google Scholar] [CrossRef] [Green Version]
- Hugnot, J.P.; Gilgenkrantz, H.; Chafey, P.; Lambert, M.; Eveno, E.; Kaplan, J.C.; Kahn, A. Expression of the Dystrophin Gene in Cultured Fibroblasts. Biochem. Biophys Res. Commun. 1993, 192, 69–74. [Google Scholar] [CrossRef]
- Harricane, M.-C.; Febris, E.; Lees, D.; Prades, C.; Travo, P.; Mornet, D. Dystrophin Does Not Influence Regular Cytoskeletal Architecture but Is Required for Contractile Performance in Smooth Muscle Aortic Cells. Cell Biol. Int. 1994, 18, 947–958. [Google Scholar] [CrossRef]
- Loufrani, L.; Dubroca, C.; You, D.; Li, Z.; Levy, B.; Paulin, D.; Henrion, D. Absence of Dystrophin in Mice Reduces NO-Dependent Vascular Function and Vascular Density: Total Recovery after a Treatment with the Aminoglycoside Gentamicin. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 671–676. [Google Scholar] [CrossRef] [Green Version]
- Podkalicka, P.; Mucha, O.; Dulak, J.; Loboda, A. Targeting Angiogenesis in Duchenne Muscular Dystrophy. Cell. Mol. Life Sci. 2019, 76, 1507–1528. [Google Scholar] [CrossRef] [Green Version]
- Scholz, D.; Thomas, S.; Sass, S.; Podzuweit, T. Angiogenesis and Myogenesis as Two Facets of Inflammatory Post-Ischemic Tissue Regeneration. Mol. Cell Biochem. 2003, 246, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Tahergorabi, Z.; Khazaei, M. A Review on Angiogenesis and Its Assays. Iran. J. Basic Med. Sci. 2012, 15, 1110–1126. [Google Scholar]
- Straino, S.; Germani, A.; Di Carlo, A.; Porcelli, D.; De Mori, R.; Mangoni, A.; Napolitano, M.; Martelli, F.; Biglioli, P.; Capogrossi, M.C. Enhanced Arteriogenesis and Wound Repair in Dystrophin-Deficient Mdx Mice. Circulation 2004, 110, 3341–3348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palladino, M.; Gatto, I.; Neri, V.; Straino, S.; Smith, R.C.; Silver, M.; Gaetani, E.; Marcantoni, M.; Giarretta, I.; Stigliano, E.; et al. Angiogenic Impairment of the Vascular Endothelium: A Novel Mechanism and Potential Therapeutic Target in Muscular Dystrophy. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 2867–2876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latroche, C.; Matot, B.; Martins-Bach, A.; Briand, D.; Chazaud, B.; Wary, C.; Carlier, P.G.; Chrétien, F.; Jouvion, G. Structural and Functional Alterations of Skeletal Muscle Microvasculature in Dystrophin-Deficient Mdx Mice. Am. J. Pathol. 2015, 185, 2482–2494. [Google Scholar] [CrossRef] [PubMed]
- Dangain, J.; Vrbova, G. Muscle Development in Mdx Mutant Mice. Muscle Nerve 1984, 7, 700–704. [Google Scholar] [CrossRef]
- DiMario, J.X.; Uzman, A.; Strohman, R.C. Fiber Regeneration Is Not Persistent in Dystrophic (MDX) Mouse Skeletal Muscle. Dev. Biol 1991, 148, 314–321. [Google Scholar] [CrossRef]
- Banks, G.B.; Chamberlain, J.S. The Value of Mammalian Models for Duchenne Muscular Dystrophy in Developing Therapeutic Strategies. Curr. Top. Dev. Biol. 2008, 84, 431–453. [Google Scholar] [CrossRef] [PubMed]
- Massopust, R.T.; Lee, Y.I.; Pritchard, A.L.; Nguyen, V.-K.M.; McCreedy, D.A.; Thompson, W.J. Lifetime Analysis of Mdx Skeletal Muscle Reveals a Progressive Pathology That Leads to Myofiber Loss. Sci. Rep. 2020, 10, 17248. [Google Scholar] [CrossRef]
- Ennen, J.P.; Verma, M.; Asakura, A. Vascular-Targeted Therapies for Duchenne Muscular Dystrophy. Skelet Muscle 2013, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Podkalicka, P.; Mucha, O.; Bronisz-Budzyńska, I.; Kozakowska, M.; Pietraszek-Gremplewicz, K.; Cetnarowska, A.; Głowniak-Kwitek, U.; Bukowska-Strakova, K.; Cieśla, M.; Kulecka, M.; et al. Lack of MiR-378 Attenuates Muscular Dystrophy in Mdx Mice. JCI Insight 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Bronisz-Budzyńska, I.; Kozakowska, M.; Podkalicka, P.; Kachamakova-Trojanowska, N.; Łoboda, A.; Dulak, J. The Role of Nrf2 in Acute and Chronic Muscle Injury. Skelet Muscle 2020, 10, 35. [Google Scholar] [CrossRef] [PubMed]
- Bronisz-Budzyńska, I.; Chwalenia, K.; Mucha, O.; Podkalicka, P.; Bukowska-Strakova, K.; Józkowicz, A.; Łoboda, A.; Kozakowska, M.; Dulak, J. MiR-146a Deficiency Does Not Aggravate Muscular Dystrophy in Mdx Mice. Skelet Muscle 2019, 9, 22. [Google Scholar] [CrossRef] [PubMed]
- Pietraszek-Gremplewicz, K.; Kozakowska, M.; Bronisz-Budzynska, I.; Ciesla, M.; Mucha, O.; Podkalicka, P.; Madej, M.; Glowniak, U.; Szade, K.; Stepniewski, J.; et al. Heme Oxygenase-1 Influences Satellite Cells and Progression of Duchenne Muscular Dystrophy in Mice. Antioxid. Redox Signal. 2018, 29, 128–148. [Google Scholar] [CrossRef]
- Shin, J.-H.; Hakim, C.H.; Zhang, K.; Duan, D. Genotyping Mdx, Mdx3cv, and Mdx4cv Mice by Primer Competition Polymerase Chain Reaction. Muscle Nerve 2011, 43, 283–286. [Google Scholar] [CrossRef] [Green Version]
- Carpentier, G.; Berndt, S.; Ferratge, S.; Rasband, W.; Cuendet, M.; Uzan, G.; Albanese, P. Angiogenesis Analyzer for ImageJ—A Comparative Morphometric Analysis of “Endothelial Tube Formation Assay” and “Fibrin Bead Assay”. Sci. Rep. 2020, 10, 11568. [Google Scholar] [CrossRef]
- Niiyama, H.; Huang, N.F.; Rollins, M.D.; Cooke, J.P. Murine Model of Hindlimb Ischemia. J. Vis. Exp. 2009. [Google Scholar] [CrossRef]
- Krist, B.; Podkalicka, P.; Mucha, O.; Mendel, M.; Sępioł, A.; Rusiecka, O.M.; Józefczuk, E.; Bukowska-Strakova, K.; Grochot-Przęczek, A.; Tomczyk, M.; et al. MiR-378a Influences Vascularization in Skeletal Muscles. Cardiovasc. Res. 2020, 116, 1386–1397. [Google Scholar] [CrossRef]
- Stępniewski, J.; Tomczyk, M.; Andrysiak, K.; Kraszewska, I.; Martyniak, A.; Langrzyk, A.; Kulik, K.; Wiśniewska, E.; Jeż, M.; Florczyk-Soluch, U.; et al. Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes, in Contrast to Adipose Tissue-Derived Stromal Cells, Efficiently Improve Heart Function in Murine Model of Myocardial Infarction. Biomedicines 2020, 8, 578. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. Single-Step Method of RNA Isolation by Acid Guanidinium Thiocyanate-Phenol-Chloroform Extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Kozakowska, M.; Pietraszek-Gremplewicz, K.; Ciesla, M.; Seczynska, M.; Bronisz-Budzynska, I.; Podkalicka, P.; Bukowska-Strakova, K.; Loboda, A.; Jozkowicz, A.; Dulak, J. Lack of Heme Oxygenase-1 Induces Inflammatory Reaction and Proliferation of Muscle Satellite Cells after Cardiotoxin-Induced Skeletal Muscle Injury. Am. J. Pathol. 2018, 188, 491–506. [Google Scholar] [CrossRef] [Green Version]
- Sicinski, P.; Geng, Y.; Ryder-Cook, A.S.; Barnard, E.A.; Darlison, M.G.; Barnard, P.J. The Molecular Basis of Muscular Dystrophy in the Mdx Mouse: A Point Mutation. Science 1989, 244, 1578–1580. [Google Scholar] [CrossRef] [PubMed]
- McGreevy, J.W.; Hakim, C.H.; McIntosh, M.A.; Duan, D. Animal Models of Duchenne Muscular Dystrophy: From Basic Mechanisms to Gene Therapy. Dis. Model. Mech. 2015, 8, 195–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hathout, Y.; Marathi, R.L.; Rayavarapu, S.; Zhang, A.; Brown, K.J.; Seol, H.; Gordish-Dressman, H.; Cirak, S.; Bello, L.; Nagaraju, K.; et al. Discovery of Serum Protein Biomarkers in the Mdx Mouse Model and Cross-Species Comparison to Duchenne Muscular Dystrophy Patients. Hum. Mol. Genet. 2014, 23, 6458–6469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuraoka, M.; Kimura, E.; Nagata, T.; Okada, T.; Aoki, Y.; Tachimori, H.; Yonemoto, N.; Imamura, M.; Takeda, S. Serum Osteopontin as a Novel Biomarker for Muscle Regeneration in Duchenne Muscular Dystrophy. Am. J. Pathol. 2016, 186, 1302–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vetrone, S.A.; Montecino-Rodriguez, E.; Kudryashova, E.; Kramerova, I.; Hoffman, E.P.; Liu, S.D.; Miceli, M.C.; Spencer, M.J. Osteopontin Promotes Fibrosis in Dystrophic Mouse Muscle by Modulating Immune Cell Subsets and Intramuscular TGF-Beta. J. Clin. Investig. 2009, 119, 1583–1594. [Google Scholar] [CrossRef] [Green Version]
- Pagel, C.N.; Wasgewatte, W.D.K.; Taghavi, E.N.; Mackie, E.J. Osteopontin, Inflammation and Myogenesis: Influencing Regeneration, Fibrosis and Size of Skeletal Muscle. J. Cell Commun. Signal. 2014, 8, 95–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, J.; Peng, L.; Fan, K.; Wang, H.; Wei, R.; Ji, G.; Cai, J.; Lu, B.; Li, B.; Zhang, D.; et al. Osteopontin Induces Angiogenesis through Activation of PI3K/AKT and ERK1/2 in Endothelial Cells. Oncogene 2009, 28, 3412–3422. [Google Scholar] [CrossRef] [Green Version]
- Woodfin, A.; Voisin, M.-B.; Nourshargh, S. PECAM-1: A Multi-Functional Molecule in Inflammation and Vascular Biology. Arterioscler. Thromb Vasc. Biol. 2007, 27, 2514–2523. [Google Scholar] [CrossRef] [Green Version]
- Watt, S.M.; Gschmeissner, S.E.; Bates, P.A. PECAM-1: Its Expression and Function as a Cell Adhesion Molecule on Hemopoietic and Endothelial Cells. Leuk. Lymphoma 1995, 17, 229–244. [Google Scholar] [CrossRef]
- Shireman, P.K. The Chemokine System in Arteriogenesis and Hind Limb Ischemia. J. Vasc. Surg. 2007, 45, A48–A56. [Google Scholar] [CrossRef] [Green Version]
- Rossi, E.; Smadja, D.; Goyard, C.; Cras, A.; Dizier, B.; Bacha, N.; Lokajczyk, A.; Guerin, C.L.; Gendron, N.; Planquette, B.; et al. Co-Injection of Mesenchymal Stem Cells with Endothelial Progenitor Cells Accelerates Muscle Recovery in Hind Limb Ischemia through an Endoglin-Dependent Mechanism. Thromb. Haemost. 2017, 117, 1908–1918. [Google Scholar] [CrossRef]
- Füchtbauer, E.-M.; Westphal, H. MyoD and Myogenin Are Coexpressed in Regenerating Skeletal Muscle of the Mouse. Dev. Dyn. 1992, 193, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, S.; Rossi, A.C.; Smerdu, V.; Leinwand, L.A.; Reggiani, C. Developmental Myosins: Expression Patterns and Functional Significance. Skelet Muscle 2015, 5, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, M.; Asakura, Y.; Hirai, H.; Watanabe, S.; Tastad, C.; Fong, G.-H.; Ema, M.; Call, J.A.; Lowe, D.A.; Asakura, A. Flt-1 Haploinsufficiency Ameliorates Muscular Dystrophy Phenotype by Developmentally Increased Vasculature in Mdx Mice. Hum. Mol. Genet. 2010, 19, 4145–4159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, M.; Shimizu-Motohashi, Y.; Asakura, Y.; Ennen, J.P.; Bosco, J.; Zhou, Z.; Fong, G.-H.; Josiah, S.; Keefe, D.; Asakura, A. Inhibition of FLT1 Ameliorates Muscular Dystrophy Phenotype by Increased Vasculature in a Mouse Model of Duchenne Muscular Dystrophy. PLoS Genet. 2019, 15, e1008468. [Google Scholar] [CrossRef] [Green Version]
- Messina, S.; Mazzeo, A.; Bitto, A.; Aguennouz, M.; Migliorato, A.; De Pasquale, M.G.; Minutoli, L.; Altavilla, D.; Zentilin, L.; Giacca, M.; et al. VEGF Overexpression via Adeno-Associated Virus Gene Transfer Promotes Skeletal Muscle Regeneration and Enhances Muscle Function in Mdx Mice. FASEB J. 2007, 21, 3737–3746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, M.; Asakura, Y.; Murakonda, B.S.R.; Pengo, T.; Latroche, C.; Chazaud, B.; McLoon, L.K.; Asakura, A. Muscle Satellite Cell Cross-Talk with a Vascular Niche Maintains Quiescence via VEGF and Notch Signaling. Cell Stem Cell 2018, 23, 530–543. [Google Scholar] [CrossRef] [Green Version]
- Christov, C.; Chrétien, F.; Abou-Khalil, R.; Bassez, G.; Vallet, G.; Authier, F.-J.; Bassaglia, Y.; Shinin, V.; Tajbakhsh, S.; Chazaud, B.; et al. Muscle Satellite Cells and Endothelial Cells: Close Neighbors and Privileged Partners. Mol. Biol. Cell 2007, 18, 1397–1409. [Google Scholar] [CrossRef] [Green Version]
- Brodal, P.; Ingjer, F.; Hermansen, L. Capillary Supply of Skeletal Muscle Fibers in Untrained and Endurance-Trained Men. Am. J. Physiol 1977, 232, H705–H712. [Google Scholar] [CrossRef] [PubMed]
- Waters, R.E.; Rotevatn, S.; Li, P.; Annex, B.H.; Yan, Z. Voluntary Running Induces Fiber Type-Specific Angiogenesis in Mouse Skeletal Muscle. Am. J. Physiol. Cell Physiol. 2004, 287, C1342–C1348. [Google Scholar] [CrossRef] [Green Version]
- Matsakas, A.; Yadav, V.; Lorca, S.; Narkar, V. Muscle ERRγ Mitigates Duchenne Muscular Dystrophy via Metabolic and Angiogenic Reprogramming. FASEB J. 2013, 27, 4004–4016. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, H.; Betsholtz, C. Endothelial-Pericyte Interactions in Angiogenesis. Cell Tissue Res. 2003, 314, 15–23. [Google Scholar] [CrossRef]
- Ieronimakis, N.; Balasundaram, G.; Reyes, M. Direct Isolation, Culture and Transplant of Mouse Skeletal Muscle Derived Endothelial Cells with Angiogenic Potential. PLoS ONE 2008, 3, e0001753. [Google Scholar] [CrossRef] [Green Version]
- Yun, J.-H.; Koh, Y.J.; Jeong, H.-S.; Lee, D.-H.; Lee, E.H.; Cho, C.-H. Propranolol Increases Vascular Permeability through Pericyte Apoptosis and Exacerbates Oxygen-Induced Retinopathy. Biochem. Biophys. Res. Commun. 2018, 503, 2792–2799. [Google Scholar] [CrossRef]
- Sweeney, M.; Foldes, G. It Takes Two: Endothelial-Perivascular Cell Cross-Talk in Vascular Development and Disease. Front. Cardiovasc. Med. 2018, 5, 154. [Google Scholar] [CrossRef]
- Miike, T.; Sugino, S.; Ohtani, Y.; Taku, K.; Yoshioka, K. Vascular Endothelial Cell Injury and Platelet Embolism in Duchenne Muscular Dystrophy at the Preclinical Stage. J. Neurol. Sci. 1987, 82, 67–80. [Google Scholar] [CrossRef]
- Loufrani, L.; Matrougui, K.; Gorny, D.; Duriez, M.; Blanc, I.; Lévy, B.I.; Henrion, D. Flow (Shear Stress)-Induced Endothelium-Dependent Dilation Is Altered in Mice Lacking the Gene Encoding for Dystrophin. Circulation 2001, 103, 864–870. [Google Scholar] [CrossRef]
- Heil, M.; Ziegelhoeffer, T.; Pipp, F.; Kostin, S.; Martin, S.; Clauss, M.; Schaper, W. Blood Monocyte Concentration Is Critical for Enhancement of Collateral Artery Growth. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H2411–H2419. [Google Scholar] [CrossRef] [PubMed]
- Cochain, C.; Rodero, M.P.; Vilar, J.; Récalde, A.; Richart, A.L.; Loinard, C.; Zouggari, Y.; Guérin, C.; Duriez, M.; Combadière, B.; et al. Regulation of Monocyte Subset Systemic Levels by Distinct Chemokine Receptors Controls Post-Ischaemic Neovascularization. Cardiovasc. Res. 2010, 88, 186–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capoccia, B.J.; Gregory, A.D.; Link, D.C. Recruitment of the Inflammatory Subset of Monocytes to Sites of Ischemia Induces Angiogenesis in a Monocyte Chemoattractant Protein-1-Dependent Fashion. J. Leukoc. Biol. 2008, 84, 760–768. [Google Scholar] [CrossRef]
- Muhs, B.E.; Gagne, P.; Plitas, G.; Shaw, J.P.; Shamamian, P. Experimental Hindlimb Ischemia Leads to Neutrophil-Mediated Increases in Gastrocnemius MMP-2 and -9 Activity: A Potential Mechanism for Ischemia Induced MMP Activation. J. Surg. Res. 2004, 117, 249–254. [Google Scholar] [CrossRef]
- Stabile, E.; Burnett, M.S.; Watkins, C.; Kinnaird, T.; Bachis, A.; la Sala, A.; Miller, J.M.; Shou, M.; Epstein, S.E.; Fuchs, S. Impaired Arteriogenic Response to Acute Hindlimb Ischemia in CD4-Knockout Mice. Circulation 2003, 108, 205–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paoni, N.F.; Peale, F.; Wang, F.; Errett-Baroncini, C.; Steinmetz, H.; Toy, K.; Bai, W.; Williams, P.M.; Bunting, S.; Gerritsen, M.E.; et al. Time Course of Skeletal Muscle Repair and Gene Expression Following Acute Hind Limb Ischemia in Mice. Physiol. Genom. 2002, 11, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.W.; Stabile, E.; Kinnaird, T.; Shou, M.; Devaney, J.M.; Epstein, S.E.; Burnett, M.S. Temporal Patterns of Gene Expression after Acute Hindlimb Ischemia in Mice: Insights into the Genomic Program for Collateral Vessel Development. J. Am. Coll. Cardiol. 2004, 43, 474–482. [Google Scholar] [CrossRef] [Green Version]
- Duvall, C.L.; Weiss, D.; Robinson, S.T.; Alameddine, F.M.F.; Guldberg, R.E.; Taylor, W.R. The Role of Osteopontin in Recovery from Hind Limb Ischemia. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Jazwa, A.; Florczyk, U.; Grochot-Przeczek, A.; Krist, B.; Loboda, A.; Jozkowicz, A.; Dulak, J. Limb Ischemia and Vessel Regeneration: Is There a Role for VEGF? Vascul. Pharmacol. 2016, 86, 18–30. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Full Gene Name | Sequence 5′-3′ |
|---|---|---|
| Mouse primers | ||
| Ang 1 | Angiopoietin 1 | F:CAGTGGCTGCAAAAACTTGA R:TGGGCCATCTCCGACTTCAT |
| Ang2 | Angiopoietin 2 | F:CTCTTCTTTACGGATAGCAA R:AGCCACGGTCAACAACTCGC |
| Col3a1 | Collagen type III alpha 1 chain | F:ATCTATGAATGGTGGTTTTCA R:TTTTGCAGTGGTATGTAATGT |
| Cxcl1 | C-X-C motif chemokine ligand 1 | F:AAAGATGCTAAAAGGTGTCC R:GTATAGTGTTGTCAGAAGCC |
| Cxcl12 | C-X-C motif chemokine ligand 12 | F:CCTTCAGATTGTTGCACGGCT R:CCCACCACTGCCCTTGCATC |
| Cxcr4 | C-X-C chemokine receptor type 4 | F:AAACCTCTGAGGCGTTTGGT R:AGCAGGGTTCCTTGTTGGAG |
| Eef2 | Eukaryotic elongation factor 2 | F:AGAACATATTATTGCTGGCG R:AACAGGGTCAGATTTCTTG |
| Hmox1 | Heme oxygenase 1 | F:CCTCACTGGCAGGAAATCATC R:CCTCGTGGAGACGCTTTACATA |
| Kdr | Kinase insert domain receptor | F:CGGCCAAGTGATTGAGGCAG R:ATGAGGGCTCGATGCTCGCT |
| Mmp11 | Matrix metalloproteinase 11 | F:CAGATTTGGTTCTTCCAAGG R:AGATCTTGTTCTTCTCAGGAC |
| Myog | Myogenin | F:CAGTACATTGAGCGCCTACAG R:GGACCGAACTCCAGTGCAT |
| Myh3 | Myosin heavy chain 3 | F:TCTAGCCGGATGGTGGTCC R:GAATTGTCAGGAGCCACGAA |
| Spp1 | Secreted phosphoprotein 1 | F:CCATCTCAGAAGCAGAATCTCCTT R:GGTCATGGCTTTCATTGGAATT |
| Tgfb1 | Transforming growth factor beta 1 | F:GGATACCAACTATTGCTTGAG R:TGTCCAGGCTCCAAATATAG |
| Tie2 | Tek receptor tyrosine kinase | F:TGTCCAAAGGAGAATGGCTC R:GGCGGCATCCATCCGTAACC |
| Human primers | ||
| EEF2 | Eukaryotic elongation factor 2 | F:GAGATCCAGTGTCCAGAGCAG R:CTCGTTGACGGGCAGATAGG |
| VEGF | Vascular endothelial growth factor A | F:AAGGAGGAGGGCAGAATCAT R:CTCAGTGGGCACACACTCCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Podkalicka, P.; Mucha, O.; Kaziród, K.; Bronisz-Budzyńska, I.; Ostrowska-Paton, S.; Tomczyk, M.; Andrysiak, K.; Stępniewski, J.; Dulak, J.; Łoboda, A. Age-Dependent Dysregulation of Muscle Vasculature and Blood Flow Recovery after Hindlimb Ischemia in the mdx Model of Duchenne Muscular Dystrophy. Biomedicines 2021, 9, 481. https://doi.org/10.3390/biomedicines9050481
Podkalicka P, Mucha O, Kaziród K, Bronisz-Budzyńska I, Ostrowska-Paton S, Tomczyk M, Andrysiak K, Stępniewski J, Dulak J, Łoboda A. Age-Dependent Dysregulation of Muscle Vasculature and Blood Flow Recovery after Hindlimb Ischemia in the mdx Model of Duchenne Muscular Dystrophy. Biomedicines. 2021; 9(5):481. https://doi.org/10.3390/biomedicines9050481
Chicago/Turabian StylePodkalicka, Paulina, Olga Mucha, Katarzyna Kaziród, Iwona Bronisz-Budzyńska, Sophie Ostrowska-Paton, Mateusz Tomczyk, Kalina Andrysiak, Jacek Stępniewski, Józef Dulak, and Agnieszka Łoboda. 2021. "Age-Dependent Dysregulation of Muscle Vasculature and Blood Flow Recovery after Hindlimb Ischemia in the mdx Model of Duchenne Muscular Dystrophy" Biomedicines 9, no. 5: 481. https://doi.org/10.3390/biomedicines9050481