
Circulating Extracellular Vesicle Proteins and MicroRNA Profiles in Subcortical and Cortical-Subcortical Ischaemic Stroke


 , , ,
, , ,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
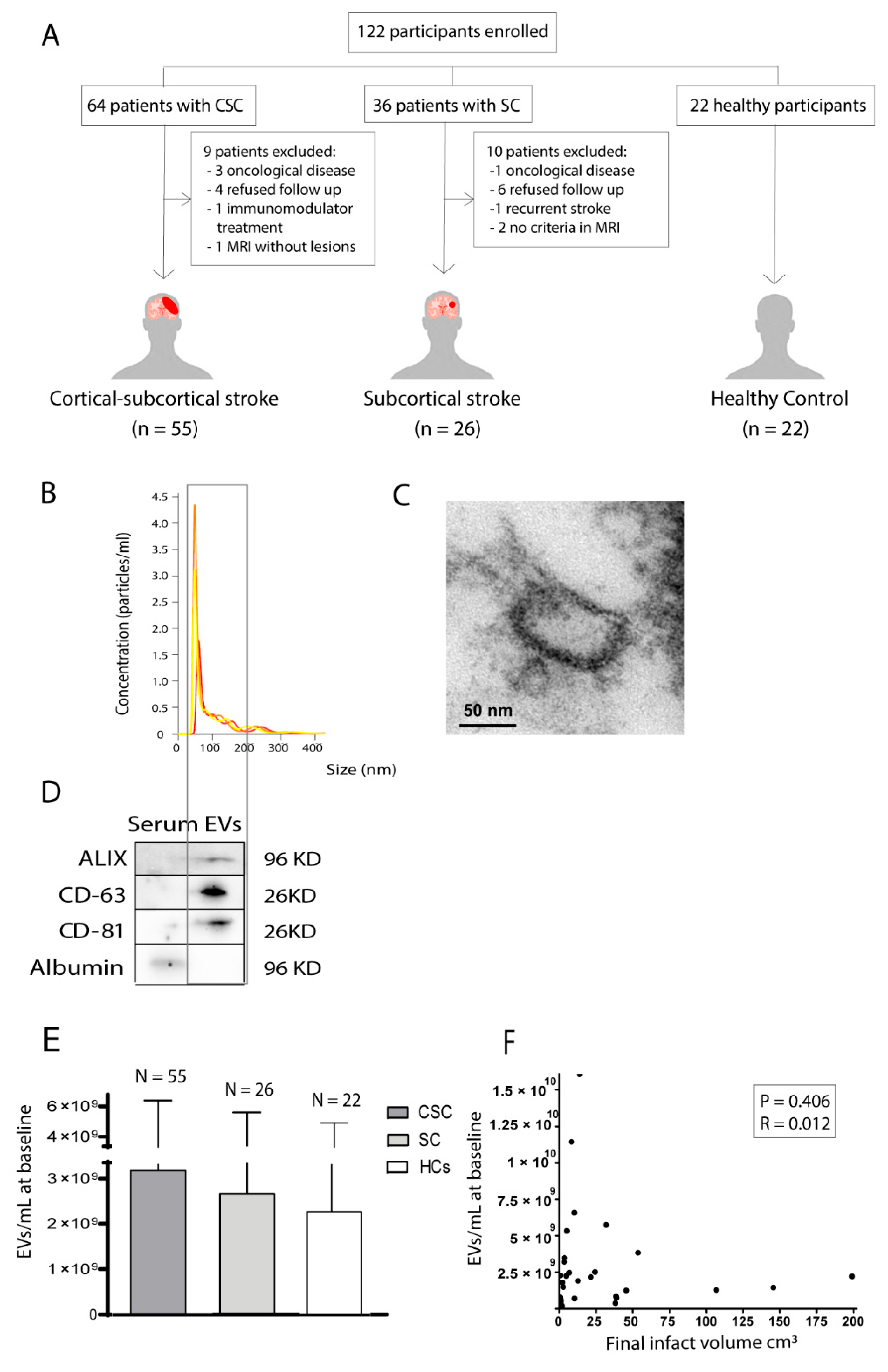
2.1. Study Design
2.2. Blood Extraction, Extracellular Vesicle Isolation and Characterisation
2.3. Quantification of Circulating Extracellular Vesicles
2.4. Proteomic Analysis of Circulating Extracellular Vesicles
2.5. Validation of Selected Proteins
2.6. Proteomic Analysis of Neuron-Derived Extracellular Vesicles
2.7. RNA Extraction and Reverse Transcription Quantitative Polymerase Chain Reaction
2.8. miRNA Array Profiling
2.9. Validation of Selected miRNAs
2.10. Association between miRNAs and Outcomes
2.11. Bioinformatic Analysis
2.12. Statistical Analysis
3. Results
3.1. Association between the Quantity of Circulating Extracellular Vesicles of Healthy Controls and Patients with Either Subcortical or Cortical-Subcortical Stroke
3.2. Correlation between the Quantity of Circulating Extracellular Vesicles and Final Lesion Volume
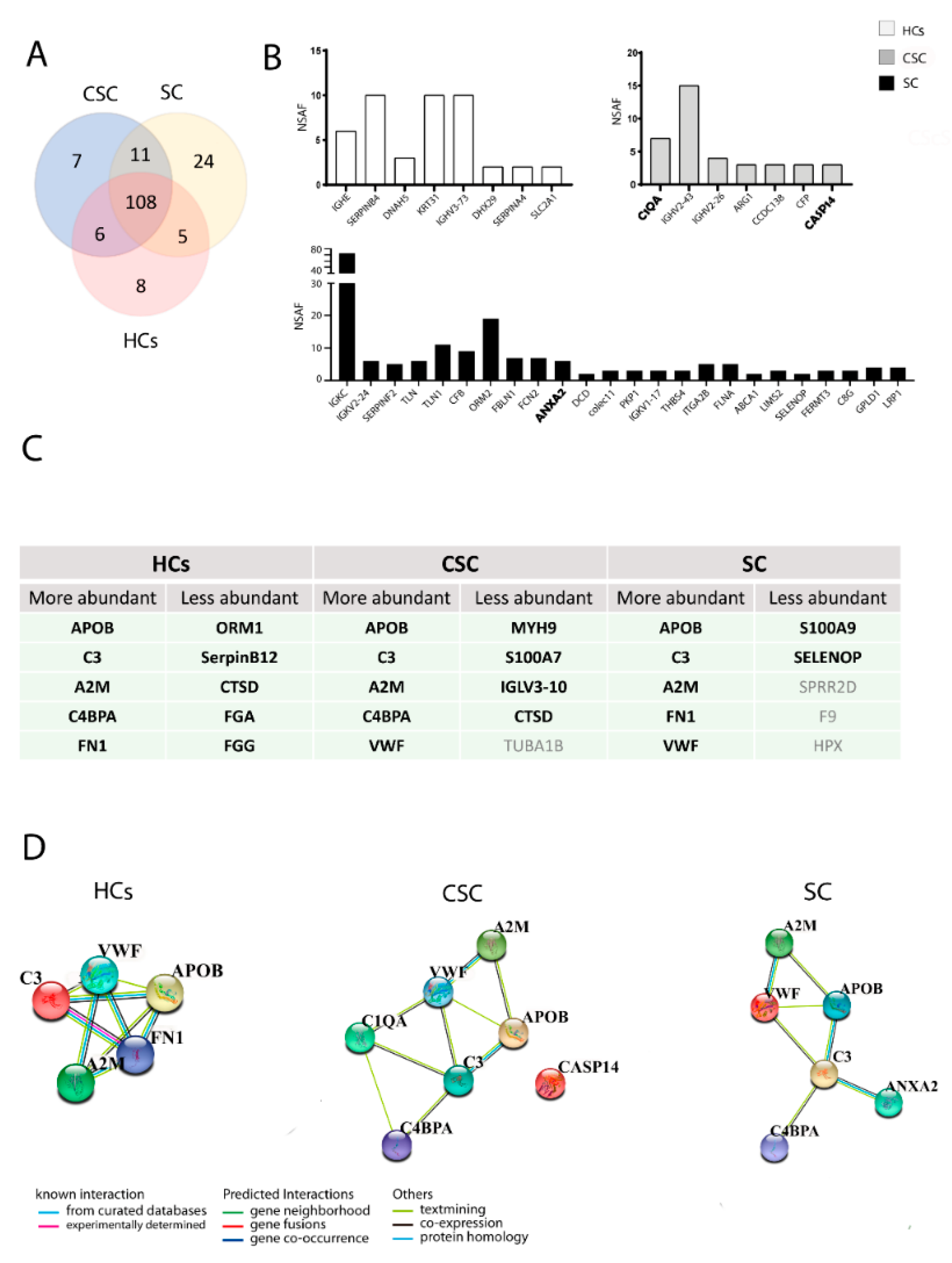
3.3. Proteomic Analysis
3.4. Validation of the Identified Proteins
3.5. Proteomic Analysis of Neuron-Derived Extracellular Vesicles
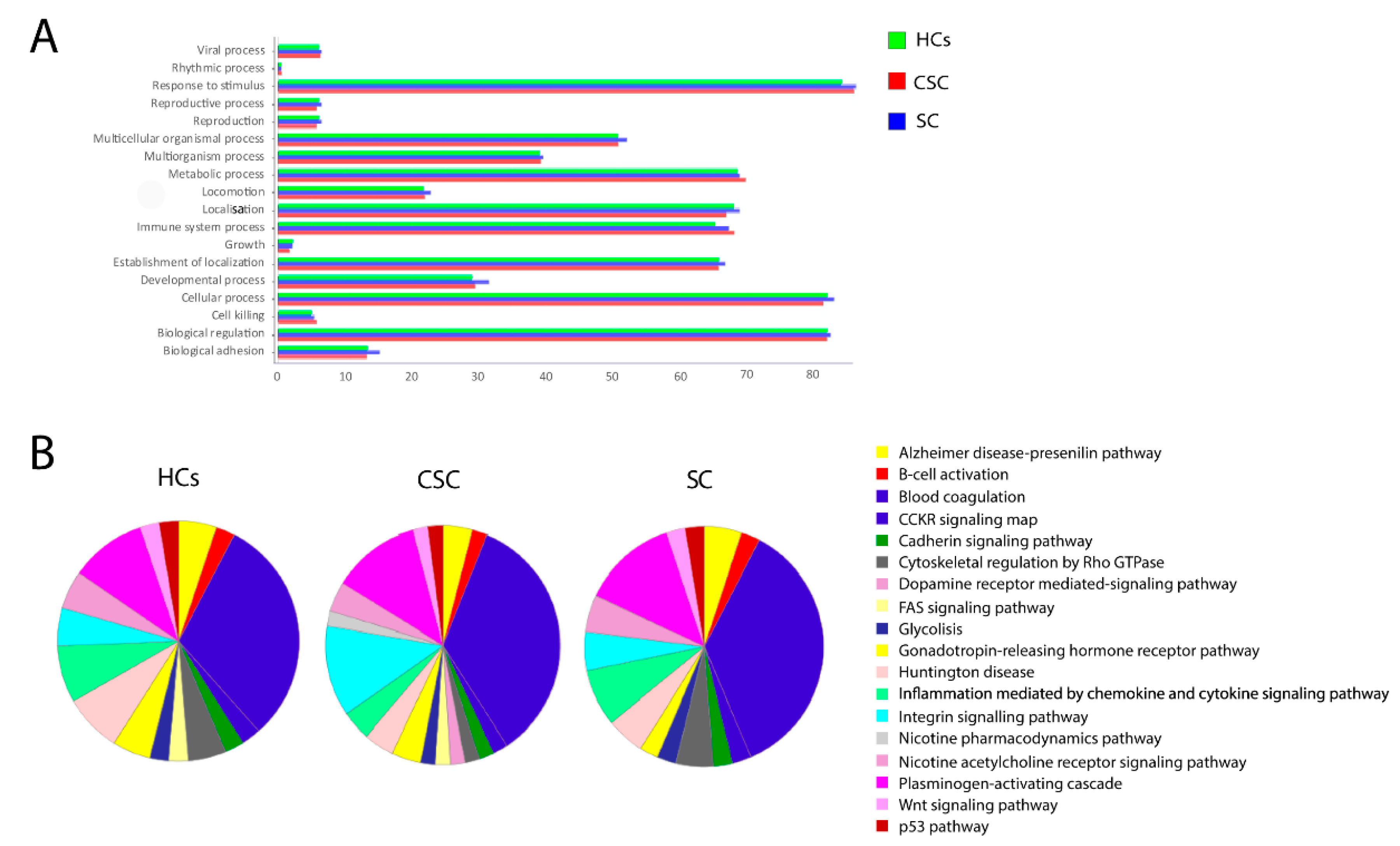
3.6. Gene Ontology Enrichment
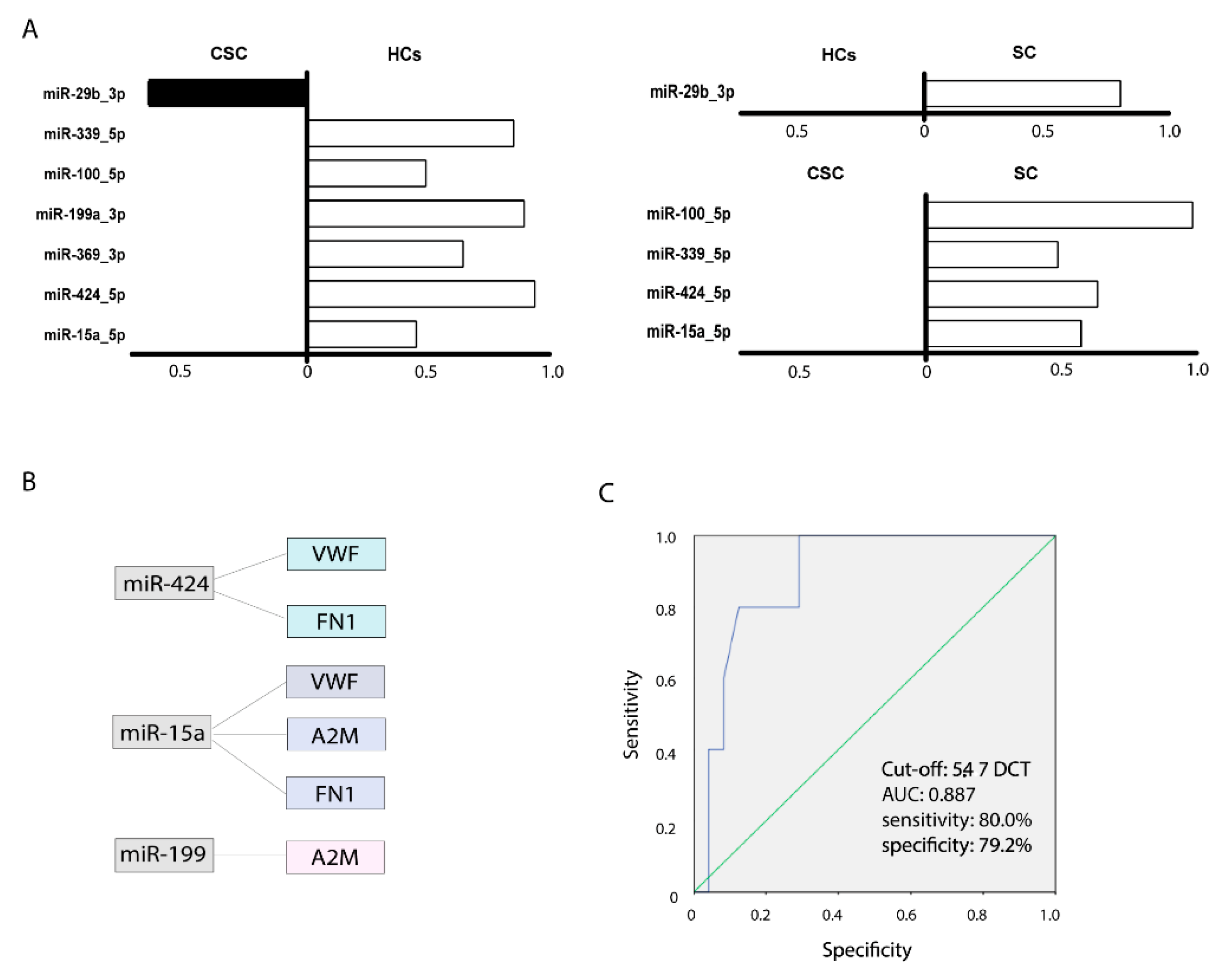
3.7. miRNA Analysis
3.8. Bioinformatics Analysis of Predicted Targets
3.9. miRNA as a Prognostic Biomarker in Ischaemic Stroke
4. Discussion
4.1. Cargo of Circulating Extracellular Vesicles in Cortical-Subcortical Ischaemic Stroke
4.2. Cargo of Circulating Extracellular Vesicles in Subcortical Ischaemic Stroke
4.3. Cargo of Circulating Neural-Derived Extracellular Vesicles
4.4. Extracellular Vesicle Cargo Related to Recovery
4.5. Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ortega, L.O.; Laso-García, F.; Frutos, M.G.-D.; Fuentes, B.; Diekhorst, L.; Díez-Tejedor, E.; Gutiérrez-Fernández, M. Role of Exosomes as a Treatment and Potential Biomarker for Stroke. Transl. Stroke Res. 2019, 10, 241–249. [Google Scholar] [CrossRef]
- Van Kralingen, J.C.; McFall, A.; Ord, E.N.J.; Coyle, T.F.; Bissett, M.; McClure, J.D.; McCabe, C.; Macrae, I.M.; Dawson, J.; Work, L.M. Altered Extracellular Vesicle MicroRNA Expression in Ischemic Stroke and Small Vessel Disease. Transl. Stroke Res. 2019, 10, 495–508. [Google Scholar] [CrossRef] [Green Version]
- Kalani, M.Y.S.; Alsop, E.; Meechoovet, B.; Beecroft, T.; Agrawal, K.; Whitsett, K.G.; Huentelman, M.J.; Spetzler, R.F.; Nakaji, P.; Kim, S.; et al. Extracellular microRNAs in blood differentiate between ischaemic and haemorrhagic stroke subtypes. J. Extracell. Vesicles 2020, 9, 1713540. [Google Scholar] [CrossRef] [Green Version]
- Otero-Ortega, L.; Alonso-López, E.; Pérez-Mato, M.; Laso-García, F.; Gómez-de Frutos, M.C.; Diekhorst, L.; García-Bermejo, M.L.; Conde-Moreno, E.; Fuentes, B.; Alonso de Leciñana, M.; et al. Similarities and differences in Extracellular Vesicle Profiles between Ischaemic Stroke and myocardial infarction. Biomedicines 2020, 24, 8. [Google Scholar] [CrossRef]
- Zheng, X.; Hermann, D.M.; Bähr, M.; Doeppner, T.R. The role of small extracellular vesicles in cerebral and myocardial ischemia-Molecular signals, treatment targets, and future clinical translation. STEM CELLS 2021, 39, 403–413. [Google Scholar] [CrossRef]
- Venkat, P.; Chen, J.; Chopp, M. Exosome-mediated amplification of endogenous brain repair mechanisms and brain and systemic organ interaction in modulating neurological outcome after stroke. Br. J. Pharmacol. 2018, 38, 2165–2178. [Google Scholar] [CrossRef]
- Jafarzadeh-Esfehani, R.; Soudyab, M.; Parizadeh, S.M.; Jaripoor, M.E.; Nejad, P.S.; Shariati, M.; Nabavi, A.S. Circulating Exosomes and Their Role in Stroke. Curr. Drug Targets 2019, 21, 89–95. [Google Scholar] [CrossRef]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef]
- Wang, M.-M.; Feng, Y.-S.; Tan, Z.-X.; Xing, Y.; Dong, F.; Zhang, F. The role of exosomes in stroke. Mol. Biol. Rep. 2020, 47, 6217–6228. [Google Scholar] [CrossRef]
- Huo, S.; Kränkel, N.; Nave, A.H.; Sperber, P.S.; Rohmann, J.L.; Piper, S.K.; Heuschmann, P.; Landmesser, U.; Endres, M.; Siegerink, B.; et al. Endothelial and leukocyte-derived microvesicles and cardiovascular risk after stroke—PROSCIS-B. Neurology 2020, 96, e937–e946. [Google Scholar] [CrossRef]
- Brouns, R.; Bossche, J.V.D.; De Surgeloose, D.; Sheorajpanday, R.; De Deyn, P.P. Clinical and biochemical diagnosis of small-vessel disease in acute ischemic stroke. J. Neurol. Sci. 2009, 285, 185–190. [Google Scholar] [CrossRef]
- Stenz, K.T.; Just, J.; Blauenfeldt, R.A.; Drasbek, K.R. Extracellular Vesicles in Acute Stroke Diagnostics. Biomedicines 2020, 8, 248. [Google Scholar] [CrossRef]
- Smith, E.E.; Saposnik, G.; Biessels, G.J.; Doubal, F.N.; Fornage, M.; Gorelick, P.B.; Greenberg, S.M.; Higashida, R.T.; Kasner, S.E.; Seshadri, S. Prevention of Stroke in Patients with Silent Cerebrovascular Disease: A Scientific Statement for Healthcare Professionals From the American Heart Association/American Stroke Association. Stroke 2017, 48, e44–e71. [Google Scholar] [CrossRef]
- Coughlan, C.; Bruce, K.D.; Burgy, O.; Boyd, T.D.; Michel, C.R.; Garcia-Perez, J.E.; Adame, V.; Anton, P.; Bettcher, B.M.; Chial, H.J.; et al. Exosome Isolation by Ultracentrifugation and Precipitation and Techniques for Downstream Analyses. Curr. Protoc. Cell Biol. 2020, 88, e110. [Google Scholar] [CrossRef]
- Camino, T.; Lago-Baameiro, N.; Bravo, S.; Sueiro, A.; Couto, I.; Santos, F.; Baltar, J.; Casanueva, F.; Pardo, M. Vesicles Shed by Pathological Murine Adipocytes Spread Pathology: Characterization and Functional Role of Insulin Resistant/Hypertrophied Adiposomes. Int. J. Mol. Sci. 2020, 21, 2252. [Google Scholar] [CrossRef] [Green Version]
- Hermida, L. Proteomic analysis of extracellular vesicles derived from platelet concentrates treated with Mirasol® identifies biomarkers of platelet storage lesion. J. Proteom. 2020, 210, 103529. [Google Scholar] [CrossRef]
- Brenna, S.; Krisp, C.; Altmeppen, H.C.; Magnus, T.; Puig, B. Brain-Derived Extracellular Vesicles in Health and Disease: A Methodological Perspective. Int. J. Mol. Sci. 2021, 22, 1365. [Google Scholar] [CrossRef]
- Gallo, A.; Tandon, M.; Alevizos, I.; Illei, G.G. The majority of microRNAs detectable in serum and saliva is concentrated in exosomes. PLoS ONE 2012, 7, e30679. [Google Scholar] [CrossRef] [Green Version]
- Herrero-Aguayo, V.; Jiménez-Vacas, J.M.; Sáez-Martínez, P.; Gómez-Gómez, E.; López-Cánovas, J.L.; Garrido-Sánchez, L.; Herrera-Martínez, A.D.; García-Bermejo, L.; Macías-González, M.; López-Miranda, J.; et al. Influence of Obesity in the miRNome: miR-4454, a Key Regulator of Insulin Response Via Splicing Modulation in Prostate. J. Clin. Endocrinol. Metab. 2021, 106, e469–e484. [Google Scholar] [CrossRef]
- Wouters, A.; Nysten, C.; Thijs, V.; Lemmens, R. Prediction of Outcome in Patients with Acute Ischemic Stroke Based on Initial Severity and Improvement in the First 24 h. Front. Neurol. 2018, 9, 308. [Google Scholar] [CrossRef] [Green Version]
- Tsivgoulis, G.; Katsanos, A.H.; Kadlecová, P.; Czlonkowska, A.; Kobayashi, A.; Brozman, M.; Švigelj, V.; Csiba, L.; Fekete, K.; Kõrv, J.; et al. Intravenous thrombolysis for ischemic stroke in the golden hour: Propensity-matched analysis from the SITS-EAST registry. J. Neurol. 2017, 264, 912–920. [Google Scholar] [CrossRef]
- Searle, B.C. Scaffold: A bioinformatic tool for validating MS/MS-based proteomic studies. Proteomics 2010, 10, 1265–1269. [Google Scholar] [CrossRef]
- String. Available online: https://string-db.org (accessed on 10 March 2021).
- MirTarget. Available online: http://www.mirdb.org (accessed on 10 March 2021).
- Ji, Q.; Ji, Y.; Peng, J.; Zhou, X.; Chen, X.; Zhao, H.; Xu, T.; Chen, L.; Xu, Y. Increased Brain-Specific MiR-9 and MiR-124 in the Serum Exosomes of Acute Ischemic Stroke Patients. PLoS ONE 2016, 11, e0163645. [Google Scholar] [CrossRef] [Green Version]
- Li, D.-B.; Liu, J.-L.; Wang, W.; Li, R.-Y.; Yu, D.-J.; Lan, X.-Y.; Li, J.-P. Plasma exosomal miR-422a and miR-125b-2-3p serve as biomarkers for ischemic stroke. Curr. Neurovasc. Res. 2018, 14, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Nayak, A.; Pednekar, L.; Reid, K.B.; Kishore, U. Complement and non-complement activating functions of C1q: A prototypical innate immune molecule. Innate Immun. 2011, 18, 350–363. [Google Scholar] [CrossRef]
- Pisalyaput, K.; Tenner, A.J. Complement component C1q inhibits betaamyloid- and serum amyloid P-induced neurotoxicity via caspase- and calpain-independent mechanisms. J. Neurochem. 2008, 104, 696–707. [Google Scholar]
- Benoit, M.E.; Tenner, A.J. Complement Protein C1q-Mediated Neuroprotection Is Correlated with Regulation of Neuronal Gene and MicroRNA Expression. J. Neurosci. 2011, 31, 3459–3469. [Google Scholar] [CrossRef] [Green Version]
- Shimomura, R.; Nezu, T.; Hosomi, N.; Aoki, S.; Sugimoto, T.; Kinoshita, N.; Araki, M.; Takahashi, T.; Maruyama, H.; Matsumoto, M. Alpha-2-macroglobulin as a Promising Biological Marker of Endothelial Function. J. Atheroscler. Thromb. 2018, 25, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Singh, Y.; Garden, O.A.; Lang, F.; Cobb, B.S. MicroRNA-15b/16 Enhances the Induction of Regulatory T Cells by Regulating the Expression of Rictor and mTOR. J. Immunol. 2015, 195, 5667–5677. [Google Scholar] [CrossRef] [Green Version]
- Haskard, D.; Boyle, J.J.; Mason, J. The role of complement in atherosclerosis. Curr. Opin. Lipidol. 2008, 19, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Cheung, E.Y.L.; Bos, M.J.; Leebeek, F.W.G.; Koudstaal, P.J.; Hofman, A.; De Maat, M.P.M.; Breteler, M.M.B. Variation in fibrinogen FGG and FGA genes and risk of stroke. The Rotterdam Study. Thromb. Haemost. 2008, 100, 308–313. [Google Scholar] [CrossRef]
- Zhai, Z.; Wu, J.; Xu, X.; Ding, K.; Ni, R.; Hu, W.; Sun, Z.; Ni, H. Fibrinogen controls human platelet fibronectin internalization and cell-surface retention. J. Thromb. Haemost. 2007, 5, 1740–1746. [Google Scholar] [CrossRef]
- Charla, E.; Mercer, J.; Maffia, P.; Nicklin, S.; Charla, E.; Mercer, J.; Maffia, P.; Nicklin, S. Extracellular vesicle signalling in atherosclerosis. Cell. Signal. 2020, 75, 109751. [Google Scholar] [CrossRef]
- Kölm, R.; Schaller, M.; Roumenina, L.T.; Niemiec, I.; Hovinga, J.A.K.; Khanicheh, E.; Kaufmann, B.A.; Hopfer, H.; Trendelenburg, M. Von Willebrand Factor Interacts with Surface-Bound C1q and Induces Platelet Rolling. J. Immunol. 2016, 197, 3669–3679. [Google Scholar] [CrossRef]
- Datta, A.; Akatsu, H.; Heese, K.; Sze, S.K. Quantitative clinical proteomic study of autopsied human infarcted brain specimens to elucidate the deregulated pathways in ischemic stroke pathology. J. Proteom. 2013, 91, 556–568. [Google Scholar] [CrossRef]
- Datta, A.; Park, J.E.; Li, X.; Zhang, H.; Ho, Z.S.; Heese, K.; Lim, S.K.; Tam, J.P.; Sze, S.K. Phenotyping of an in vitro model of ischemic penumbra by iTRAQ-based shotgun quantitative proteomics. J. Proteome Res. 2010, 9, 472–484. [Google Scholar] [CrossRef]
- Fan, X.; Jiang, Y.; Yu, Z.; Liu, Q.; Guo, S.; Sun, X.; Van Leyen, K.; Ning, M.; Gao, X.; Lo, E.H.; et al. Annexin A2 Plus Low-Dose Tissue Plasminogen Activator Combination Attenuates Cerebrovascular Dysfunction After Focal Embolic Stroke of Rats. Transl. Stroke Res. 2017, 8, 549–559. [Google Scholar] [CrossRef]
- Sun, S.-L.; Shu, Y.-G.; Tao, M.-Y. LncRNA CCAT2 promotes angiogenesis in glioma through activation of VEGFA signalling by sponging miR-424. Mol. Cell. Biochem. 2020, 468, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Cao, Y.; Yang, X.; Cai, P.; Kang, L.; Zhu, X.; Luo, H.; Lu, L.; Wei, L.; Bai, X.; et al. ADAMTS13 controls vascular remodeling by modifying VWF reactivity during stroke recovery. Blood 2017, 130, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pankratz, F.; Hohnloser, C.; Bemtgen, X.; Jaenich, C.; Kreuzaler, S.; Hoefer, I.; Pasterkamp, G.; Mastroianni, J.; Zeiser, R.; Smolka, C.; et al. MicroRNA-100 Suppresses Chronic Vascular Inflammation by Stimulation of Endothelial Autophagy. Circ. Res. 2018, 122, 417–432. [Google Scholar] [CrossRef] [PubMed]
- Khoshnam, S.E.; Winlow, W.; Farbood, Y.; Moghaddam, H.F.; Farzaneh, M. Emerging Roles of microRNAs in Ischemic Stroke: As Possible Therapeutic Agents. J. Stroke 2017, 19, 166–187. [Google Scholar] [CrossRef] [PubMed]
- Thulin, Å.; Lindbäck, J.; Granger, C.B.; Wallentin, L.; Lind, L.; Siegbahn, A. Extracellular vesicles in atrial fibrillation and stroke. Thromb. Res. 2020, 193, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Tabb, D.; Vega-Montoto, L.; Rudnick, P.A.; Variyath, A.M.; Ham, A.-J.L.; Bunk, D.M.; Kilpatrick, L.E.; Billheimer, D.D.; Blackman, R.K.; Cardasis, H.L.; et al. Repeatability and Reproducibility in Proteomic Identifications by Liquid Chromatography—Tandem Mass Spectrometry. J. Proteome Res. 2010, 9, 761–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CSC n = 55 | SC n = 26 | HC n = 22 | p | |
|---|---|---|---|---|
| Demographic Data | ||||
| Age, years [mean (SD)] | 70.65 (15.39) * | 61.65 (11.83) | 61 (12.74) † | 0.003 |
| Male, n (%) | 24 (43.6) | 18 (69.2) | 7 (31.8) | 0.002 |
| Medical History | ||||
| Hypertension, n (%) | 35 (63.6) | 19 (73.1) | 3 (13.63) | 0.001 |
| Diabetes mellitus, n (%) | 10 (18.2) | 5 (19.2) | 2 (9.09) | 0.53 |
| Dyslipidaemia, n (%) | 25 (45.5) | 11 (42.3) | 3 (13.63) † | 0.001 |
| Smoker, n (%) | 9 (16.4) | 9 (34.6) | 3 (13.63) | 0.001 |
| Coronary arterial disease, n (%) | 6 (10.9%) | 0 (0%) | 0 (0%) | 0.064 |
| Atrial fibrillation, n (%) | 15 (27.27%) | 2 (7.69%) | 0 (0%) † | 0.004 |
| Infection, n (%) | 6 (10.9%) | 1 (3.84%) | 0 (0%) | 0.178 |
| Charlson comorbidity index, (median; IQR) | 1.00 (2.00) | 1.00 (2.25) | 0.00 (1.00) † | 0.001 |
| Treatments | ||||
| Patients undergoing reperfusion therapy, n (%) | 34 (61.81%) | 5 (19.23%) | -- | <0.001 |
| IVT alone, n (%) | 18 (34.54) | 5 (19.2) | -- | 0.001 |
| MT alone, n (%) | 3 (11.53) | 0 (0) | -- | 0.002 |
| IVT and MT, n (%) | 13 (32.73) | 0 (0) | -- | 0.008 |
| Recovery | ||||
| Baseline NIHSS (median; IQR) | 9.00 (9.75) | 3.00 (3.25) | -- | <0.001 |
| 3 months NIHSS (median; IQR) | 0.00 (1.00) | 0.00 (1.00) | -- | 0.159 |
| 3 months mRS (median; IQR) | 1.00 (2.00) | 1.00 (2.00) | -- | 0.265 |
| Percentage improvement from baseline NIHSS, mean (SD) | 87.59% (22.85%) | 78.65% (31.45%) | -- | 0.249 |
| Patients who improved more than 50% baseline NIHSS, n (%) | 37 (67.27%) | 17 (56.38%) | -- | 0.090 |
| Deaths, n (%) | 1 (1.81%) | 0 (0%) | -- | 0.323 |
| Final Infarct Volume | ||||
| FLAIR, cm3 [mean (SD)] | 30.95 (13.17) | 3.15 (3.4) | -- | 0.012 |
| DWI, cm3 [mean (SD)] | 36.17 (50.23) | 2.7 (2.6) | -- | 0.008 |
| Baseline Vascular Disease Burden | ||||
| Fazekas PV score (median; IQR) | 1(2) | 1(2) | - | 0.8 |
| Fazekas DWM score (median; IQR) | 1(2) | 1(2) | - | 0.759 |
| Silent brain infarcts, n (%) | 4 (7,1) | 9 (36) | - | 0.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otero-Ortega, L.; Alonso-López, E.; Pérez-Mato, M.; Laso-García, F.; Gómez-de Frutos, M.C.; Diekhorst, L.; García-Bermejo, M.L.; Conde-Moreno, E.; Fuentes, B.; de Leciñana, M.A.; et al. Circulating Extracellular Vesicle Proteins and MicroRNA Profiles in Subcortical and Cortical-Subcortical Ischaemic Stroke. Biomedicines 2021, 9, 786. https://doi.org/10.3390/biomedicines9070786
Otero-Ortega L, Alonso-López E, Pérez-Mato M, Laso-García F, Gómez-de Frutos MC, Diekhorst L, García-Bermejo ML, Conde-Moreno E, Fuentes B, de Leciñana MA, et al. Circulating Extracellular Vesicle Proteins and MicroRNA Profiles in Subcortical and Cortical-Subcortical Ischaemic Stroke. Biomedicines. 2021; 9(7):786. https://doi.org/10.3390/biomedicines9070786
Chicago/Turabian StyleOtero-Ortega, Laura, Elisa Alonso-López, María Pérez-Mato, Fernando Laso-García, Mari Carmen Gómez-de Frutos, Luke Diekhorst, María Laura García-Bermejo, Elisa Conde-Moreno, Blanca Fuentes, María Alonso de Leciñana, and et al. 2021. "Circulating Extracellular Vesicle Proteins and MicroRNA Profiles in Subcortical and Cortical-Subcortical Ischaemic Stroke" Biomedicines 9, no. 7: 786. https://doi.org/10.3390/biomedicines9070786
APA StyleOtero-Ortega, L., Alonso-López, E., Pérez-Mato, M., Laso-García, F., Gómez-de Frutos, M. C., Diekhorst, L., García-Bermejo, M. L., Conde-Moreno, E., Fuentes, B., de Leciñana, M. A., Bravo, S. B., Díez-Tejedor, E., & Gutiérrez-Fernández, M. (2021). Circulating Extracellular Vesicle Proteins and MicroRNA Profiles in Subcortical and Cortical-Subcortical Ischaemic Stroke. Biomedicines, 9(7), 786. https://doi.org/10.3390/biomedicines9070786